
Taxonomic Description of Stenodiplosis tectori n. sp. (Diptera: Cecidomyiidae), a Seed Parasite of Cheatgrass, Anisantha tectorum, Based on Morphological and Mitochondrial DNA Data


 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Collections of Specimens
2.2. Taxonomic Studies
2.3. Molecular and Phylogenetic Analyses
3. Results
3.1. Field Collections of Specimens
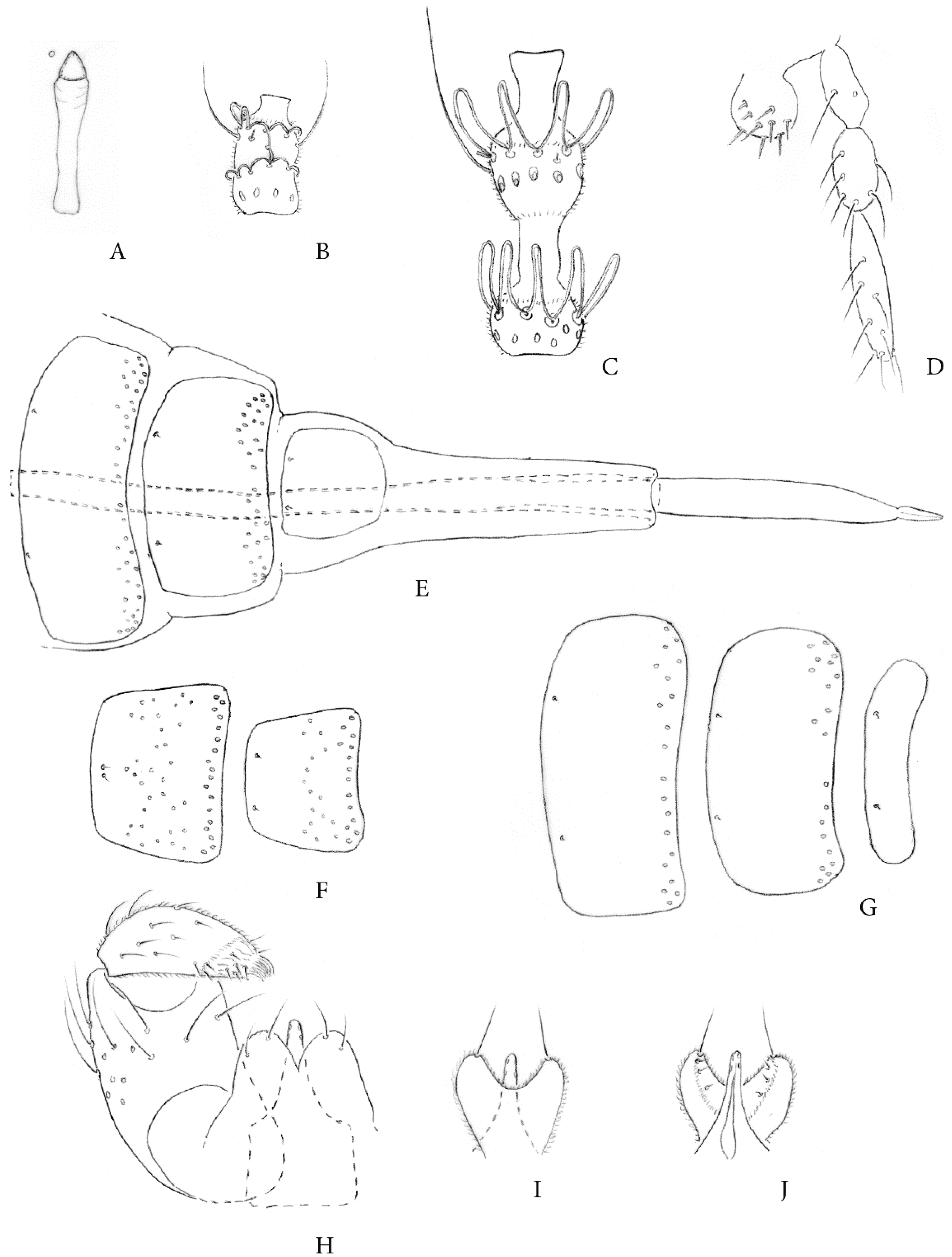
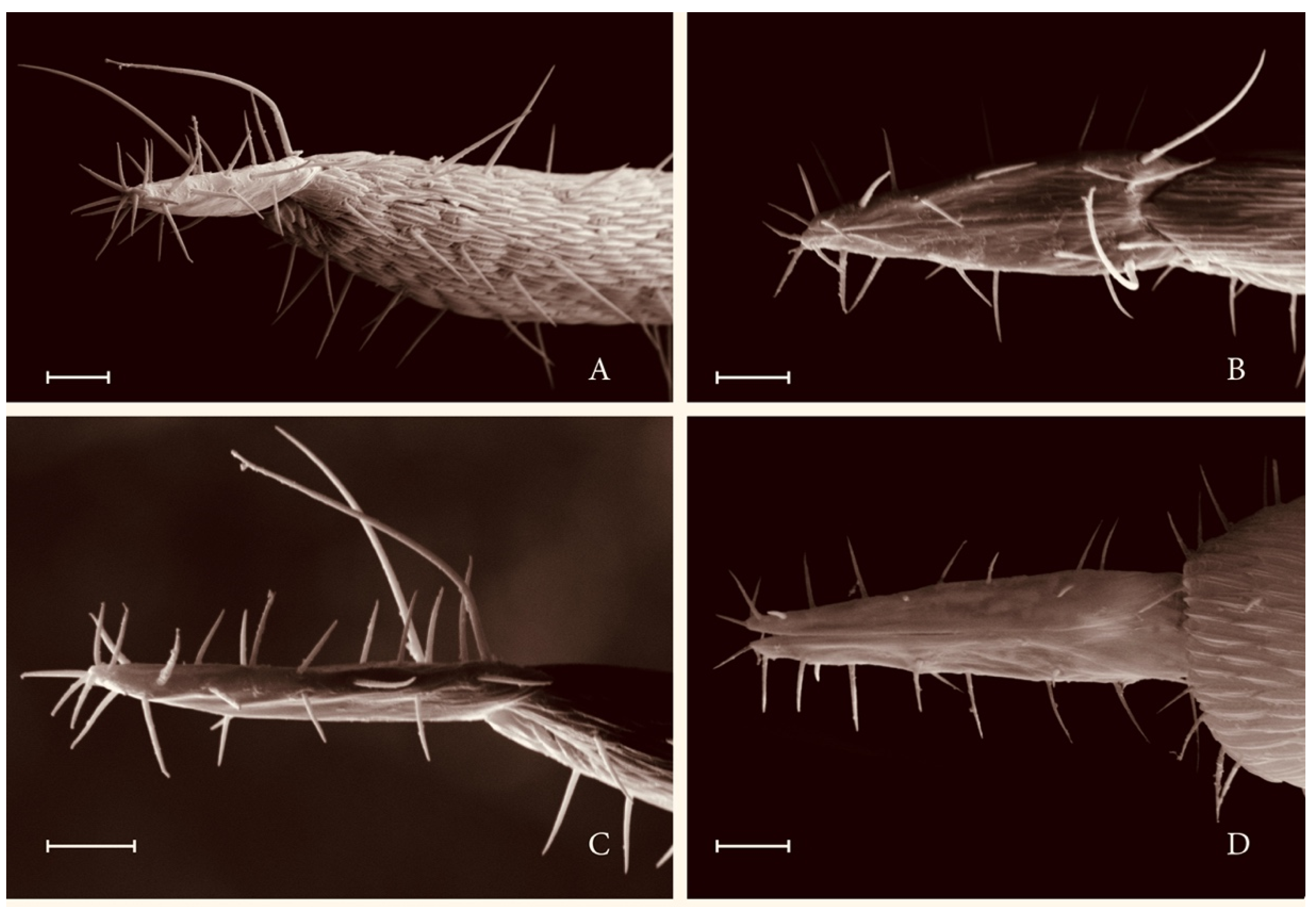
3.2. Taxonomic Description
3.2.1. Etymology
3.2.2. Notes on Differential Diagnosis
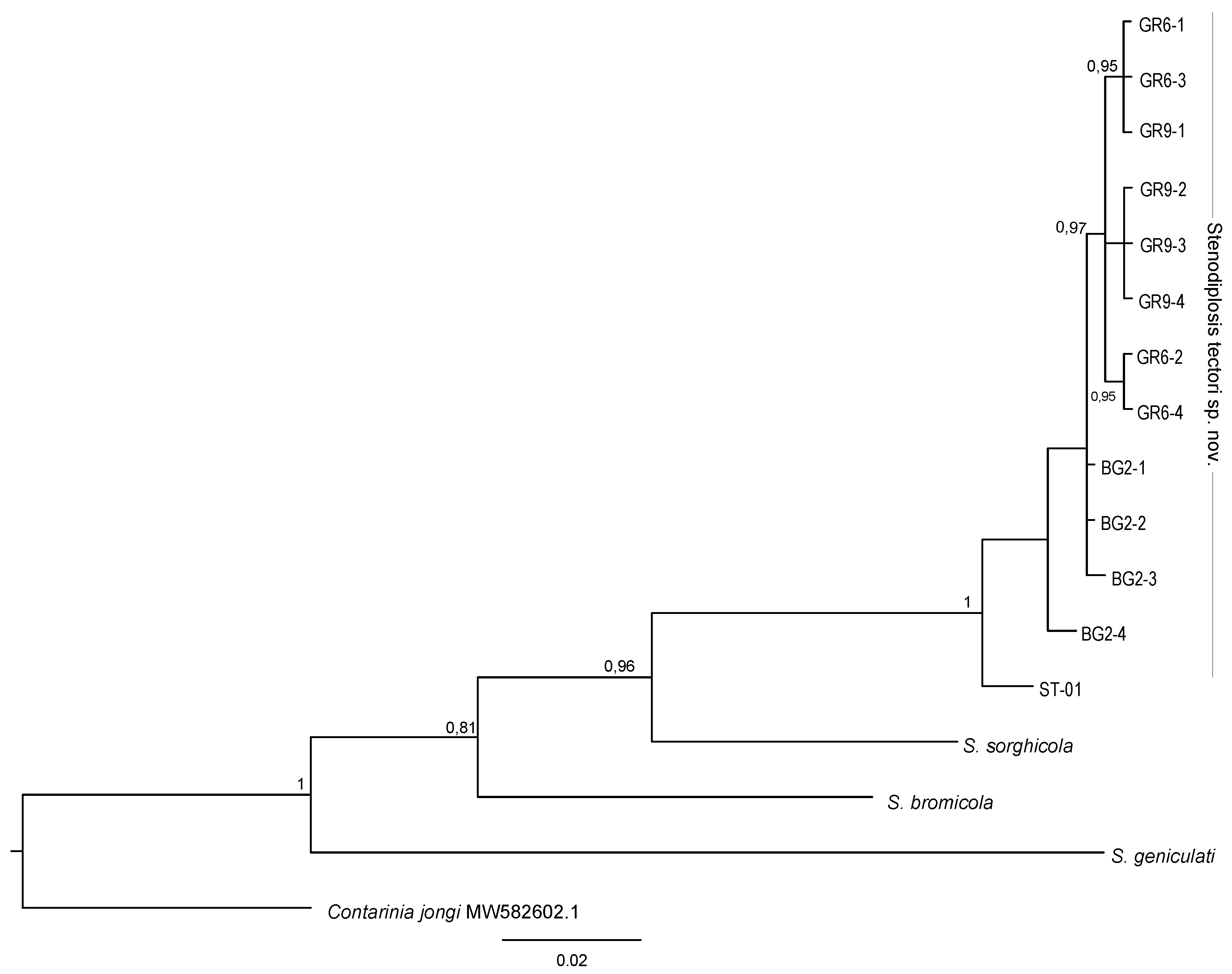
3.3. Molecular and Phylogenetic Analyses
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, J.A.; Evans, R.A. Downy brome—Intruder in the plant succession of big sagebrush communities in the Great Basin. J. Range Manag. 1973, 26, 410–415. [Google Scholar] [CrossRef]
- Mack, R.N. Invasion of Bromus tectorum L. into western North America: An ecological chronicle. Agro-Ecosyst. 1981, 7, 145–165. [Google Scholar] [CrossRef]
- Bradley, B.A.; Mustard, J.F. Identifying land cover variability distinct from land cover change: Cheatgrass in the Great Basin. Remote Sens. Environ. 2005, 94, 204–213. [Google Scholar] [CrossRef]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Balch, J.K.; Bradley, B.A.; D’Antonio, C.M.; Gómez-Dans, J. Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob. Chang. Biol. 2013, 19, 173–183. [Google Scholar] [CrossRef]
- Melgoza, G.; Nowak, R.S.; Tausch, R.J. Soil water exploitation after fire: Competition between Bromus tectorum (cheatgrass) and two native species. Oecologia 1990, 83, 7–13. [Google Scholar] [CrossRef]
- Rau, B.M.; Chambers, J.C.; Pyke, D.A.; Roundy, B.A.; Schupp, E.W.; Doescher, P.; Caldwell, T.G. Soil resources influence vegetation and response to fire and fire-surrogate treatments in sagebrush-steppe ecosystems. Rangel. Ecol. Manag. 2014, 67, 506–521. [Google Scholar] [CrossRef] [Green Version]
- Monty, A.; Brown, C.S.; Johnston, D.B. Fire promotes downy brome (Bromus tectorum L.) seed dispersal. Biol. Invasions 2013, 15, 1113–1123. [Google Scholar] [CrossRef]
- Baynes, M.; Newcombe, G.; Dixon, L.; Castlebury, L.; O’Donnell, K. A novel plant–fungal mutualism associated with fire. Fungal Biol. 2012, 116, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Stringham, T.K.; Krueger, W.C.; Shaver, P.L. State and transition modeling: An ecological process approach. J. Range Manag. 2003, 56, 106–113. [Google Scholar] [CrossRef]
- D’Antonio, C.M. Fire, plant invasions, and global changes. In Invasive Species in a Changing World; Mooney, H.A., Hobbs, R.J., Eds.; Island Press: Washington, DC, USA, 2000; pp. 65–93. [Google Scholar]
- DiTomaso, J.M.; Masters, R.A.; Peterson, V.F. Rangeland invasive plant management. Rangelands 2010, 32, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Horn, K.J.; St. Clair, S.B. Wildfire and exotic grass invasion alter plant productivity in response to climate variability in the Mojave Desert. Landsc. Ecol. 2017, 32, 635–646. [Google Scholar] [CrossRef]
- Hull, A.C., Jr.; Stewart, G. Replacing cheatgrass by reseeding with perennial grass on southern Idaho ranges. J. Am. Soc. Agron. 1948, 40, 694–703. [Google Scholar] [CrossRef] [Green Version]
- USDA-ARS. Development of Ecological Strategies for Invasive Plant Management and Rehabilitation of Western Rangelands. 2020. Available online: https://www.ars.usda.gov/research/project/?accnNo=439311 (accessed on 17 August 2021).
- Gagné, R.J.; Jaschhof, M.A. Catalog of the Cecidomyiidae (Diptera) of the World, 5th ed.; Digital; Systematic Entomology Laboratory, Agricultural Research Service, U.S. Department of Agriculture: Washington, DC, USA, 2021; 816p. Available online: https://www.ars.usda.gov/ARSUserFiles/80420580/Gagne_Jaschhof_2021_World_Cat_5th_Ed.pdf (accessed on 15 June 2021).
- Gagné, R.J. Key to adults of North American genera of the subfamily Cecidomyiinae (Diptera: Cecidomyiidae). Zootaxa 2018, 4392, 401–457. [Google Scholar] [CrossRef]
- Perilla Lopez, J.M.; Johnson, P.J.; Gagné, R.J.; Boe, A. A new species of Stenodiplosis (Diptera: Cecidomyiidae) on Spartina grasses (Poaceae) with notes on its biology and its parasitoid Tetrastichus bromi (Hymenoptera: Eulophidae). Zootaxa 2015, 4057, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Gagné, R.J. The Plant-Feeding Gall Midges of North America. Cornell University Press: Ithaca, NY, USA, 1989. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vriienhoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit 1 from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Sabbatini Peverieri, G.; Talamas, E.; Bon, M.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.; Hoelmer, K. Two Asian egg parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Möhn, E. Beiträge zur Systematik der Larven der Itonididae-(Cecidomyiidae, Diptera). Zoologica 1955, 105, 1–247. [Google Scholar]
- Agfonova, Z.Y. Development of the midge Stenodiplosis bromicola Mar. et Ag. (Diptera: Itonididae) in relation to peculiarities of brome biology (Bromus inermis Leyss. and riparius Rehm). Entomol. Obozr. 1962, 41, 11–21. [Google Scholar]
- Nieman, E.L.; Manglitz, G.R. The Biology and Ecology of the Bromegrass Seed Midge in Nebraska. Hist. Res. Bull. Neb. Agric. Exp. Stn. 1972, 102, 1913–1993. Available online: http://digitalcommons.unl.edu/ardhistrb/102 (accessed on 17 August 2021).




{kind=link}
{kind=link}
{kind=link}
| S. tectori Specimen ID/Stenodiplosis Species | Locality | Collector | Collection Date (dd/mm/yyyy) | Latitude, Longitude | MT-CO1 Fragment Length (bp) | S. tectori Haplotype | GenBank Accession Number |
|---|---|---|---|---|---|---|---|
| ST-01 | Maroneia, Greece | B.G.R. | 30/04/2018 | 40.87°, 25.53° | 615 | H1 | MZ382457 |
| BG2-1 | Hisarya, Bulgaria | M.C. | 01/06/2019 | 42.515°, 24.727° | 657 | H2 | MZ382458 |
| BG2-2 | Hisarya, Bulgaria | M.C. | 01/06/2019 | 42.515°, 24.727° | 657 | H2 | MZ382459 |
| BG2-3 | Hisarya, Bulgaria | M.C. | 01/06/2019 | 42.515°, 24.727° | 657 | H3 | MZ382460 |
| BG2-4 | Hisarya, Bulgaria | M.C. | 01/06/2019 | 42.515°, 24.727° | 657 | H4 | MZ382461 |
| GR6-1 | Maroneia, Greece | M.C. | 15/05/2019 | 40.87°, 25.53° | 657 | H5 | MZ382462 |
| GR6-2 | Maroneia, Greece | M.C. | 15/05/2019 | 40.87°, 25.53° | 657 | H6 | MZ382463 |
| GR6-3 | Maroneia, Greece | M.C. | 15/05/2019 | 40.87°, 25.53° | 657 | H5 | MZ382464 |
| GR6-4 | Maroneia, Greece | M.C. | 15/05/2019 | 40.87°, 25.53° | 657 | H6 | MZ382465 |
| GR9-1 | Kalabaka, Greece | M.C. | 16/05/2019 | 39.705°, 21.612° | 657 | H5 | MZ382466 |
| GR9-2 | Kalabaka, Greece | M.C. | 16/05/2019 | 39.705°, 21.612° | 657 | H7 | MZ382467 |
| GR9-3 | Kalabaka, Greece | M.C. | 16/05/2019 | 39.705°, 21.612° | 657 | H7 | MZ382468 |
| GR9-4 | Kalabaka, Greece | M.C. | 16/05/2019 | 39.705°, 21.612° | 657 | H7 | MZ382469 |
| S. bromicola ST-03 | Brookings, SD, USA | J.M.P.L. | 05/06/2015 | 44.5°, −96.53° | 615 | n/a | MZ382470 |
| S. sorghicola ST-04 | Brookings, SD, USA | J.M.P.L. | 05/06/2015 | 44.319°, −96.774° | 615 | n/a | MZ382471 |
| S. geniculati ST-02 | Brookings, SD, USA | J.M.P.L. | 05/06/2015 | 44.5°, −96.53° | 615 | n/a | MZ382472 |
| S. tectori (n = 13) | S. sorghicola (n = 1) | S. geniculati (n = 1) | S. bromicola (n = 1) | |
|---|---|---|---|---|
| S. tectori (n = 13) | 0.0067 ± 0.0017 | 0.0773 ± 0.011 | 0.129 ± 0.0145 | 0.0905 ± 0.0119 |
| S. sorghicola (n = 1) | 0.1295 ± 0.0149 | 0.0918 ± 0.0124 | ||
| S. geniculati (n = 1) | 0.1275 ± 0.0146 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rector, B.G.; Gagné, R.J.; Perilla López, J.M.; Tonkel, K.C.; Bon, M.-C.; Guermache, F.; Cristofaro, M. Taxonomic Description of Stenodiplosis tectori n. sp. (Diptera: Cecidomyiidae), a Seed Parasite of Cheatgrass, Anisantha tectorum, Based on Morphological and Mitochondrial DNA Data. Insects 2021, 12, 755. https://doi.org/10.3390/insects12080755
Rector BG, Gagné RJ, Perilla López JM, Tonkel KC, Bon M-C, Guermache F, Cristofaro M. Taxonomic Description of Stenodiplosis tectori n. sp. (Diptera: Cecidomyiidae), a Seed Parasite of Cheatgrass, Anisantha tectorum, Based on Morphological and Mitochondrial DNA Data. Insects. 2021; 12(8):755. https://doi.org/10.3390/insects12080755
Chicago/Turabian StyleRector, Brian G., Raymond J. Gagné, Juan Manuel Perilla López, Kirk C. Tonkel, Marie-Claude Bon, Fatiha Guermache, and Massimo Cristofaro. 2021. "Taxonomic Description of Stenodiplosis tectori n. sp. (Diptera: Cecidomyiidae), a Seed Parasite of Cheatgrass, Anisantha tectorum, Based on Morphological and Mitochondrial DNA Data" Insects 12, no. 8: 755. https://doi.org/10.3390/insects12080755
APA StyleRector, B. G., Gagné, R. J., Perilla López, J. M., Tonkel, K. C., Bon, M. -C., Guermache, F., & Cristofaro, M. (2021). Taxonomic Description of Stenodiplosis tectori n. sp. (Diptera: Cecidomyiidae), a Seed Parasite of Cheatgrass, Anisantha tectorum, Based on Morphological and Mitochondrial DNA Data. Insects, 12(8), 755. https://doi.org/10.3390/insects12080755