
Defensive Traits during White Spruce (Picea glauca) Leaf Ontogeny


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Literature Review on Phenology of Spruce Defensive Traits
2.1. Leaf Nutritional Traits
2.2. Phenolics
2.3. Terpenoids
2.4. Alkaloids
2.5. Toughness
3. Materials and Methods
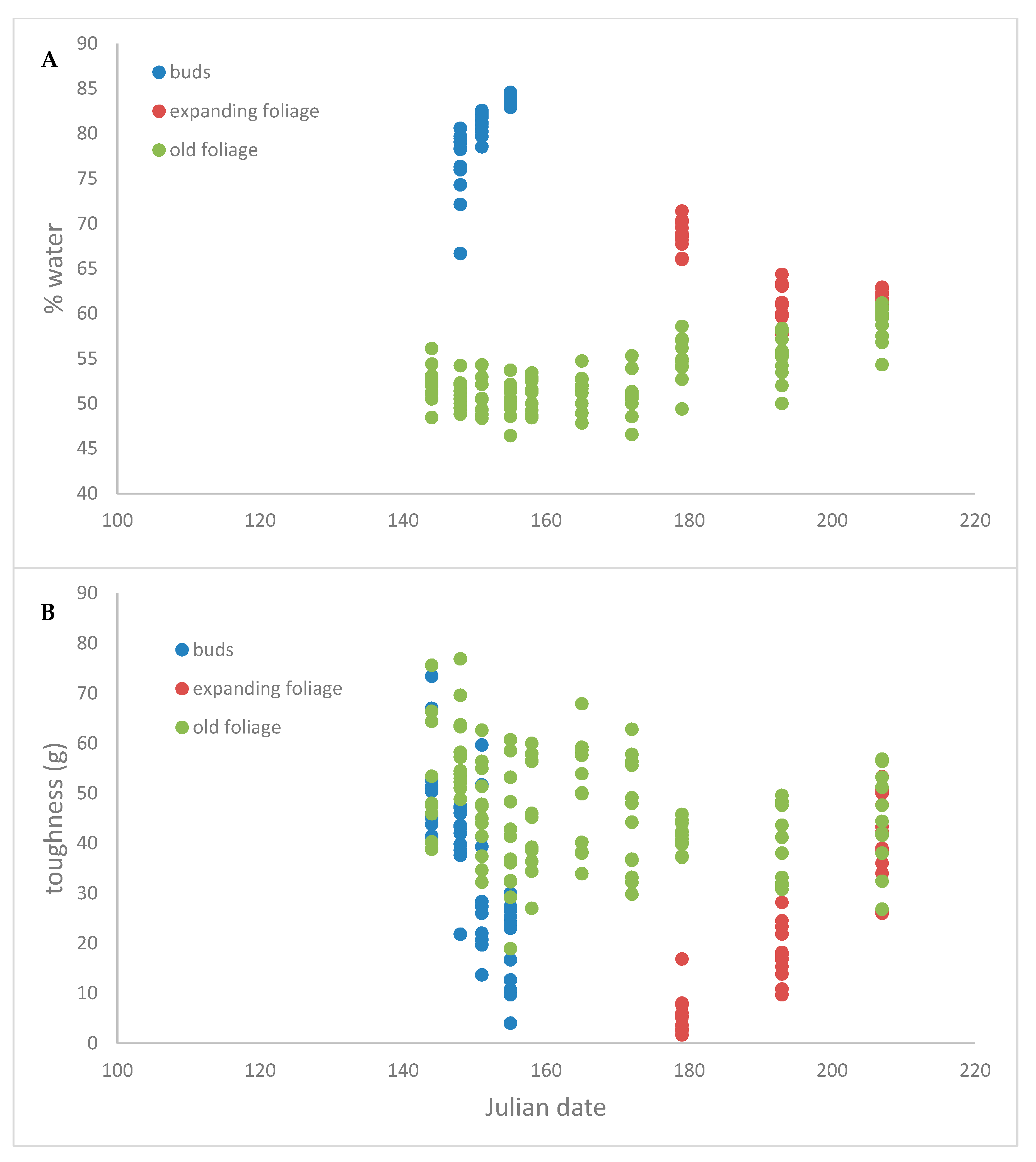
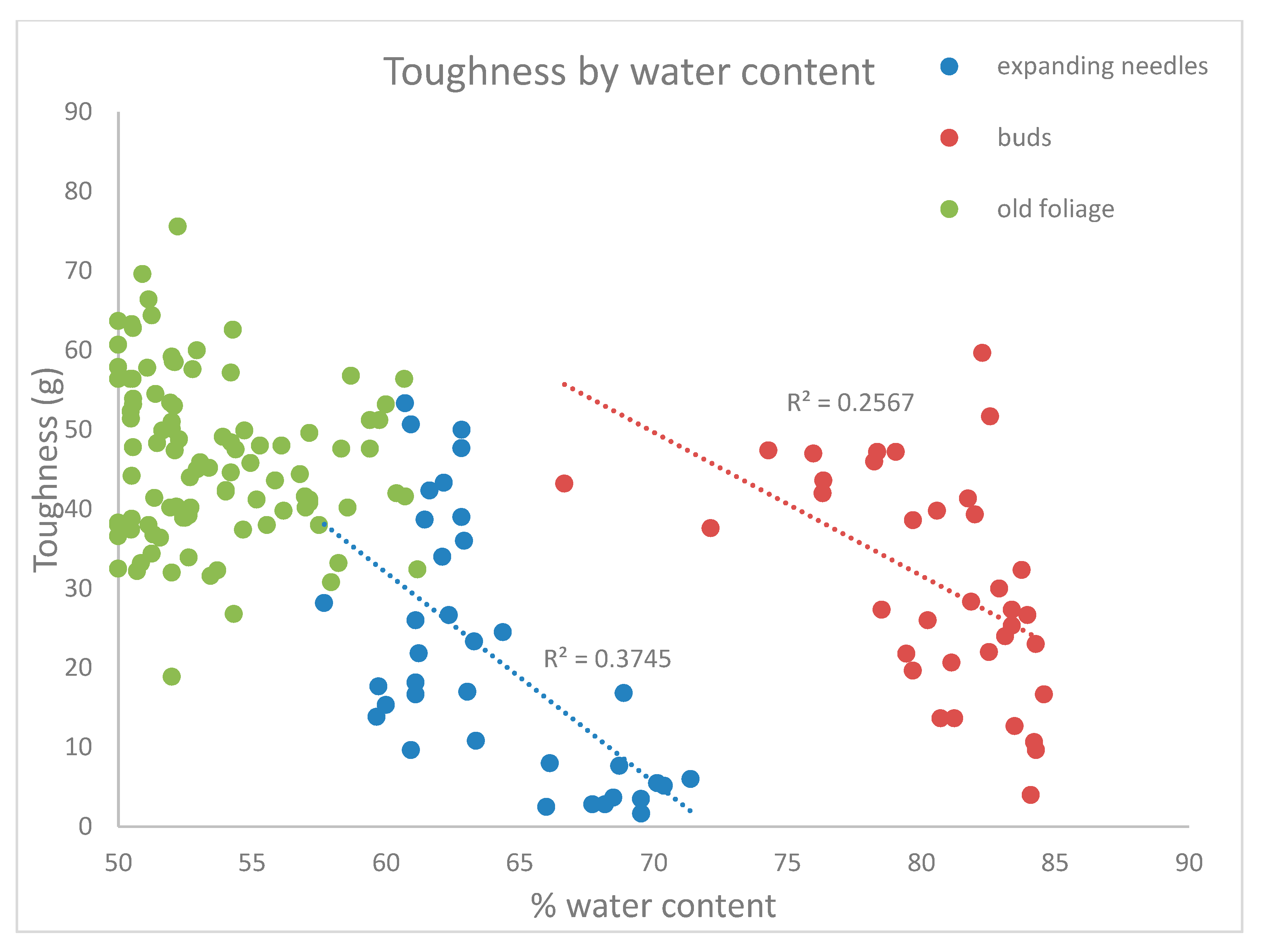
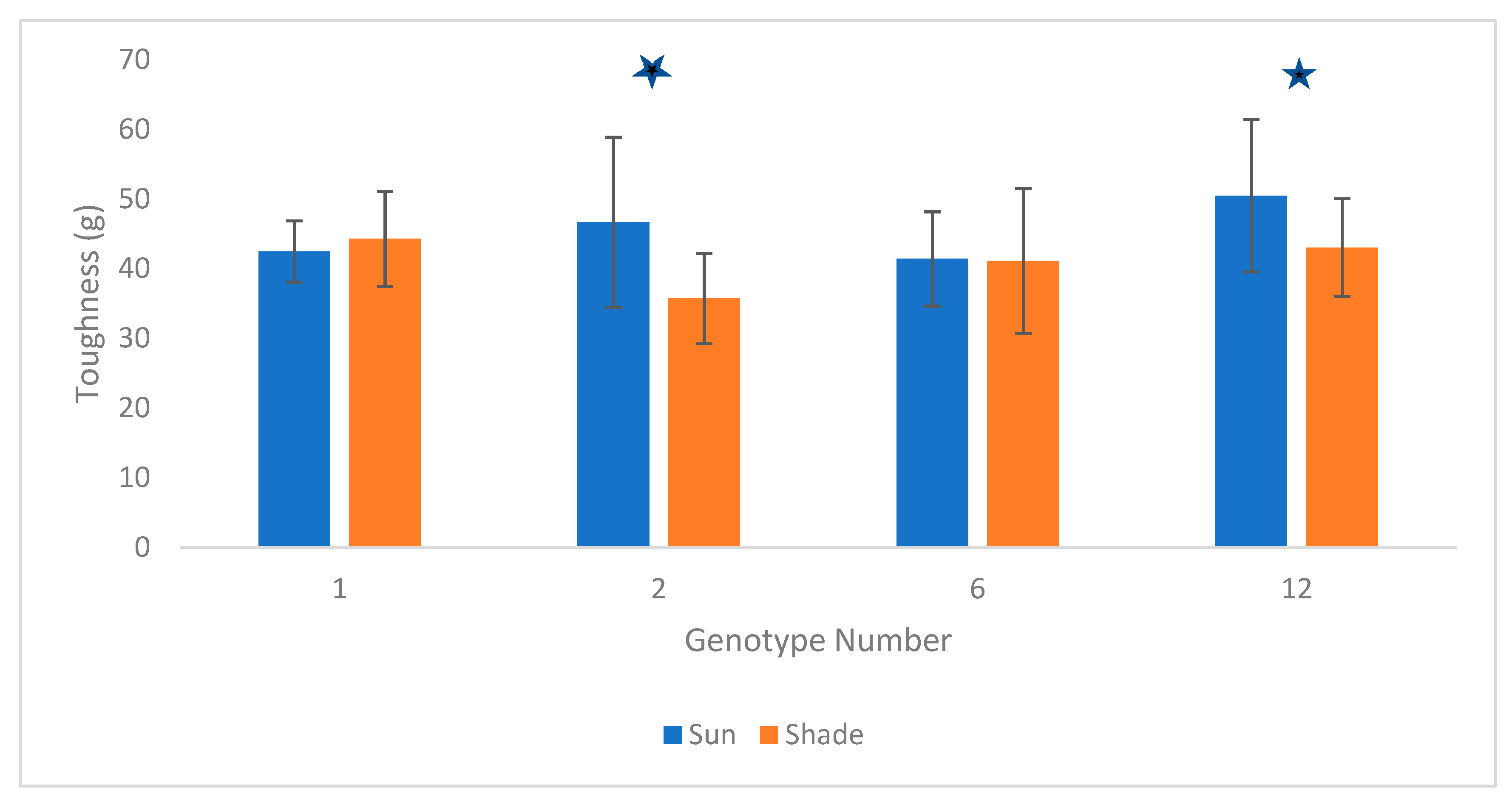
4. Results
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barton, K.E.; Edwards, K.F.; Koricheva, J. Shifts in Woody Plant Defence Syndromes during Leaf Development. Funct. Ecol. 2019, 33, 2095–2104. [Google Scholar] [CrossRef] [Green Version]
- Coley, P.D. Effects of Leaf Age and Plant Life History Patterns on Herbivory. Nature 1980, 284, 545–546. [Google Scholar] [CrossRef]
- Kursar, T.A.; Coley, P.D. Convergence in Defense Syndromes of Young Leaves in Tropical Rainforests. Biochem. Syst. Ecol. 2003, 31, 929–949. [Google Scholar] [CrossRef]
- Albersheim, P.; Darvill, A.; Roberts, K.; Sederoff, R.; Staehelin, A. Plant Cell Walls; Garland Science: New York, NY, USA, 2011. [Google Scholar]
- Esau, K. Anatomy of Seed Plants, 2nd ed.; Wiley: New York, NY, USA, 1977; ISBN 978-0-471-24520-9. [Google Scholar]
- Boege, K.; Marquis, R.J. Facing Herbivory as You Grow up: The Ontogeny of Resistance in Plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef]
- Carroll, A.L. Physiological Adaptation to Temporal Variation in Conifer Foliage by a Caterpillar. Can. Entomol. 1999, 131, 659–669. [Google Scholar] [CrossRef]
- Chen, Z.; Kolb, T.E.; Clancy, K.M. The Role of Monoterpenes in Resistance of Douglas Fir to Western Spruce Budworm Defoliation. J. Chem. Ecol. 2002, 28, 897–920. [Google Scholar] [CrossRef]
- Nealis, V. The Phenological Window for Western Spruce Budworm: Seasonal Decline in Resources Quality. Agric. For. Entomol. 2012, 14, 4. [Google Scholar] [CrossRef]
- Watt, A.D. The Effect of Shoot Growth Stage of Pinus contorta and Pinus sylvestris on the Growth and Survival of Panolis Flammea Larvae. Oecologia 1987, 72, 429–433. [Google Scholar] [CrossRef]
- Prado, A.; Sierra, A.; Windsor, D.M.; Bede, J.C. Leaf Traits and Herbivory Levels in a Tropical Gymnosperm, Zamia stevensonii (Zamiaceae). Am. J. Bot. 2014, 10, 437–447. [Google Scholar] [CrossRef] [Green Version]
- van Asch, M.; Visser, M.E. Phenology of Forest Caterpillars and Their Host Trees: The Importance of Synchrony. Annu. Rev. Entomol. 2007, 52, 37–55. [Google Scholar] [CrossRef]
- Feeny, P. Plant Apparency and Chemical Defense. In Biochemical Interaction Between Plants and Insects; Recent Advances in Phytochemistry; Wallace, J.W., Mansell, R.L., Eds.; Springer: Boston, MA, USA, 1976; pp. 1–40. ISBN 978-1-4684-2646-5. [Google Scholar]
- Hunter, A.F.; Lechowicz, M.J. Foliage Quality Changes during Canopy Development of Some Northern Hardwood Trees. Oecologia 1992, 89, 316–323. [Google Scholar] [CrossRef]
- Fuentealba, A.; Sagne, S.; Legendre, G.; Pureswaran, D.; Bauce, E.; Despland, E. Leaf Toughness as a Mechanism of Defense against Spruce Budworm. Arthropod Plant Interact. 2020, 14, 481–489. [Google Scholar] [CrossRef]
- Birol, I.; Raymond, A.; Jackman, S.; Pleasance, S.; Coope, R.; Taylor, G.; Yuen, M.M.S.; Keeling, C.; Brand, D.; Vandervalk, B.; et al. Assembling the 20 Gb White Spruce (Picea glauca) Genome from Whole-Genome Shotgun Sequencing Data. Bioinformatics 2013, 29, 1492–1497. [Google Scholar] [CrossRef]
- Hammerbacher, A.; Wright, L.P.; Gershenzon, J. Spruce Phenolics: Biosynthesis and Ecological Functions. In The Spruce Genome; Compendium of Plant Genomes; Porth, I.M., De la Torre, A.R., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 193–214. ISBN 978-3-030-21001-4. [Google Scholar]
- Keeling, C.I.; Bohlmann, J. Genes, Enzymes and Chemicals of Terpenoid Diversity in the Constitutive and Induced Defence of Conifers against Insects and Pathogens. New Phytol. 2006, 170, 657–675. [Google Scholar] [CrossRef]
- Parent, G.J.; Méndez-Espinoza, C.; Giguère, I.; Mageroy, M.H.; Charest, M.; Bauce, É.; Bohlmann, J.; MacKay, J.J. Hydroxyacetophenone Defenses in White Spruce against Spruce Budworm. Evol. Appl. 2020, 13, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.; Nadeau, S.; Ding, C.; Celedon, J.M.; Azaiez, A.; Ritland, C.; Laverdière, J.-P.; Deslauriers, M.; Adams, G.; Fullarton, M.; et al. Genomic Selection for Resistance to Spruce Budworm in White Spruce and Relationships with Growth and Wood Quality Traits. Evol. Appl. 2020, 13, 2704–2722. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, A.; Pureswaran, D.S.; Bauce, E.; Despland, E. How Does Synchrony with Host Plant Affect Spruce Budworm Performance? Oecologia 2017, 184, 847–857. [Google Scholar] [CrossRef]
- Lawrence, R.K.; Mattson, W.J.; Haack, R.A. White Spruce and the Spruce Budworm: Defining the Phenological Window of Susceptibility. Can. Entomol. 1997, 129, 291–318. [Google Scholar] [CrossRef]
- Quiring, D.T. Rapid Change in Suitability of White Spruce for a Specialist Herbivore, Zeiraphera canadensis, as a Function of Leaf Age. Can. J. Zool. 1992, 70, 2132–2138. [Google Scholar] [CrossRef]
- Fuentealba, A.; Sagne, S.; Pureswaran, D.; Bauce, É.; Despland, E. Defining the Window of Opportunity for Feeding Initiation by Second-Instar Spruce Budworm Larvae. Can. J. For. Res. 2018, 48, 285–291. [Google Scholar] [CrossRef]
- Millard, P.; Grelet, G. Nitrogen Storage and Remobilization by Trees: Ecophysiological Relevance in a Changing World. Tree Physiol. 2010, 30, 1083–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhuli, P.; Rohloff, J.; Strimbeck, G.R. Metabolite Changes in Conifer Buds and Needles during Forced Bud Break in Norway Spruce (Picea abies) and European Silver Fir (Abies alba). Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durzan, D.J. Nitrogen Metabolism of Picea Glauca. I. Seasonal Changes of Free Amino Acids in Buds, Shoot Apices, and Leaves, and the Metabolism of Uniformly Labelled 14C-L-Arginine by Buds during the Onset of Dormancy. Can. J. Bot. 1968, 46, 909–919. [Google Scholar] [CrossRef]
- Deslauriers, A.; Fournier, M.-P.; Cartenì, F.; Mackay, J. Phenological Shifts in Conifer Species Stressed by Spruce Budworm Defoliation. Tree Physiol. 2019, 39, 590–605. [Google Scholar] [CrossRef]
- Wiley, R.G.; Helliker, A. Re-evaluation of Carbon Storage in Trees Lends Greater Support for Carbon Limitation to Growth. New Phytol. 2012, 195, 285–289. [Google Scholar] [CrossRef]
- Heinrich, S.; Dippold, M.A.; Werner, C.; Wiesenberg, G.L.; Kuzyakov, Y.; Glaser, B. Allocation of Freshly Assimilated Carbon into Primary and Secondary Metabolites after in Situ 13C Pulse Labelling of Norway Spruce (Picea abies). Tree Physiol. 2015, 35, 1176–1191. [Google Scholar] [PubMed]
- Despland, E. Effects of Phenological Synchronization on Caterpillar Early-Instar Survival under a Changing Climate. Can. J. For. Res. 2018, 58, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Mattson, W.J.; Haack, R.A.; Lawrence, R.K.; Slocum, S.S. Considering the Nutritional Ecology of the Spruce Budworm and Its Management. For. Ecol. Manag. 1991, 39, 83–210. [Google Scholar] [CrossRef]
- Salminen, J.P.; Karonen, M. Chemical Ecology of Tannins and Other Phenolics: We Need a Change in Approach. Funct. Ecol. 2011, 25, 325–338. [Google Scholar] [CrossRef]
- Mumm, R.; Hilker, M. Direct and Indirect Chemical Defence of Pine against Folivorous Insects. Trends Plant Sci. 2006, 11, 351–358. [Google Scholar] [CrossRef]
- Delvas, N.; Bauce, E.; Labbé, C.; Ollevier, T.; Bélanger, R. Phenolic Compounds That Confer Resistance to Spruce Budworm. Entomol. Exp. Appl. 2011, 141, 35–44. [Google Scholar] [CrossRef]
- Parent, G.; Giguere, I.; Germanos, G.; Lamara, M.; Bauce, E.; MacKay, J.J. Insect Herbivory (Choristoneura fumiferana, Tortricidea) Underlies Tree Population Structure (Picea glauca, Pinaceae). Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mageroy, M.H.; Parent, G.; Germanos, G.; Giguere, I.; Delvas, N.; Maaroufi, H.; Bauce, E.; Bohlmann, J.; Mackay, J. Expression of the Beta-Glucosidase Gene Pg-Glu-1 Underpins Natural Resistance of White Spruce against Spruce Budworm. Plant J. 2014, 81, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, R.I. An Induced Defense Reaction in White Spruce to Attack by the White Pine Weevil, Pissodes strobi. Can. J. For. Res. 1995, 25, 1725–1730. [Google Scholar] [CrossRef]
- Huber, D.P.; Philippe, R.N.; Madilao, L.L.; Sturrock, R.N.; Bohlmann, J. Changes in Anatomy and Terpene Chemistry in Roots of Douglas-Fir Seedlings Following Treatment with Methyl Jasmonate. Tree Physiol. 2005, 25, 1075–1083. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Gershenzon, J.; Bohlmann, J. Induction of Volatile Terpene Biosynthesis and Diurnal Emission by Methyl Jasmonate in Foliage of Norway Spruce. Plant Physiol. 2003, 132, 1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cates, R.G.; Henderson, C.B.; Redak, R.A. Responses of the Western Spruce Budworm to Varying Levels of Nitrogen and Terpenes. Oeocologia 1987, 73, 312–316. [Google Scholar] [CrossRef]
- Ikeda, T.; Matsumura, F.; Benjamin, D.M. Chemical Basis for Feeding Adaptation of Pine Sawflies Neodiprion rugions and Neodiprion swainei. Science 1977, 197, 497–498. [Google Scholar] [CrossRef]
- Nealis, V.G.; Nault, J.R. Seasonal Changes in Foliar Terpenes Indicate Suitability of Douglas-Fir Buds for Western Spruce Budworm. J. Chem. Ecol. 2005, 31, 683–696. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The Function of Terpene Natural Products in the Natural World. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Litvak, M.A.; Monson, R.K. Patterns of Induced and Constitutive Monoterpene Production in Conifer Needles in Relation to Insect Herbivory. Oecologia 1998, 114, 531–540. [Google Scholar] [CrossRef]
- Miller, B.; Madilao, L.L.; Ralph, S.; Bohlmann, J. Insect-Induced Conifer Defense. White Pine Weevil and Methyl Jasmonate Induce Traumatic Resinosis, de Novo Formed Volatile Emissions, and Accumulation of Terpenoid Synthase and Putative Octadecanoid Pathway Transcripts in Sitka Spruce. Plant Physiol. 2005, 137, 369–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Despland, E.; Bourdier, T.; Dion, E.; Bauce, E. Do White Spruce Epicuticular Was Monoterpenes Follow Foliar Patterns? Can. J. For. Res. 2016, 46, 1051–1058. [Google Scholar] [CrossRef] [Green Version]
- Rudloff, E.V. Seasonal Variation in the Composition of the Volatile Oil of the Leaves, Buds, and Twigs of White Spruce (Picea glauca). Can. J. Bot. 1972, 50, 1595–1603. [Google Scholar] [CrossRef]
- Thoss, V.; O’Reilly-Wapstra, J.; Iason, G.R. Assessment and Implications of Intraspecific and Phenological Variability in Monoterpenes of Scots Pine (Pinus sylvestris) Foliage. J. Chem. Ecol. 2007, 33, 477–491. [Google Scholar] [CrossRef]
- Hall, D.E.; Robert, J.A.; Keeling, C.I.; Domanski, D.; Quesada, A.L.; Jancsik, S.; Kuzyk, M.A.; Hamberger, B.; Borchers, C.H.; Bohlmann, J. An Integrated Genomic, Proteomic and Biochemical Analysis of (+)-3-Carene Biosynthesis in Sitka Spruce (Picea sitchensis) Genotypes That Are Resistant or Susceptible to White Pine Weevil. Plant J. 2011, 65, 939–948. [Google Scholar] [CrossRef]
- Roach, C.R.; Hall, D.E.; Zerbe, P.; Bohlmann, J. Plasticity and Evolution of (+)-3-Carene Synthase and (-)-Sabinene Synthase Functions of a Sitka Spruce Monoterpene Synthase Gene Family Associated with Weevil Resistance. J. Biol. Chem. 2014, 289, 23859–23869. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.J.; Montali, J.A.; Hazen, D.; Stanton, C.E. Alkaloids of Picea. J. Nat. Prod. 1991, 54, 905–909. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Kamm, C.D.; Tawara, J.N. Piperidine Alkaloids of Spruce (Picea) and Fir (Abies) Species. Biochem. Syst. Ecol. 2000, 28, 177–181. [Google Scholar] [CrossRef]
- Kamm, C.D.; Tawara, J.N.; Stermitz, F.R. Spruce Budworm Larval Processing of Piperidine Alkaloids from Spruce Needles. J. Chem. Ecol. 1998, 24, 1153–1160. [Google Scholar] [CrossRef]
- Zhao, B.; Grant, G.G.; Langevin, D.; MacDonald, L. Deterring and Inhibiting Effects of Quinolizidine Alkaloids on Spruce Budworm (Lepidoptera: Tortricidae) Oviposition. Environ. Entomol. 1998, 27, 984–992. [Google Scholar] [CrossRef]
- Virjamo, V.; Julkunen-Tiitto, R. Shoot Development of Norway Spruce (Picea abies) Involves Changes in Piperidine Alkaloids and Condensed Tannins. Trees 2014, 28, 427–437. [Google Scholar] [CrossRef]
- Arimura, G.; Maffei, M. Plant Specialized Metabolism: Genomics, Biochemistry, and Biological Functions; CRC Press: Boca Raton, FL, USA, 2016; ISBN 978-1-315-35337-1. [Google Scholar]
- Zidorn, C. Seasonal Variation of Natural Products in European Trees. Phytochem. Rev. 2018, 17, 923–935. [Google Scholar] [CrossRef]
- Juniper, B.; Southwood, R. Insects and the Plant Surface; Arnold, E., Ed.; Edward Arnold: Baltimore, MD, USA, 1986; p. 360. ISBN 0-7131-2909-3. [Google Scholar]
- Coley, P.D. Herbivory and Defensive Characteristics of Tree Species in a Lowland Tropical Forest. Ecol. Monogr. 1983, 53, 209–234. [Google Scholar] [CrossRef]
- Lowman, M.; Box, J.D. Variation in Leaf Toughness and Phenolic Content among Five Species of Australian Rain Forest Trees. Aust. J. Ecol. 1983, 8, 17–25. [Google Scholar] [CrossRef]
- Clissold, F. The biomechanics of chewing and plant fracture: Mechanisms and implications. In Advances in Insect Physiology: Insect Mechanics and Control; Casas, J., Simpson, S.J., Eds.; Academic Press: London, UK, 2008; Volume 34, pp. 317–372. [Google Scholar]
- Hochuli, D.F. Insect Herbivory and Ontogeny: How Do Growth and Development Influence Feeding Behaviour, Morphology and Host Use? Austral Ecol. 2001, 26, 563–570. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Clarke, A.R.; Malcolm, S.B. Ecology and Behavior of First Instar Larval Lepidoptera. Annu. Rev. Entomol. 2002, 47, 361–393. [Google Scholar] [CrossRef] [PubMed]
- Pinault, L.; Thurston, G.; Quiring, D. Interaction of Foliage and Larval Age Influences Preference and Performance of a Geometrid Moth. Can. Entomol. 2009, 141, 136–144. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Overton, C.; Stewart, D.; Rutledge, R.G. Initiation of Somatic Embryos and Regeneration of Plants from Primordial Shoots of 10-Year-Old Somatic White Spruce and Expression Profiles of 11 Genes Followed during the Tissue Culture Process. Planta 2011, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Dorais, L.G.; Kettala, E. Revue, Par Région, Des Techniques d’inventaire Entomologique et d’évaluation Des Programmes de Pulvérisation à Grande Échelle Contre La Tordeuse Des Bourgeons de l’épinette Choristoneura Fumiferana (Clem.); Ministère de l’Énergie et des Ressources du Québec: Québec, QC, Canada, 1982; p. 51. [Google Scholar]
- Dhont, C.; Sylvestre, P.; Gros-Louis, M.C.; Isabel, N. Guide-Terrain Pour l’identifcation Des Stades de Débourrement et de Formation Du Bourgeon Apical Chez l’épinette Blanche; Natural Resources Canada, Laurentian Forestry Centre: Ste-Foy, QC, Canada, 2010. [Google Scholar]
- Donkor, D.; Mirzahosseini, Z.; Bede, J.; Bauce, E.; Despland, E. Detoxification of Host Plant Phenolic Aglycones by the Spruce Budworm. PLoS ONE 2019, 14, e0208288. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, K.; Llorens, A.M.; Stefanescu, C.; Timchenko, M.V.; Lucas, P.W.; Wright, S.J. How Cellulose-Based Leaf Toughness and Lamina Density Contribute to Long Leaf Lifespans of Shade-Tolerant Species. New Phytol. 2012, 195, 640–652. [Google Scholar] [CrossRef]
- Lucas, P.W.; Turner, I.M.; Dominy, N.J.; Yamashita, N. Mechanical Defences to Herbivory. Ann. Bot. 2000, 86, 913–920. [Google Scholar] [CrossRef]
- Marco, H.F. The Anatomy of Spruce Needles. J. Agric. Res. 1939, 58, 357–368. [Google Scholar]
- Coley, P.D.; Lokvam, J.; Rudolph, K.; Bromberg, K.; Sackett, T.E.; Wright, L.; Brenes-Arguedas, T.; Dvorett, D.; Ring, S.; Clark, A.; et al. Divergent Defensive Strategies of Young Leaves in Two Species of Inga. Ecology 2005, 86, 2633–2643. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, J.R.; Lindroth, R.L. Genetics, environment, and their interaction determine efficacy of chemical defense in trembling aspen. Ecology 2007, 88, 729–739. [Google Scholar] [CrossRef]
- Nealis, V.G.; Régnière, J. Insect-Host Relationships Influencing Disturbance by the Spruce Budworm in a Boreal Mixedwood Forest. Can. J. For. Res. 2004, 34, 1870–1882. [Google Scholar] [CrossRef] [Green Version]
- Blais, J.R. Trends in the Frequency, Extent, and Severity of Spruce Budworm Outbreaks in Eastern Canada. Can. J. Zool. 1983, 13, 539–547. [Google Scholar] [CrossRef]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex Responses of Global Insect Pests to Climate Warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Bellemin-Noel, B.; Bourassa, S.; Despland, E.; De Grandpré, L.; Pureswaran, D. Improved Performance of the Eastern Spruce Budworm on Black Spruce as Warming Temperatures Disrupt Phenological Defenses. Glob. Chang. Biol. 2021. [Google Scholar] [CrossRef]
- Ekholm, A.; Tack, A.J.M.; Pulkkinen, P.; Roslin, T. Host Plant Phenology, Insect Outbreaks and Herbivore Communities—The Importance of Timing. J. Anim. Ecol. 2019, 89, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Forrest, J.; Miller-Rushing, A.J. Toward a Synthetic Understanding of the Role of Phenology in Ecology and Evolution. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3101–3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albert, L.P.; Restrepo-Coupe, N.; Smith, M.N.; Wu, J.; Chavana-Bryant, C.; Prohaska, N.; Taylor, T.C.; Martins, G.A.; Ciais, P.; Mao, J.; et al. Cryptic Phenology in Plants: Case Studies, Implications, and Recommendations. Glob. Chang. Biol. 2019, 25, 3591–3608. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lirette, A.-O.; Despland, E. Defensive Traits during White Spruce (Picea glauca) Leaf Ontogeny. Insects 2021, 12, 644. https://doi.org/10.3390/insects12070644
Lirette A-O, Despland E. Defensive Traits during White Spruce (Picea glauca) Leaf Ontogeny. Insects. 2021; 12(7):644. https://doi.org/10.3390/insects12070644
Chicago/Turabian StyleLirette, Antoine-Olivier, and Emma Despland. 2021. "Defensive Traits during White Spruce (Picea glauca) Leaf Ontogeny" Insects 12, no. 7: 644. https://doi.org/10.3390/insects12070644
APA StyleLirette, A. -O., & Despland, E. (2021). Defensive Traits during White Spruce (Picea glauca) Leaf Ontogeny. Insects, 12(7), 644. https://doi.org/10.3390/insects12070644