
Evaluation of Wild Flora Surrounding Blueberry Fields as Viable Hosts of Drosophila suzukii (Matsumura, 1931) (Diptera: Drosophilidae) in Georgia


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Insect Rearing
2.3. Laboratory Assays
2.4. Data Analysis
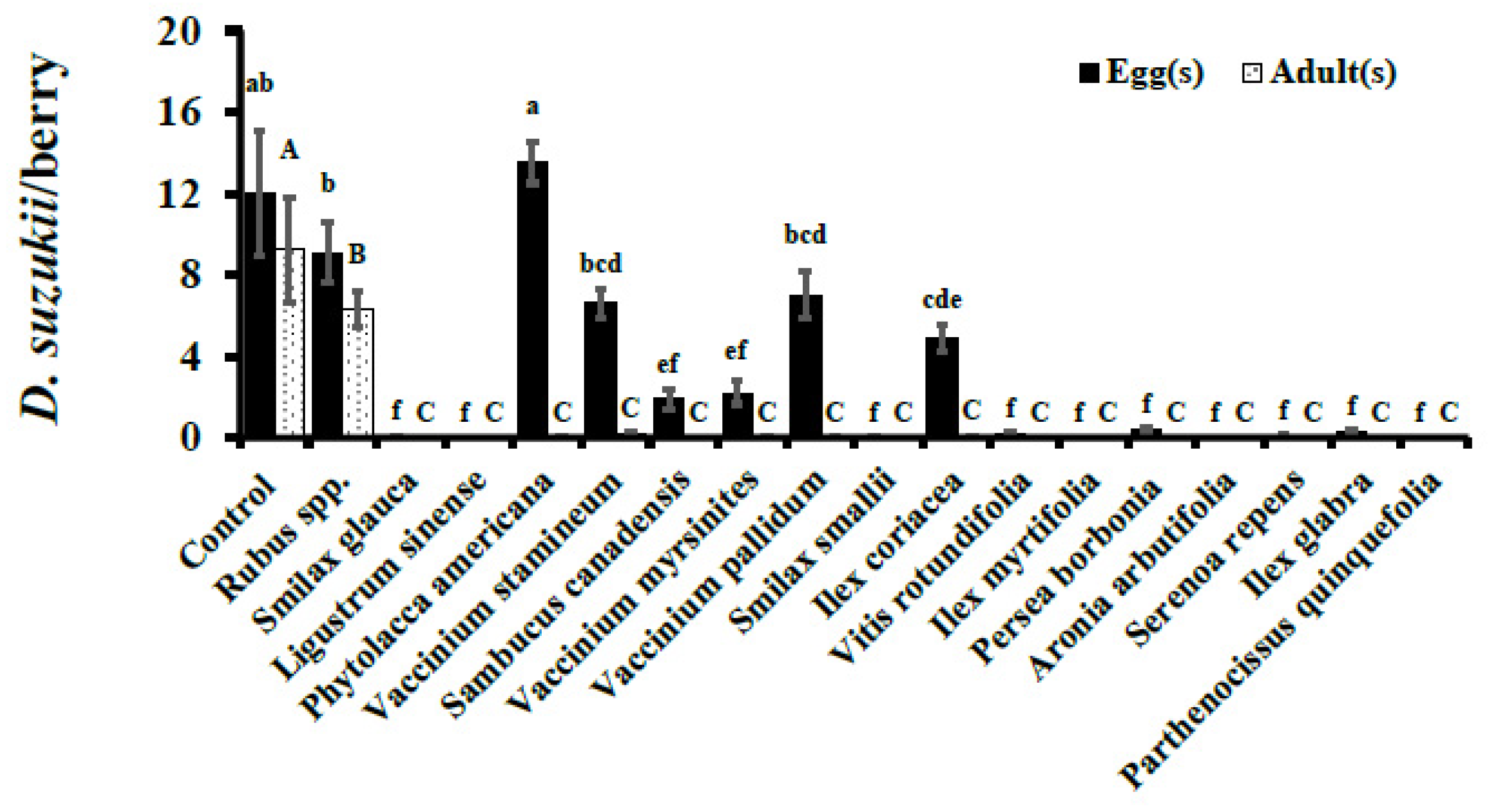
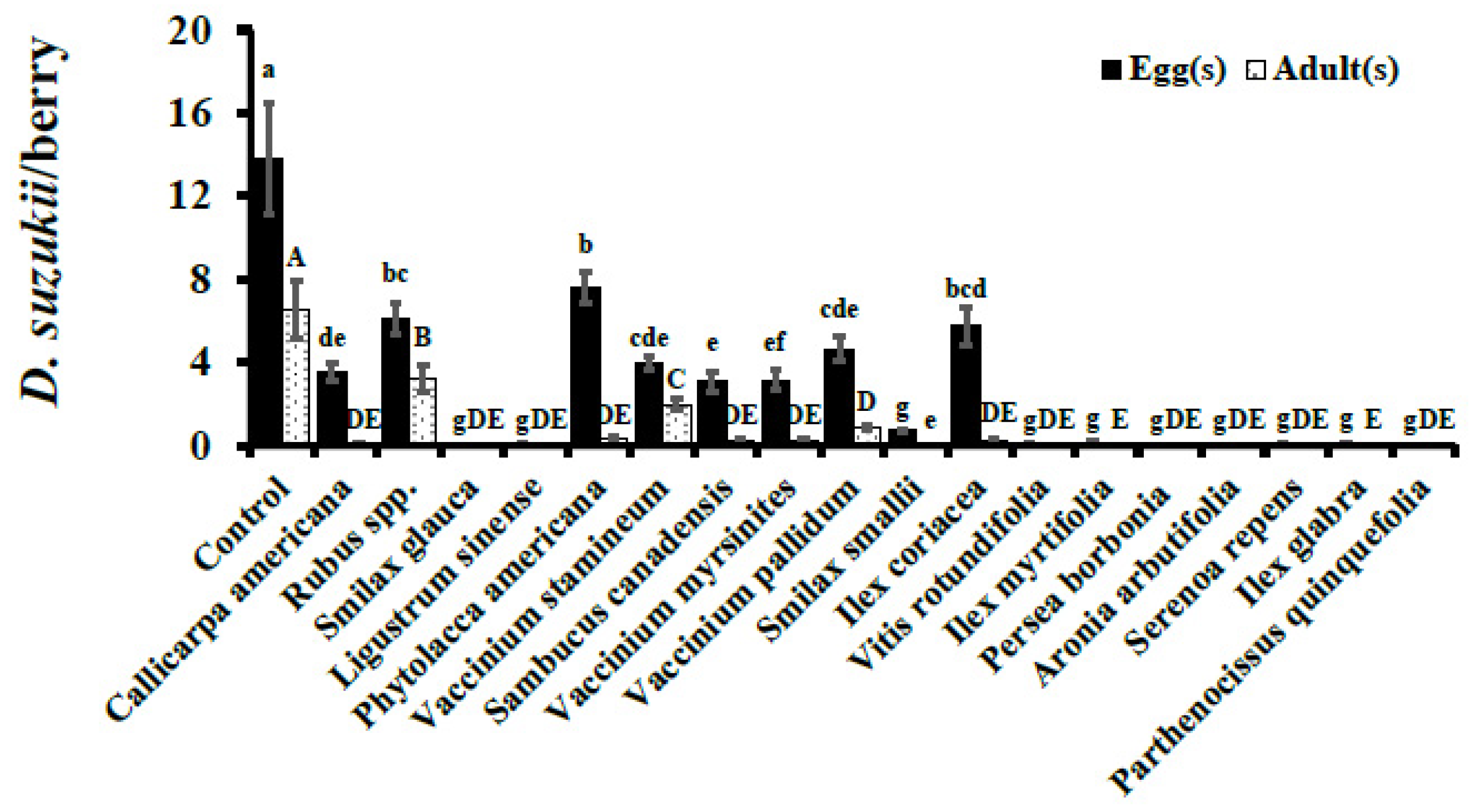
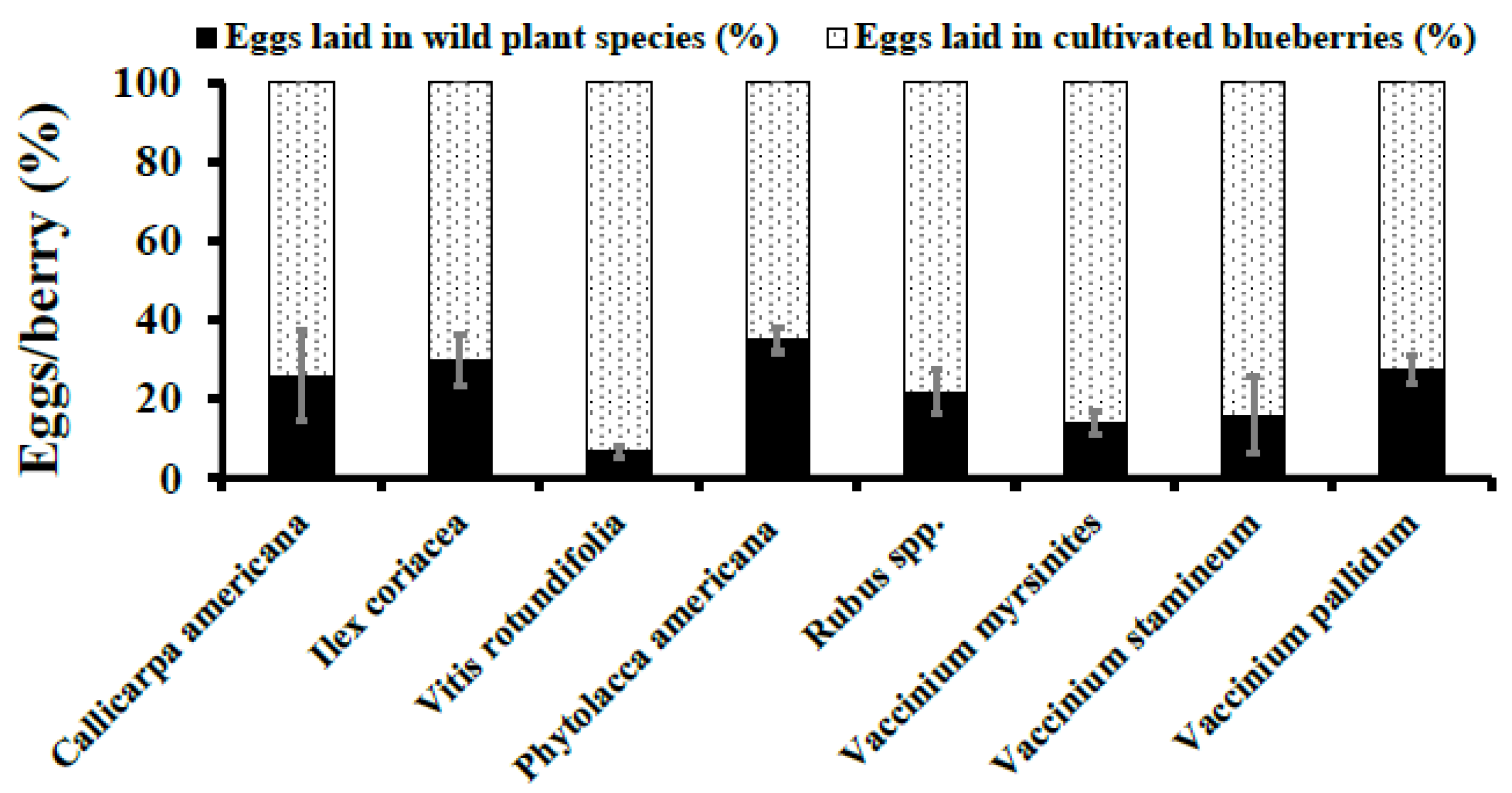
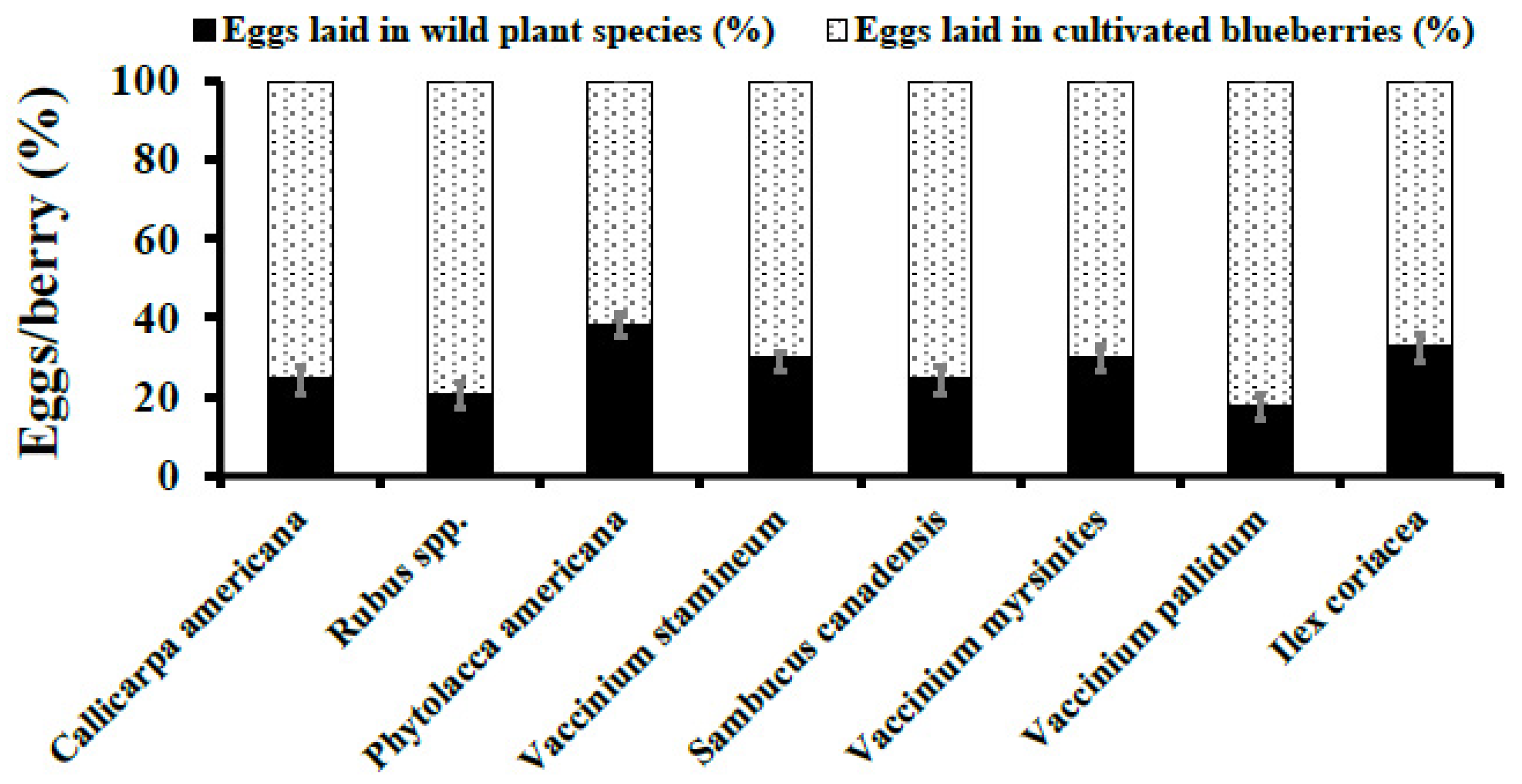
3. Results
3.1. No-Choice Assays
3.2. Choice Assays
3.3. Natural Infestation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zolam, F.G. Spotted-wing drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Steenwyk, R.A.V.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted-wing drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Burrack, H.J.; Smith, J.P.; Pfeiffer, D.G.; Koeher, G.; Laforest, J. Using volunteer-based networks to track Drosophila suzukii (Diptera: Drosophilidae) an invasive pest of fruit crops. J. Integr. Pest Manag. 2012, 3, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In focus: Spotted-wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, S.L.; Mehlferber, E.; Moore, P.J. Life-history trade-offs under different larval diets in Drosophila suzukii (Diptera: Drosophilidae). Physiol. Entomol. 2015, 40, 2–9. [Google Scholar] [CrossRef]
- Lee, J.C.; Dreves, A.J.; Cave, A.M.; Kawai, S.; Isaacs, R.; Miller, J.C.; Timmeren, S.V.; Bruck, D.J. Infestation of wild and ornamental noncrop fruits by Drosophila suzukii (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2015, 108, 117–129. [Google Scholar] [CrossRef]
- Mitsui, H.; Beppu, K.; Kimura, M.T. Seasonal life cycles and resource uses of flower- and fruit-feeding drosophilid flies (Diptera: Drosophilidae) in central Japan. Entomol. Sci. 2010, 13, 60–67. [Google Scholar] [CrossRef]
- Barata, A.; Santos, S.C.; Malfeito-Ferreira, M.; Loureiro, V. New insights into the ecological interaction between grape berry microorganisms and Drosophila flies during the development of sour rot. Microb. Ecol. 2012, 64, 416–430. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zalom, F.G.; Hamby, K.A. Host status and fruit odor response of Drosophila suzukii (Diptera: Drosophilidae) to figs and mulberries. J. Econ. Entomol. 2013, 106, 1932–1937. [Google Scholar] [CrossRef]
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The making of a pest: The evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proc. R. Soc. B 2014, 281, 2013–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Liburd, O.E.; Iglesias, L.E. Spotted-Wing Drosophila: Pest Management Recommendations for South-Eastern Blueberries; University of Florida, Institute of Food and Agricultural Services: Gainesville, FL, USA, 2013; Available online: https://www.sare.org/wp-content/uploads/Spotted-Wing-Drosophila.pdf (accessed on 16 November 2020).
- Beers, E.H.; Steenwyk, R.A.V.; Shearer, P.W.; Coates, W.W.; Grant, J.A. Developing Drosophila suzukii management programs for sweet cherry in the western United States. Pest Manag. Sci. 2011, 67, 1386–1395. [Google Scholar] [CrossRef]
- Iglesias, L.E.; Nyoike, T.W.; Liburd, O.E. Effect of trap design, bait type, and age on captures of Drosophila suzukii (Diptera: Drosophilidae) in berry crops. J. Econ. Entomol. 2014, 107, 1508–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diepenbrock, L.M.; Olivieri Rosensteel, D.; Harden, J.A.; Sial, A.A.; Burrack, H.J. Season-long programs for control of Drosophila suzukii in southeastern U.S. blueberries. Crop Prot. 2016, 81, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.; Stubbs, K. Georgia Farm Gate Value Report 2018; UGA Center for Agribusiness and Economic Development: Athens, GA, USA, 2019; Available online: https://caed.uga.edu/content/dam/caes-subsite/caed/publications/annual-reports-farm-gate-value-reports/2018%20Farm%20Gate.pdf (accessed on 16 November 2020).
- Lee, J.C.; Dalton, D.T.; Swoboda-Bhattari, K.A.; Bruck, D.J.; Burrack, H.J.; Strik, B.C.; Woltz, J.M.; Walton, V.M. Characterization and manipulation of fruit susceptibility to Drosophila suzukii. J. Pest Sci. 2016, 89, 771–780. [Google Scholar] [CrossRef] [Green Version]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying host potentials: Indexing postharvest fresh fruits for spotted-wing drosophila, Drosophila suzukii. PLoS ONE 2013, 4, e61227. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Song, Y.; Yi, C.; Zhang, Q.; Xia, H.; Lin, J.; Zhang, H.; Yang, J.; Ji, Q.; Chen, J. Potential host fruits for Drosophila suzukii: Olfactory and oviposition preferences and suitability for development. Entomol. Exp. Appl. 2019, 167, 880–890. [Google Scholar] [CrossRef]
- Conner, P.J. Performance of muscadine grape cultivars in southern Georgia. J. Am. Pomol. Soc. 2009, 63, 101–107. [Google Scholar]
- Harris, D.W.; Hamby, K.A.; Wilson, H.E.; Zalom, F.G. Seasonal monitoring of Drosophila suzukii (Diptera: Drosophilidae) in a mixed fruit production system. J. Asia-Pac. Entomol. 2014, 17, 857–864. [Google Scholar] [CrossRef]
- Grant, J.A.; Sial, A.A. Potential of muscadine grapes as a viable host of Drosophila suzukii (Diptera: Drosophilidae) in blueberry-producing regions of the southeastern United States. J. Econ. Entomol. 2016, 109, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.A. Investigation of Wild Flora Bordering Blueberry Fields to Quantify Their Potential as Viable Hosts of Drosophila suzukii (Diptera: Drosophilidae). Master’s Thesis, University of Georgia, Athens, Greece, 2016. [Google Scholar]
- Tonina, L.; Mori, N.; Sancassani, M.; Dall’Ara, P.; Marini, L. Spillover of Drosophila suzukii between noncrop and crop areas: Implications for pest management. Agric. For. Entomol. 2018, 20, 575–581. [Google Scholar] [CrossRef]
- Weissinger, L.; Schrieber, K.; Breuer, M.; Muller, C. Influences of blackberry margins on population dynamics of Drosophila suzukii and grape infestation in adjacent vineyards. J. Appl. Entomol. 2019, 143, 802–812. [Google Scholar] [CrossRef]
- Burrack, H.J.; Fernandez, G.E.; Spivey, T.; Kraus, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumura (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef]
- Steffan, S.A.; Lee, J.C.; Singleton, M.E.; Vilaire, A.; Walsh, D.B.; Lavine, L.S.; Patten, K. Susceptibility of cranberries to Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2013, 106, 2424–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poyet, M.; Eslin, P.; Heraude, M.; Roux, V.L.; Prevost, G.; Gilbert, P.; Chabrerie, O. Invasive host for invasive pest: When the Asiatic cherry fly (Drosophila suzukii) meets the American black cherry (Prunus serotina) in Europe. Agric. For. Entomol. 2014, 16, 251–259. [Google Scholar] [CrossRef]
- Poyet, M.; Roux, V.L.; Gilbert, P.; Meirlaand, A.; Prevost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the Asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briem, F.; Eben, A.; Gross, J.; Vogt, H. An invader supported by a parasite: Mistletoe berries as a host food and reproduction of spotted-wing drosophila in early spring. J. Pest Sci. 2016, 89, 749–759. [Google Scholar] [CrossRef]
- Leach, H.; Hagler, J.; Machtley, S.A.; Isaacs, R. Spotted-wing drosophila (Drosophila suzukii) utilization and dispersal from the wild host Asian bush honeysuckle (Lonicera spp.). Agric. For. Entomol. 2019, 21, 149–158. [Google Scholar] [CrossRef]
- Thistlewood, H.M.A.; Rozema, B.; Acheampong, S. Infestation and timing of use of non-crop plants by Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the Okanagan Basin, Canada. Can. Entomol. 2019, 151, 34–48. [Google Scholar] [CrossRef]
- Wang, X.; Kacar, G.; Daane, K.M. Temporal dynamics of host use by Drosophila suzukii in California’s San Joaquin Valley: Implications for area-wise pest management. Insects 2019, 10, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arno, J.; Sola, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Haviland, D.R.; Caprile, J.L.; Rill, S.M.; Hamby, K.A.; Grant, J.A. Phenology of spotted-wing drosophila in the San Joaquin Valley varies by season, crop and nearby vegetation. Calif. Agric. 2016, 70, 24–31. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2016, 140, 37–46. [Google Scholar] [CrossRef]
- Kacar, G.; Wang, X.; Stewart, T.J.; Daane, K.M. Overwintering survival of Drosophila suzukii (Diptera: Drosophilidae) and the effect of food on adult survival in California’s San Joaquin Valley. Environ. Entomol. 2015, 45, 763–771. [Google Scholar] [CrossRef]
- SAS Institute. JMP; Version 11; SAS Institute: Cary, NC, USA, 2014. [Google Scholar]
- Diepenbrock, L.M.; Swoboda-Bhattarai, K.A.; Burrack, H.J. Ovipositional preference, fidelity, and fitness of Drosophila suzukii in a co-occurring crop and non-crop host system. J. Pest Sci. 2016, 89, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Saona, C.; Cloonan, K.R.; Sanchez-Pedraza, F.; Zhou, Y.; Giusti, M.M.; Benrey, B. Differential susceptibility of wild and cultivated blueberries to an invasive frugivorous pest. J. Chem. Ecol. 2019, 45, 286–297. [Google Scholar] [CrossRef]
- Pelton, E.; Gratton, C.; Isaacs, R.; Timmeren, S.V.; Blanton, A.; Guedot, C. Earlier activity of Drosophila suzukii in high woodland landscapes but relative abundance is unaffected. J. Pest Sci. 2016, 89, 725–733. [Google Scholar] [CrossRef]
- Kinjo, H.; Kunimi, Y.; Nakai, M. Effects of temperature on the reproduction and development of Drosophila suzukii (Diptera: Drosophilidae). Appl. Entomol. Zool. 2014, 49, 297–304. [Google Scholar] [CrossRef]
- Hamby, K.A.; Bellamy, D.E.; Chiu, J.C.; Lee, J.C.; Walton, V.M.; Wiman, N.G.; York, R.M.; Biondi, A. Biotic and abiotic factors impacting development, behavior, phenology, and reproductive biology of Drosophila suzukii. J. Pest Sci. 2016, 89, 605–619. [Google Scholar] [CrossRef]
- Plantamp, C.; Salort, K.; Gilbert, P.; Dumet, A.; Mialdea, G.; Mondy, N.; Voituron, Y. All or nothing: Survival, reproduction and oxidative balance in spotted-wing drosophila (Drosophila suzukii) in response to cold. J. Insect Phys. 2016, 89, 28–36. [Google Scholar] [CrossRef]
- Ryan, G.D.; Emiljanowicz, L.; Wilkinson, F.; Kornya, M.; Newman, J.A. Thermal Tolerances of the spotted-wing drosophila Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 746–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, P.W.; West, J.D.; Walton, V.M.; Brown, P.H.; Svetec, N.; Chiu, J.C. Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecol. 2016, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Stockton, D.G.; Brown, R.; Loeb, G.M. Not berry hungry? Discovering the hidden food sources of a small fruit specialist, Drosophila suzukii. Ecol. Entomol. 2019, 44, 810–822. [Google Scholar] [CrossRef]
- Tochen, S.; Walton, V.M.; Lee, J.C. Impact of floral feeding on adult Drosophila suzukii survival and nutrient status. J. Pest Sci. 2016, 89, 793–802. [Google Scholar] [CrossRef]
- Wiman, N.G.; Walton, V.M.; Dalton, D.T.; Anfora, G.; Burrack, H.J.; Chiu, J.C.; Daane, K.M.; Grassi, A.; Miller, B.; Tochen, S.; et al. Integrating temperature-dependent life table data into a matrix projection estimation. PLoS ONE 2014, 9, e106909. [Google Scholar] [CrossRef]
- Entling, W.; Anslinger, S.; Jarausch, B.; Michl, G.; Hoffmann, C. Berry skin resistance explains oviposition preferences of Drosophila suzukii at the level of grape cultivars and single berries. J. Pest Sci. 2019, 93, 477–484. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; McGhee, P.S.; Hermann, S.L.; Gut, L.J.; Miller, J.R. Alightment of spotted-wing drosophila (Diptera: Drosophilidae) on odorless disks varying in color. Environ. Entomol. 2015, 45, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Little, C.M.; Dixon, P.L.; Chapman, T.W.; Hillier, N.K. Role of fruit characters and colour on host selection of boreal fruits and berries by Drosophila suzukii (Diptera: Drosophilidae). Can. Entomol. 2020, 152, 546–562. [Google Scholar] [CrossRef]
- Wohner, T.; Pinggera, J.; Fritzsche, E.; Peil, A.; Pinczinger, D.; Hanke, M. Insights into the susceptibility of raspberries to Drosophila suzukii oviposition. J. Appl. Entomol. 2021, 145, 182–190. [Google Scholar] [CrossRef]
- Letaief, H.; Rolle, L.; Zeppa, G.; Gerbi, V. Assessment of grape skin hardness by a puncture test. J. Sci. Food Agric. 2008, 88, 1567–1575. [Google Scholar] [CrossRef]
- Kinjo, H.; Kunimi, Y.; Ban, T.; Nakai, M. Oviposition efficacy of Drosophila suzukii (Diptera: Drosophilidae) on different cultivars of blueberry. J. Econ. Entomol. 2013, 106, 1767–1771. [Google Scholar] [CrossRef] [Green Version]
- Stewart, T.J.; Wang, X.G.; Molinar, A.; Danne, K.M. Factors limiting peach as potential host for Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2014, 107, 1771–1779. [Google Scholar] [CrossRef] [Green Version]
- Haye, T.; Girod, P.; Cuthbertsen, A.G.S.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef]
- Olazcuaga, L.; Rode, N.O.; Foucaud, J.; Facon, B.; Ravigne, V.; Ausset, A.; Lemenager, N.; Loiseau, A.; Gautier, M.; Estoup, A.; et al. Oviposition preference and larval performance of Drosophila suzukii (Diptera: Drosophilidae), spotted-wing drosophila: Effects of fruit identity and composition. Environ. Entomol. 2019, 48, 867–881. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, R.; Couty, A.; Eslin, P.; Gabola, F.; Chabrerie, O. The firethorn (Pyracantha coccinea), a promising dead-end trap plant for the biological control of the spotted-wing drosophila (Drosophila suzukii). Biol. Control 2020, 150, 104345. [Google Scholar] [CrossRef]
- Park, C.G.; Jang, M.; Yoon, K.A.; Kim, J. Insecticidal and acetylcholinesterase inhibitory activities of Lamiaceae plant essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Ind. Crops Prod. 2016, 89, 507–513. [Google Scholar] [CrossRef]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Tochen, S.; Dalton, D.T.; Wiman, N.G.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Tait, G.; Park, K.; Nieri, R.; Crava, M.C.; Mermer, S.; Clappa, E.; Boyer, G.; Dalton, D.T.; Carlin, S.; Brewer, L.; et al. Reproductive site selection: Evidence of an oviposition cue in a highly adaptive dipteran, Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2020, 49, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.G.; Stewart, T.J.; Bionidi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County | Latitude (N) | Longitude (W) |
|---|---|---|
| Appling | 31.75317 | −82.44336 |
| Bacon | 31.51020 | −82.45230 |
| Brantley | 31.18022 | −82.00486 |
| Clinch | 30.94030 | −82.68980 |
| Pierce | 31.45169 | −82.17831 |
| Ware | 31.15472 | −82.59180 |
| Species | 2015 | 2016 | ||
|---|---|---|---|---|
| n | Survivorship | n | Survivorship | |
| Callicarpa americana L. | 28 | 0.02 ± 0.01 b | ||
| Rubus spp. L. | 54 | 0.69 ± 0.14 ab | 46 | 0.72 ± 0.14 a |
| Smilax glauca Walt. | 2 | 0.0 ± 0.0 c | ||
| Ligustrum sinense Lour. | 1 | 0.0 ± 0.0 b | ||
| Phytolacca americana L. | 97 | 0.02 ± 0.01 c | 49 | 0.05 ± 0.02 b |
| Control | 8 | 0.82 ± 0.16 a | 10 | 0.57 ± 0.09 ab |
| Vaccinium stamineum L. | 69 | 0.04 ± 0.03 c | 50 | 0.66 ± 0.08 a |
| Sambucus Canadensis L. | 20 | 0.0 ± 0.0 c | 43 | 0.15 ± 0.06 b |
| Vaccinium myrsinites Lam. | 21 | 0.01 ± 0.01 c | 27 | 0.11 ± 0.05 b |
| Ilex glabra (L.) A. Gray | 6 | 0.0 ± 0.0 c | 4 | 0.0 ± 0.0 b |
| Vaccinium pallidum | 27 | 0.0 ± 0.0 c | 47 | 0.32 ± 0.06 b |
| Smilax smallii Morong. | 1 | 0.0 ± 0.0 c | 2 | 0.0 ± 0.0 b |
| Ilex coriacea (Purch) Chapm. | 46 | 0.01 ± 0.01 c | 46 | 0.09 ± 0.04 b |
| Vitis rotundifolia Michx. | 4 | 0.0 ± 0.0 c | 1 | 0.0 ± 0.0 b |
| Ilex myrtifolia Walt. | 5 | 0.0 ± 0.0 b | ||
| Persea borbonia (L.) Spreng. | 6 | 0.0 ± 0.0 c | ||
| Serenoa repens (W. Bartram) Small | 1 | 0.0 ± 0.0 c | 2 | 0.0 ± 0.0 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grant, J.A.; Sial, A.A. Evaluation of Wild Flora Surrounding Blueberry Fields as Viable Hosts of Drosophila suzukii (Matsumura, 1931) (Diptera: Drosophilidae) in Georgia. Insects 2021, 12, 667. https://doi.org/10.3390/insects12080667
Grant JA, Sial AA. Evaluation of Wild Flora Surrounding Blueberry Fields as Viable Hosts of Drosophila suzukii (Matsumura, 1931) (Diptera: Drosophilidae) in Georgia. Insects. 2021; 12(8):667. https://doi.org/10.3390/insects12080667
Chicago/Turabian StyleGrant, Joshua A., and Ashfaq A. Sial. 2021. "Evaluation of Wild Flora Surrounding Blueberry Fields as Viable Hosts of Drosophila suzukii (Matsumura, 1931) (Diptera: Drosophilidae) in Georgia" Insects 12, no. 8: 667. https://doi.org/10.3390/insects12080667
APA StyleGrant, J. A., & Sial, A. A. (2021). Evaluation of Wild Flora Surrounding Blueberry Fields as Viable Hosts of Drosophila suzukii (Matsumura, 1931) (Diptera: Drosophilidae) in Georgia. Insects, 12(8), 667. https://doi.org/10.3390/insects12080667