
Stage- and Rearing-Dependent Metabolomics Profiling of Ophiocordyceps sinensis and Its Pipeline Products



Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals
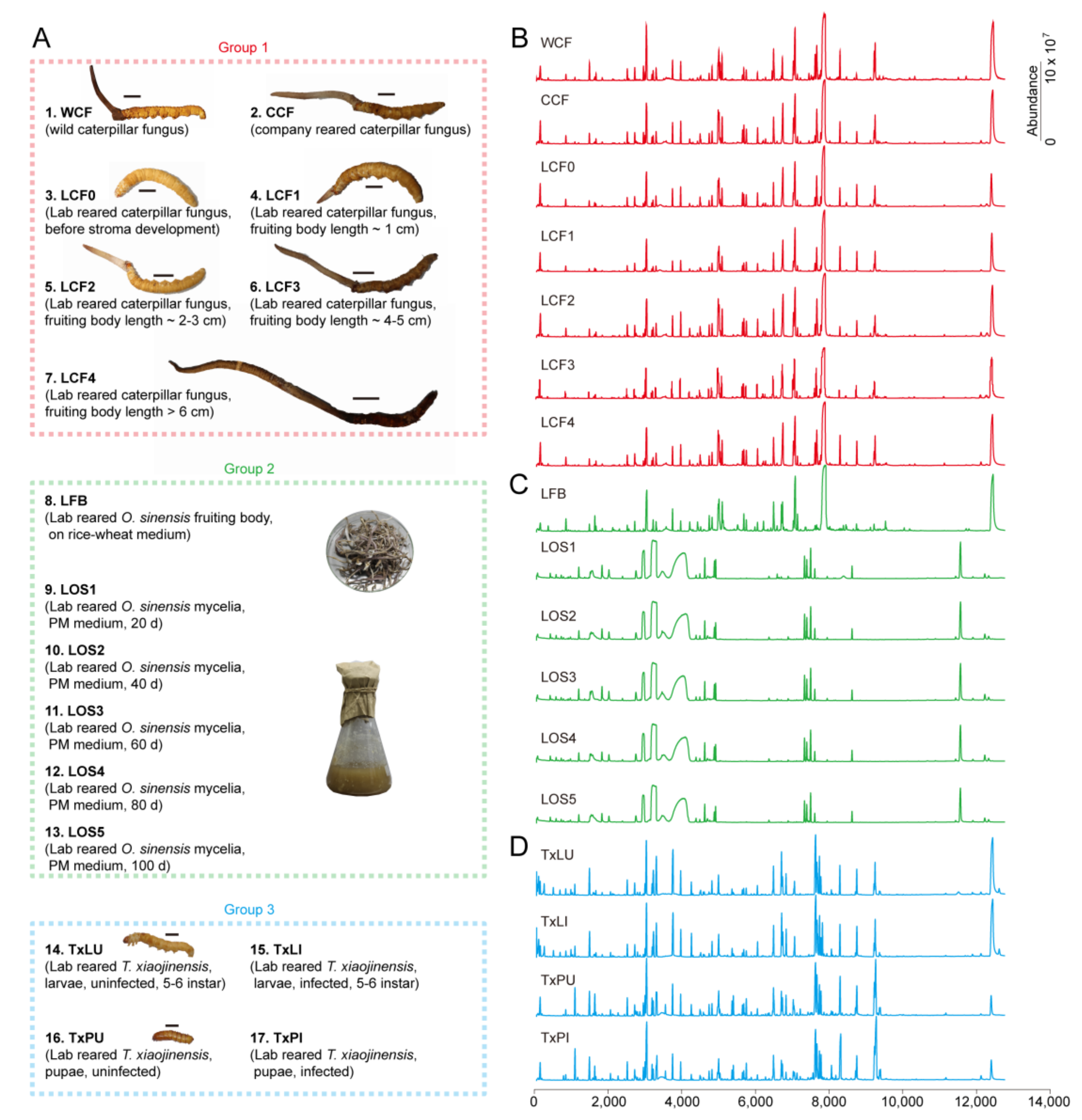
2.2. Materials
2.3. Sample Preparation
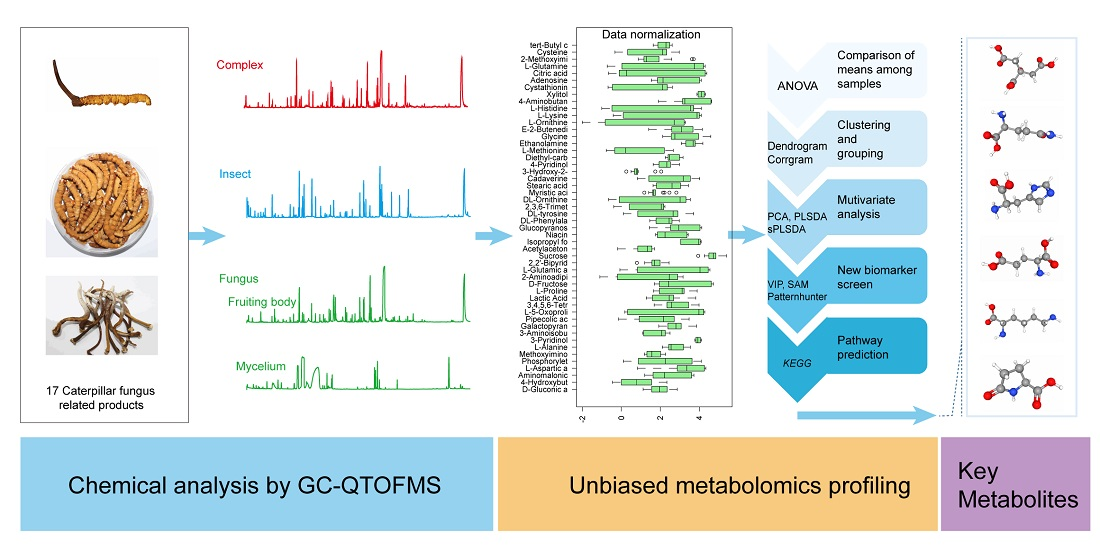
2.4. Chemical Analysis
2.5. Data Processing
3. Results
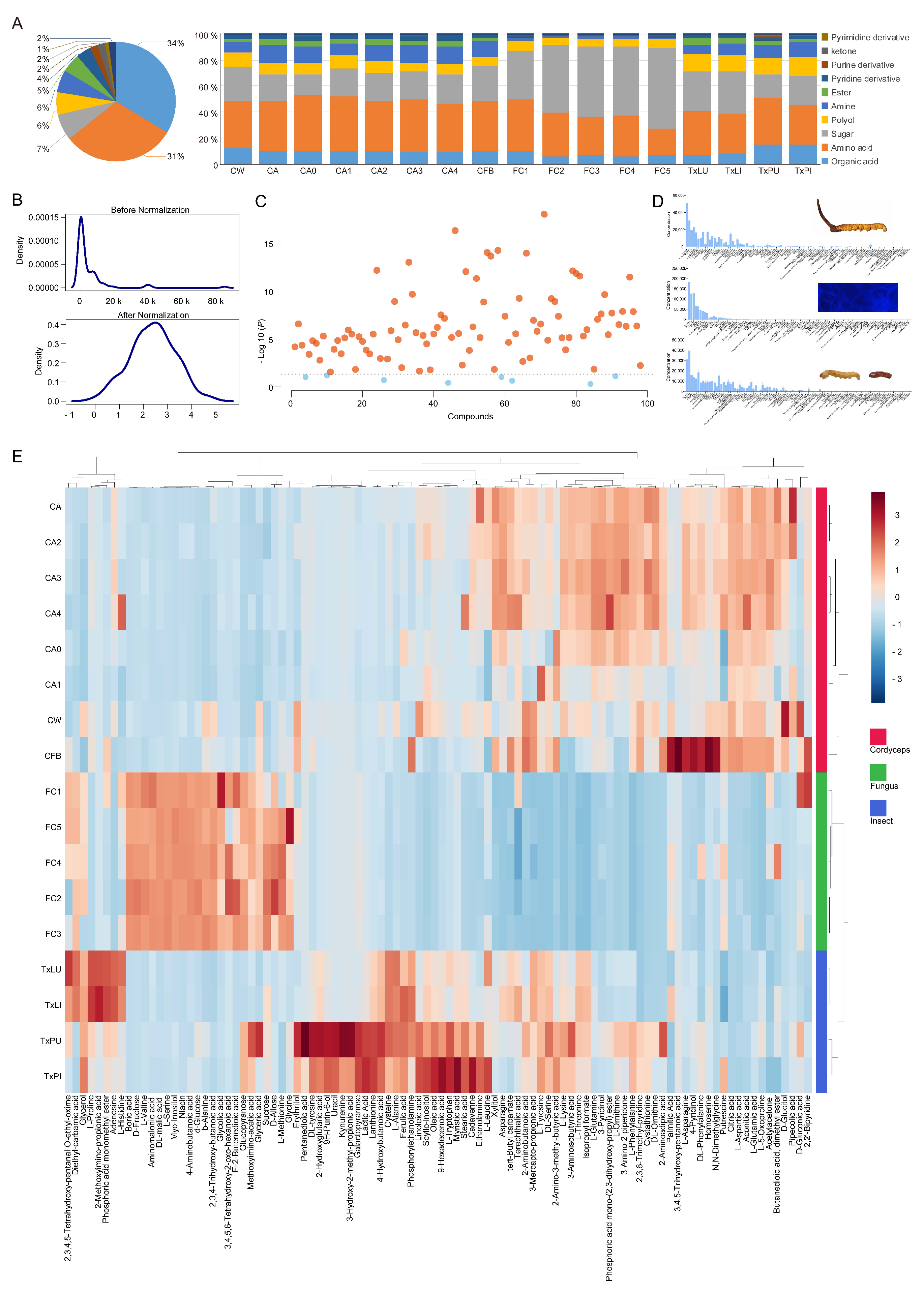
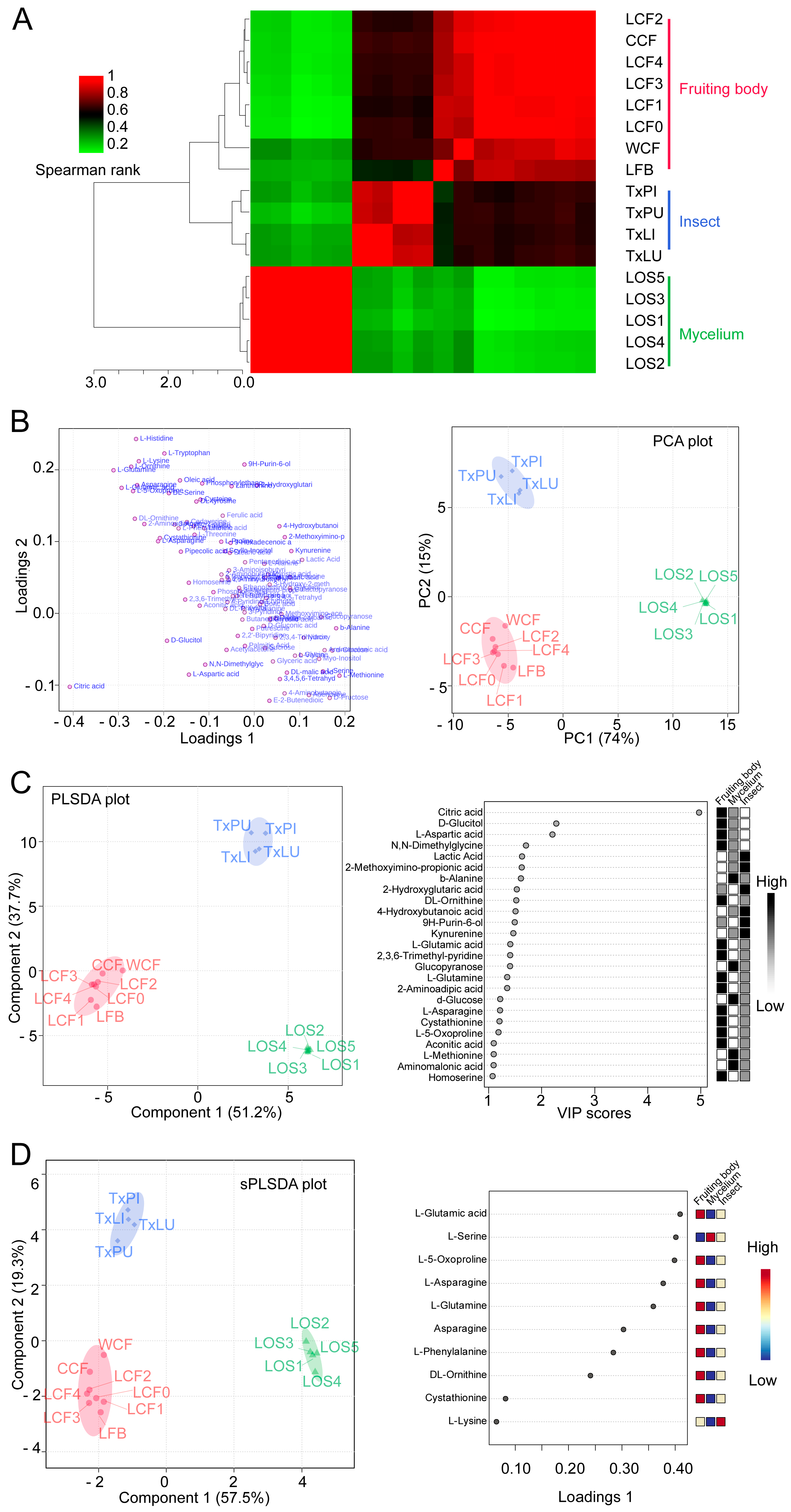
3.1. Metabolic Profiles and Overall Assessment
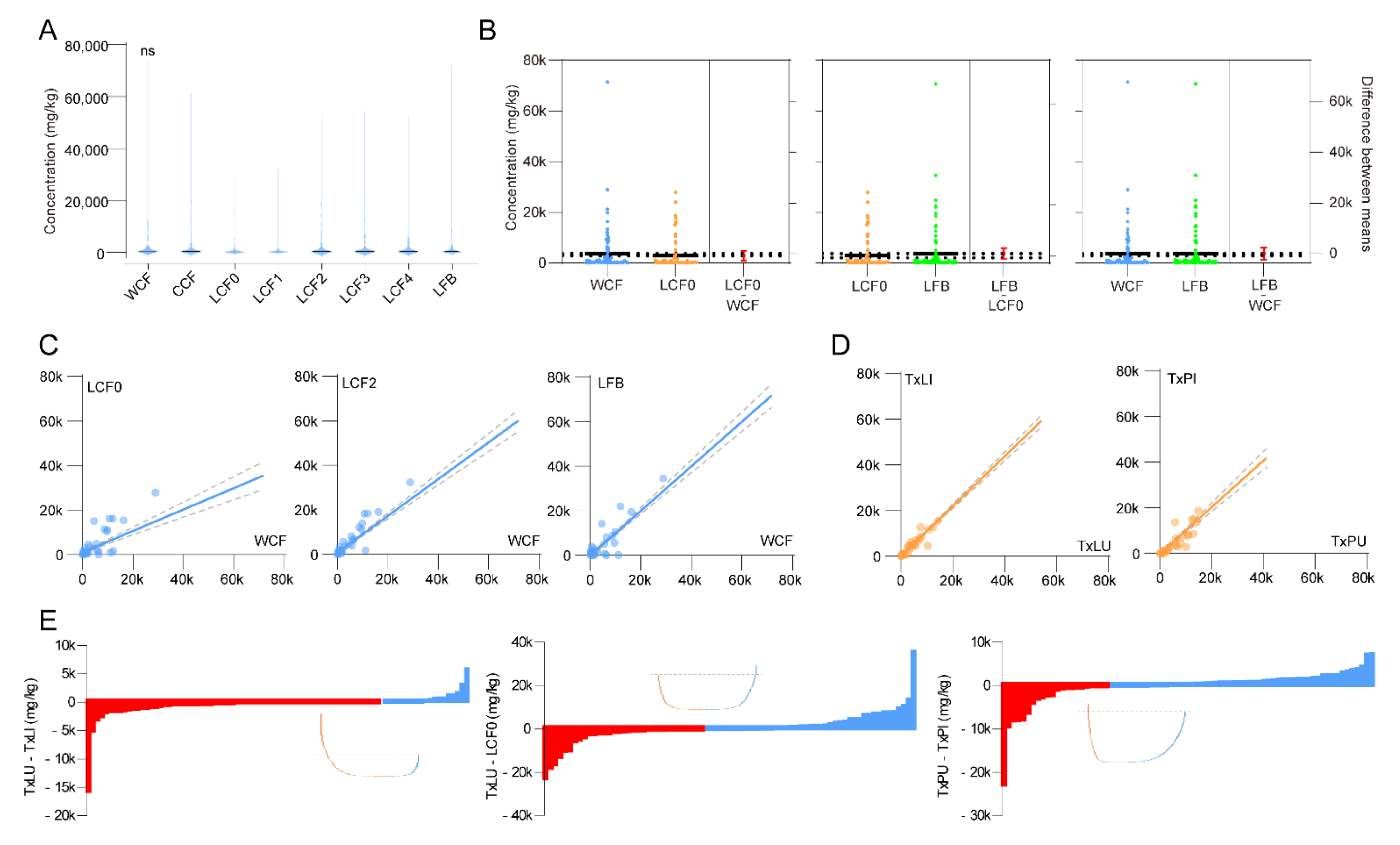
3.2. Stage, Spatial and Infection Effects
3.3. Clustering and Classification of Treatments
3.4. Selection of Featured Metabolites and Characterizations
4. Discussion
4.1. Situation Dependent Metabolomics of O. sinensis
4.2. Featured Metabolites and Their Potential Roles as New Quality Control Biomarkers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.L. 1757, Ben Cao Cong Xin (New Compilation of Materia Medica). Qing Dynasty of China: Li Ji Tang Ben. Available online: http://ctext.org/wiki.pl?If=gb&chapter=231100 (accessed on 30 September 2020).
- Isaka, M.; Kittakoop, P.; Thebtaranonth, Y. Secondary Metabolites of Clavicipitalean Fungi. In Clavicipitalean Fungi: Evolutionary Biology, Chemistry, Biocontrol, and Cultural Impacts; White, J.F., Jr., Bacon, C.W., Hywel-Jones, N.L., Spatafora, J.W., Eds.; Marcel Dekker: New York, NY, USA, 2005; pp. 331–372. [Google Scholar] [CrossRef]
- Han, R.; Wu, H.; Tao, H.; Qiu, X.; Liu, G.; Rao, Z.; Cao, L. Research on Chinese cordyceps during the past 70 years in China. Chin. J. Appl. Entomol. 2019, 56, 849–883. [Google Scholar] [CrossRef]
- Sun, Y.D.; Zhang, Y.; Yang, P.Y.; Jia, Y.J. The effect of Cordyceps sinensis in the prognosis on patients receiving chemotherapy with malignant tumors: A systematic review and meta-analysis. TMR Cancer 2020, 3, 19–27. [Google Scholar] [CrossRef]
- Bok, J.W.; Lermer, L.; Chilton, J.; Klingeman, H.G.; Towers, G. Antitumor sterols from the mycelia of Cordyceps sinensis. Phytochemistry 1999, 51, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Gong, Z.; Su, Y.; Lin, J.; Tang, K. Cordyceps fungi: Natural products, pharmacological functions and developmental products. J. Pharm. Pharmacol. 2009, 61, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-L.; Kuo, P.-C.; Hwang, T.-L.; Wu, T.-S. Anti-inflammatory Principles fromCordyceps sinensis. J. Nat. Prod. 2011, 74, 1996–2000. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Siu, K.-C.; Wang, W.-Q.; Liu, X.-X.; Wu, J.-Y. Structure and antioxidant activity of a novel poly-N-acetylhexosamine produced by a medicinal fungus. Carbohydr. Polym. 2013, 94, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-Q.; Song, A.-X.; Yin, J.-Y.; Siu, K.-C.; Wong, W.-T.; Wu, J.-Y. Anti-inflammation activity of exopolysaccharides produced by a medicinal fungus Cordyceps sinensis Cs-HK1 in cell and animal models. Int. J. Biol. Macromol. 2020, 149, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Zhou, X.; Wang, J.; Zhang, K.; Zhou, Y.; Chen, S.; Nie, S.; Xie, M. Cordyceps sinensis polysaccharide inhibits colon cancer cells growth by inducing apoptosis and autophagy flux blockage via mTOR signaling. Carbohydr. Polym. 2020, 237, 116113. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Guo, S.; Wang, W.; Liu, X. Cordyceps industry in China. Mycol. 2015, 6, 121–129. [Google Scholar] [CrossRef]
- Wang, Z.; Li, M.; Ju, W.; Ye, W.; Xue, L.; Boufford, D.E.; Gao, X.; Yue, B.; Liu, Y.; Pierce, N.E. The entomophagous caterpillar fungus Ophiocordyceps sinensis is consumed by its lepidopteran host as a plant endophyte. Fungal Ecol. 2020, 47, 100989. [Google Scholar] [CrossRef]
- Collin, M.A.; Mita, K.; Sehnal, F.; Hayashi, C.Y. Molecular Evolution of Lepidopteran Silk Proteins: Insights from the Ghost Moth, Hepialus californicus. J. Mol. Evol. 2010, 70, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, Z.; Cao, L.; Qiu, X.; Han, R. Comparative transcriptome analysis reveals molecular strategies of ghost moth Thitarodes armoricanus in response to hypoxia and anoxia. J. Insect Physiol. 2019, 112, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Q.; Li, W.; Li, Q.; Qian, Z.; Liu, X.; Dong, C. A breakthrough in the artificial cultivation of Chinese cordyceps on a large-scale and its impact on science, the economy, and industry. Crit. Rev. Biotechnol. 2018, 39, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Hopping, K.A.; Chignell, S.M.; Lambin, E.F. The demise of caterpillar fungus in the Himalayan region due to climate change and overharvesting. Proc. Natl. Acad. Sci. USA 2018, 115, 11489–11494. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Li, Y.; Wang, W.-J.; He, J.-S.; Yang, R.-H.; Wu, H.-J.; Wang, X.-L.; Jiao, L.; Tang, Z.; Yao, Y.-J. Range shifts in response to climate change of Ophiocordyceps sinensis, a fungus endemic to the Tibetan Plateau. Biol. Conserv. 2017, 206, 143–150. [Google Scholar] [CrossRef]
- Shrestha, U.B.; Shrestha, S.; Ghimire, S.; Nepali, K.; Shrestha, B.B. Chasing Chinese Caterpillar Fungus (Ophiocordyceps sinensis) Harvesters in the Himalayas: Harvesting Practice and Its Conservation Implications in Western Nepal. Soc. Nat. Resour. 2014, 27, 1242–1256. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, L.; Wang, J.; Wang, W.; Niyati, N.; Guo, Y.; Wang, X. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total. Environ. 2021, 755, 142548. [Google Scholar] [CrossRef]
- Yan, J.-K.; Wang, W.-Q.; Wu, J.-Y. Recent advances in Cordyceps sinensis polysaccharides: Mycelial fermentation, isolation, structure, and bioactivities: A review. J. Funct. Foods 2014, 6, 33–47. [Google Scholar] [CrossRef]
- Cao, L.; Han, R. Method for Artificial Cultivation of Ophiocordyceps sinensis Fruiting Bodies. U.S. Patent Application No. 10,400,209, 3 September 2019. [Google Scholar]
- Cao, L.; Ye, Y.; Han, R. Fruiting Body Production of the Medicinal Chinese Caterpillar Mushroom, Ophiocordyceps sinensis (Ascomycetes), in Artificial Medium. Int. J. Med. Mushrooms 2015, 17, 1107–1112. [Google Scholar] [CrossRef]
- Liu, G.; Han, R.; Cao, L. Artificial Cultivation of the Chinese Cordyceps From Injected Ghost Moth Larvae. Environ. Èntomol. 2019, 48, 1088–1094. [Google Scholar] [CrossRef]
- Mi, J.; Han, Y.; Xu, Y.; Kou, J.; Li, W.-J.; Wang, J.-R.; Jiang, Z.-H. Deep Profiling of Immunosuppressive Glycosphingolipids and Sphingomyelins in Wild Cordyceps. J. Agric. Food Chem. 2018, 66, 8991–8998. [Google Scholar] [CrossRef]
- Khakimov, B.; Jespersen, B.M.; Engelsen, S.B. Comprehensive and Comparative Metabolomic Profiling of Wheat, Barley, Oat and Rye Using Gas Chromatography-Mass Spectrometry and Advanced Chemometrics. Foods 2014, 3, 569–585. [Google Scholar] [CrossRef] [Green Version]
- Dona, A.; Kyriakides, M.; Scott, F.; Shephard, E.; Varshavi, D.; Veselkov, K.; Everett, J.R. A guide to the identification of metabolites in NMR-based metabonomics/metabolomics experiments. Comput. Struct. Biotechnol. J. 2016, 14, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhong, X.; Li, S.; Zhang, G.; Liu, X. Metabolic characterization of natural and cultured Ophicordyceps sinensis from different origins by 1H NMR spectroscopy. J. Pharm. Biomed. Anal. 2015, 115, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Gu, L.; Xiong, W.-T.; Wang, H.-Z.; Lian, D.-H.; Zheng, Y.-M.; Zhou, S.; Zhou, W.; Gu, J.-L.; Shen, J.-H.; et al. 1H NMR spectroscopy-based metabolic profiling of Ophiocordyceps sinensis and Cordyceps militaris in water-boiled and 50% ethanol-soaked extracts. J. Pharm. Biomed. Anal. 2020, 180, 113038. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; Mi, J.; Zhang, M.; Wang, Y.; Jiang, Z.; Hu, P. GC-MS Profiling of Volatile Components in Different Fermentation Products of Cordyceps Sinensis Mycelia. Molecules. 2017, 22, 1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, X.; Cao, L.; Han, R. Analysis of Volatile Components in Different Ophiocordyceps sinensis and Insect Host Products. Molecules. 2020, 25, 1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.-Q.; Feng, K.; Zhao, J.; Li, S. Analysis of sterols and fatty acids in natural and cultured Cordyceps by one-step derivatization followed with gas chromatography–mass spectrometry. J. Pharm. Biomed. Anal. 2009, 49, 1172–1178. [Google Scholar] [CrossRef]
- Guo, L.-X.; Xu, X.-M.; Liang, F.-R.; Yuan, J.-P.; Peng, J.; Wu, C.-F.; Wang, J.-H. Morphological Observations and Fatty Acid Composition of Indoor-Cultivated Cordyceps sinensis at a High-Altitude Laboratory on Sejila Mountain, Tibet. PLoS ONE 2015, 10, e0126095. [Google Scholar] [CrossRef]
- Cheong, K.L.; Wang, L.-Y.; Wu, D.-T.; Hu, D.-J.; Zhao, J.; Li, S.-P. Microwave-Assisted Extraction, Chemical Structures, and Chain Conformation of Polysaccharides from a Novel Cordyceps Sinensis Fungus UM01. J. Food Sci. 2016, 81, C2167–C2174. [Google Scholar] [CrossRef]
- Cao, L.; Han, R. Artificial Feeding Method at Low Altitude for Host Insect Ghost Moth of Ophiocordyceps sinensis. U.S. Patent Application No. 10,412,941, 17 September 2019. [Google Scholar]
- Quan, Q.-M.; Chen, L.-L.; Wang, X.; Li, S.; Yang, X.-L.; Zhu, Y.-G.; Wang, M.; Cheng, Z. Genetic Diversity and Distribution Patterns of Host Insects of Caterpillar Fungus Ophiocordyceps sinensis in the Qinghai-Tibet Plateau. PLoS ONE 2014, 9, e92293. [Google Scholar] [CrossRef] [Green Version]
- Souza-Silva, E.A.; Saboia, G.; Jorge, N.C.; Hoffmann, C.; Isaias, R.M.D.S.; Soares, G.L.; Zini, C.A. Development of a HS-SPME-GC/MS protocol assisted by chemometric tools to study herbivore-induced volatiles in Myrcia splendens. Talanta 2017, 175, 9–20. [Google Scholar] [CrossRef]
- Tajuddin, S.N.; Muhamad, N.S.; Yarmo, M.A.; Yusoff, M. Characterization of the Chemical Constituents of Agarwood Oils from Malaysia by Comprehensive Two-Dimensional Gas Chromatography–Time-of-Flight Mass Spectrometry. Mendeleev Commun. 2013, 23, 51–52. [Google Scholar] [CrossRef]
- Kvitvang, H.F.N.; Andreassen, T.; Adam, T.; Villas-Bôas, S.G.; Bruheim, P. Highly Sensitive GC/MS/MS Method for Quantitation of Amino and Nonamino Organic Acids. Anal. Chem. 2011, 83, 2705–2711. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Cao, K.-A.L.; Boitard, S.; Besse, P. Sparse PLS discriminant analysis: Biologically relevant feature selection and graphical displays for multiclass problems. BMC Bioinform. 2011, 12, 253. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S. Statistical evaluation and integration of multi-omics data with MetaboAnalyst. In Processing Metabolomics and Proteomics Data with Open Software: A Practical Guide; Royal Society of Chemistry: Cambridge, UK, 2020; pp. 281–301. [Google Scholar] [CrossRef]
- Li, S.; Zhong, X.; Kan, X.; Gu, L.; Sun, H.; Zhang, G.; Liu, X. De novo transcriptome analysis of Thitarodes jiachaensis before and after infection by the caterpillar fungus. Ophiocordyceps Sin. Gene 2016, 580, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Cebi, N.; Yilmaz, M.T.; Sagdic, O. A rapid ATR-FTIR spectroscopic method for detection of sibutramine adulteration in tea and coffee based on hierarchical cluster and principal component analyses. Food Chem. 2017, 229, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wu, B.-P.; Gao, L.-X.; Liu, H.-R.; Zhang, B.; Sun, C.-D.; Chen, K.-S. Glycosidically bound volatiles as affected by ripening stages of Satsuma mandarin fruit. Food Chem. 2018, 240, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wang, C.-H.; Zhang, Y.; Feng, Y.; Wang, Y.; Zhu, Y. Sparse Partial-least-squares Discriminant Analysis for Different Geographical Origins of Salvia miltiorrhiza by 1 H-NMR-based Metabolomics. Phytochem. Anal. 2013, 25, 50–58. [Google Scholar] [CrossRef]
- Park, C.H.; Baskar, T.B.; Park, S.-Y.; Kim, S.-J.; Arasu, M.V.; Al-Dhabi, N.A.; Kim, J.K.; Park, S.U. Metabolic Profiling and Antioxidant Assay of Metabolites from Three Radish Cultivars (Raphanus sativus). Molecules 2016, 21, 157. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Zheng, X.; Long, H.; Rao, Z.; Cao, L.; Han, R. Gut Bacterial and Fungal Communities of the Wild and Laboratory-Reared Thitarodes Larvae, Host of the Chinese Medicinal Fungus Ophiocordyceps sinensis on Tibetan Plateau. Insects 2021, 12, 327. [Google Scholar] [CrossRef]
- Zhong, X.; Gu, L.; Li, S.-S.; Kan, X.-T.; Zhang, G.-R.; Liu, X. Transcriptome analysis of Ophiocordyceps sinensis before and after infection of Thitarodes larvae. Fungal Biol. 2016, 120, 819–826. [Google Scholar] [CrossRef]
- Wang, J.; Kan, L.; Nie, S.; Chen, H.; Cui, S.; Phillips, A.O.; Phillips, G.O.; Li, Y.; Xie, M. A comparison of chemical composition, bioactive components and antioxidant activity of natural and cultured Cordyceps sinensis. LWT—Food Sci. Technol. 2015, 63, 2–7. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, H.; Li, S.; Zhong, X.; Wang, H.; Liu, X. Comparative metabolic profiling of Ophiocordyceps sinensis and its cultured mycelia using GC–MS. Food Res. Int. 2020, 134, 109241. [Google Scholar] [CrossRef]
- Lu, Y.; Zhi, Y.; Miyakawa, T.; Tanokura, M. Metabolic profiling of natural and cultured Cordyceps by NMR spectroscopy. Sci. Rep. 2019, 9, 7735. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Guo, Q.; Zheng, Q.; Zhang, W. Metabolomic comparison between wild Ophiocordyceps sinensis and artificial cultured Cordyceps militaris. Biomed. Chromatogr. 2018, 32, e4279. [Google Scholar] [CrossRef]
- Yao, C.-L.; Qian, Z.-M.; Tian, W.-S.; Xu, X.-Q.; Yan, Y.; Shen, Y.; Lu, S.-M.; Li, W.-J.; Guo, D.-A. Profiling and identification of aqueous extract of Cordyceps sinensis by ultra-high performance liquid chromatography tandem quadrupole-orbitrap mass spectrometry. Chin. J. Nat. Med. 2019, 17, 631–640. [Google Scholar] [CrossRef]
- Li, M.; Meng, Q.; Zhang, H.; Shu, R.; Zhao, Y.; Wu, P.; Li, X.; Zhou, G.; Qin, Q.; Zhang, J. Changes in transcriptomic and metabolomic profiles of morphotypes of Ophiocordyceps sinensis within the hemocoel of its host larvae, Thitarodes xiaojinensis. BMC Genom. 2020, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.N.; Kim, J.; Lee, M.Y.; Park, D.K.; Hong, Y.-S.; Lee, C.H. Metabolomics Revealed Novel Isoflavones and Optimal Cultivation Time of Cordyceps militaris Fermentation. J. Agric. Food Chem. 2010, 58, 4258–4267. [Google Scholar] [CrossRef]
- Rao, Z.; Cao, L.; Wu, H.; Qiu, X.; Liu, G.; Han, R. Comparative Transcriptome Analysis of Thitarodes Armoricanus in Response to the Entomopathogenic Fungi Paecilomyces Hepiali and Ophiocordyceps Sinensis. Insects 2019, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Rao, Z.-C.; Cao, L.; De Clercq, P.; Han, R.-C. Infection of Ophiocordyceps sinensis Fungus Causes Dramatic Changes in the Microbiota of Its Thitarodes Host. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xie, J.; Wang, L.; Li, S. Advanced development in chemical analysis of Cordyceps. J. Pharm. Biomed. Anal. 2014, 87, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Lipscomb, C.E. Medical subject headings (MeSH). Bull. Med. Libr. Assoc. 2000, 88, 265. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC35238/ (accessed on 30 September 2020). [PubMed]
- Bai, K.; Jiang, L.; Zhu, S.; Feng, C.; Zhao, Y.; Zhang, L.; Wang, T. Dimethylglycine sodium salt protects against oxidative damage and mitochondrial dysfunction in the small intestines of mice. Int. J. Mol. Med. 2019, 43, 2199–2211. [Google Scholar] [CrossRef] [PubMed]
- Cupp, M.J.; Tracy, T.S. Dimethylglycine (N, N-Dimethylglycine). In Dietary Supplements; Humana Press: Totowa, NJ, USA, 2003; pp. 149–160. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, R.; Qiu, X.-H.; Cao, L.; Long, H.-L.; Han, R.-C. Stage- and Rearing-Dependent Metabolomics Profiling of Ophiocordyceps sinensis and Its Pipeline Products. Insects 2021, 12, 666. https://doi.org/10.3390/insects12080666
Tang R, Qiu X-H, Cao L, Long H-L, Han R-C. Stage- and Rearing-Dependent Metabolomics Profiling of Ophiocordyceps sinensis and Its Pipeline Products. Insects. 2021; 12(8):666. https://doi.org/10.3390/insects12080666
Chicago/Turabian StyleTang, Rui, Xue-Hong Qiu, Li Cao, Hai-Lin Long, and Ri-Chou Han. 2021. "Stage- and Rearing-Dependent Metabolomics Profiling of Ophiocordyceps sinensis and Its Pipeline Products" Insects 12, no. 8: 666. https://doi.org/10.3390/insects12080666
APA StyleTang, R., Qiu, X. -H., Cao, L., Long, H. -L., & Han, R. -C. (2021). Stage- and Rearing-Dependent Metabolomics Profiling of Ophiocordyceps sinensis and Its Pipeline Products. Insects, 12(8), 666. https://doi.org/10.3390/insects12080666