
RNAi by Soaking Aedes aegypti Pupae in dsRNA


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
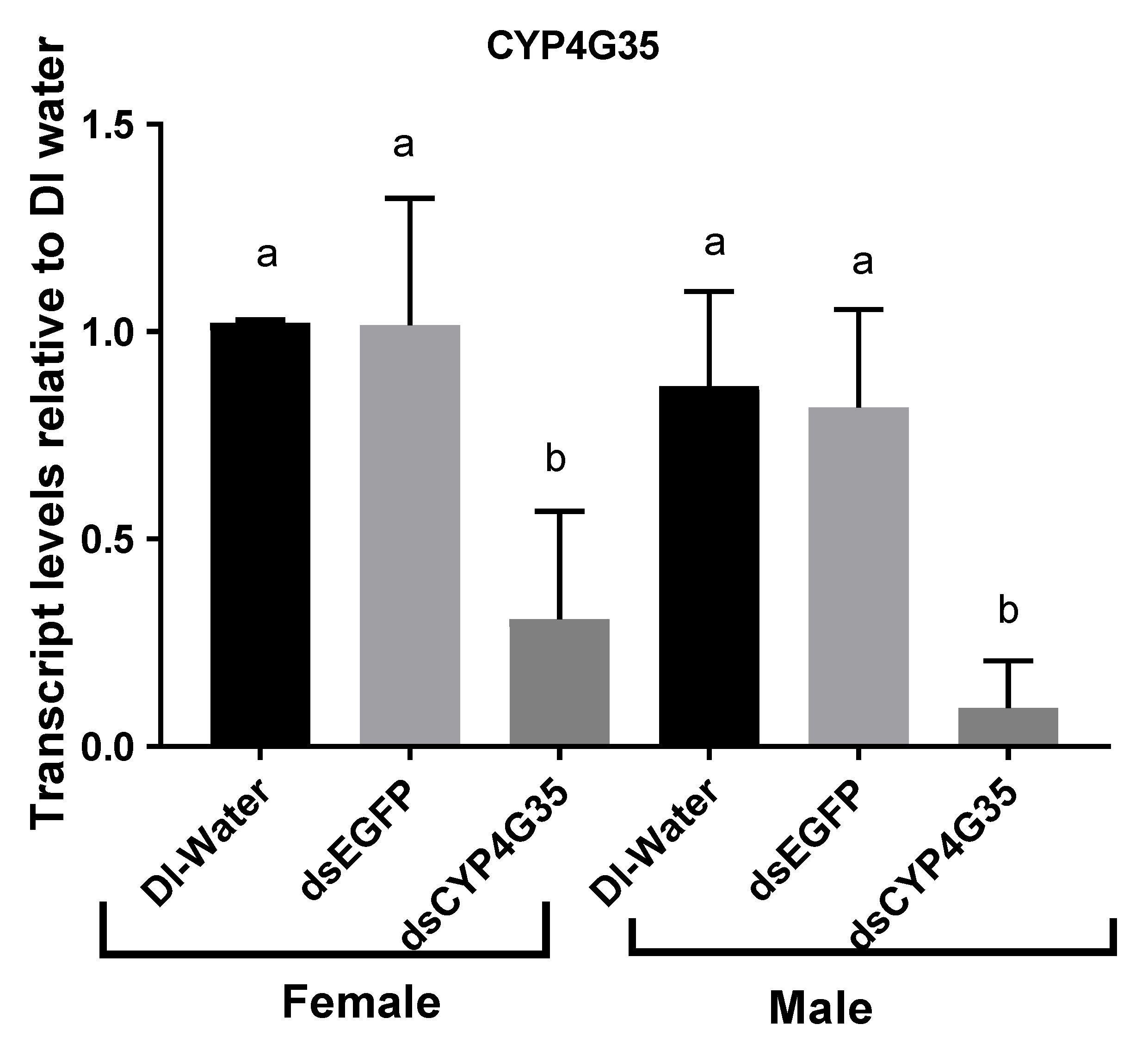
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Mello, C.C.; Conte, D., Jr. Revealing the world of RNA interference. Nature 2004, 431, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Kemphues, K.J. par-1, a gene required for establishing polarity in C. elegans embryos, encodes a putative Ser/Thr kinase that is asymmetrically distributed. Cell 1995, 81, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Franz, A.W.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA interference-based resistance to dengue virus type 2 in genetically modified Aedes aegypti. Proc. Natl. Acad. Sci. USA 2006, 103, 4198–4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhaes, T.; Brackney, D.E.; Beier, J.C.; Foy, B.D. Silencing an Anopheles gambiae catalase and sulfhydryl oxidase increases mosquito mortality after a blood meal. Arch. Insect Biochem. Physiol. 2008, 68, 134–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, C.; Denlinger, D.L. Insulin signaling and FOXO regulate the overwintering diapause of the mosquito Culex pipiens. Proc. Natl. Acad. Sci. USA 2008, 105, 6777–6781. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Denlinger, D.L. RNA interference directed against ribosomal protein S3a suggests a link between this gene and arrested ovarian development during adult diapause in Culex pipiens. Insect Mol. Biol. 2010, 19, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Gulia-Nuss, M.; Robertson, A.E.; Brown, M.R.; Strand, M.R. Insulin-like peptides and the target of rapamycin pathway coordinately regulate blood digestion and egg maturation in the mosquito Aedes aegypti. PLoS ONE 2011, 6, e20401. [Google Scholar] [CrossRef]
- Gulia-Nuss, M.; Eum, J.H.; Strand, M.R.; Brown, M.R. Ovary ecdysteroidogenic hormone activates egg maturation in the mosquito Georgecraigius atropalpus after adult eclosion or a blood meal. J. Exp. Biol. 2012, 215, 3758–3767. [Google Scholar] [CrossRef] [Green Version]
- Nuss, A.B.; Brown, M.R.; Murty, U.S.; Gulia-Nuss, M. Insulin receptor knockdown blocks filarial parasite development and alters egg production in the southern house mosquito, Culex quinquefasciatus. PLoS Negl. Trop. Dis. 2018, 12, e0006413. [Google Scholar] [CrossRef]
- Blitzer, E.J.; Vyazunova, I.; Lan, Q. Functional analysis of AeSCP-2 using gene expression knockdown in the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2005, 14, 301–307. [Google Scholar] [CrossRef]
- Liu, C.; Pitts, R.J.; Bohbot, J.D.; Jones, P.L.; Wang, G.; Zwiebel, L.J. Distinct olfactory signaling mechanisms in the malaria vector mosquito Anopheles gambiae. PLoS Biol. 2010, 31, e1000467. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zwiebel, L.J. Molecular characterization of larval peripheral thermosensory responses of the malaria vector mosquito Anopheles gambiae. PLoS ONE 2013, 7, e72595. [Google Scholar] [CrossRef] [Green Version]
- Regna, K.; Harrison, R.M.; Heyse, S.A.; Chiles, T.C.; Michel, K.; Muskavitch, M.A. RNAi Trigger Delivery into Anopheles gambiae Pupae. J. Vis. Exp. 2016, 109, 53738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munawar, K.; Alahmed, A.M.; Khalil, S.M. Delivery Methods for RNAi in Mosquito Larvae. J. Insect Sci. 2020, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.D.; Wong, S.; Ryan, C.P.; Whyard, S. Oral delivery of double-stranded RNA in larvae of the yellow fever mosquito, Aedes aegypti: Implications for pest mosquito control. J. Insect Sci. 2013, 13, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Martinez, G.; Meuti, M.; Denlinger, D.L. Rehydration driven RNAi: A novel approach for effectively delivering dsRNA to mosquito larvae. J. Med. Entomol. 2012, 49, 215–218. [Google Scholar] [CrossRef]
- Kumar, A.; Wang, S.; Ou, R.; Samrakandi, M.; Beerntsen, B.; Sayre, R. Development of an RNAi based microalgal larvicide to control mosquitoes. Malar. World J. 2013, 4, 1–7. [Google Scholar]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambie). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef]
- Cancino-Rodezno, A.; Alexander, C.; Villaseñor, R.; Pacheco, S.; Porta, H.; Pauchet, Y.; Soberón, M.; Gill, S.S.; Bravo, A. The mitogen-activated protein kinase p38 is involved in insect defense against Cry toxins from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2010, 40, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Pooraiiouby, R.; Sharma, A.; Beard, J.; Reyes, J.; Nuss, A.; Gulia-Nuss, M. Nutritional Quality during Development Alters Insulin-Like Peptides’ Expression and Physiology of the Adult Yellow Fever Mosquito, Aedes aegypti. Insects 2018, 9, 110. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.; Ayala-Chavez, C.; Sharma, A.; Pham, M.; Nuss, A.B.; Gulia-Nuss, M. Blood Digestion by Trypsin-Like Serine Proteases in the Replete Lyme Disease Vector Tick, Ixodes scapularis. Insects 2020, 11, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feyereisen, R. Origin and evolution of the CYP4G subfamily in insects, cytochrome P450 enzymes involved in cuticular hydrocarbon synthesis. Mol. Phylogenet. Evol. 2020, 143, 106695. [Google Scholar] [CrossRef]
- Kefi, M.; Balabanidou, V.; Douris, V.; Lycett, G.; Feyereisen, R.; Vontas, J. Two functionally distinct CYP4G genes of Anopheles gambiae contribute to cuticular hydrocarbon biosynthesis. Insect Biochem. Mol. Biol. 2019, 110, 52–59. [Google Scholar] [CrossRef]
- Garver, L.; Dimopoulos, G. Protocol for RNAi assays in adult mosquitoes (A. gambiae). J. Vis. Exp. 2007, 5, 230. [Google Scholar] [CrossRef]
- Airs, P.M.; Bartholomay, L.C. RNA Interference for Mosquito and Mosquito-Borne Disease Control. Insects 2017, 8, 4. [Google Scholar] [CrossRef]
- Bona, A.C.; Chitolina, R.F.; Fermino, M.L.; de Castro Poncio, L.; Weiss, A.; Lima, J.B.; Paldi, N.; Bernardes, E.S.; Henen, J.; Maori, E. Larval application of sodium channel homologous dsRNA restores pyrethroid insecticide susceptibility in a resistant adult mosquito population. Parasites Vectors 2016, 9, 397. [Google Scholar] [CrossRef] [Green Version]
- Peng, R.; Maklokova, V.I.; Chandrashekhar, J.H.; Lan, Q. In vivo functional genomic studies of sterol carrier protein-2 gene in the yellow fever mosquito. PLoS ONE 2011, 6, e18030. [Google Scholar] [CrossRef] [Green Version]
- Lopez, S.; Guimarães-Ribeiro, V.; Rodriguez, J.; Dorand, F.; Salles, S.T.; Sá-Guimarães, E.T.; Alvarenga, E.; Melo, A.; Almeida, V.R.; Moreira, F.M. RNAi-based bioinsecticide for Aedes mosquito control. Sci. Rep. 2019, 9, 4038. [Google Scholar] [CrossRef] [PubMed]
- Riehle, M.A.; Fan, Y.; Cao, C.; Brown, M.R. Molecular characterization of insulin-like peptides in the yellow fever mosquito, Aedes aegypti: Expression, cellular localization, and phylogeny. Peptides 2006, 27, 2547–2560. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| DI Water | dsEGFP | dsCYP4G35 | |
|---|---|---|---|
| Total number of pupae treated | 65 | 65 | 65 |
| Number of pupae survived to adulthood | 34 | 28 | 28 |
| Pupal mortality (%) | 41.7% | 56.92% | 56.92% |
| Total number of males eclosed | 12 | 19 | 18 |
| Total number of females eclosed | 22 | 9 | 10 |
| Total adults survived to day 2 (%) | 100% | 100% | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arshad, F.; Sharma, A.; Lu, C.; Gulia-Nuss, M. RNAi by Soaking Aedes aegypti Pupae in dsRNA. Insects 2021, 12, 634. https://doi.org/10.3390/insects12070634
Arshad F, Sharma A, Lu C, Gulia-Nuss M. RNAi by Soaking Aedes aegypti Pupae in dsRNA. Insects. 2021; 12(7):634. https://doi.org/10.3390/insects12070634
Chicago/Turabian StyleArshad, Fiza, Arvind Sharma, Charleen Lu, and Monika Gulia-Nuss. 2021. "RNAi by Soaking Aedes aegypti Pupae in dsRNA" Insects 12, no. 7: 634. https://doi.org/10.3390/insects12070634
APA StyleArshad, F., Sharma, A., Lu, C., & Gulia-Nuss, M. (2021). RNAi by Soaking Aedes aegypti Pupae in dsRNA. Insects, 12(7), 634. https://doi.org/10.3390/insects12070634