
Evaluation of Total Female and Male Aedes aegypti Proteomes Reveals Significant Predictive Protein–Protein Interactions, Functional Ontologies, and Differentially Abundant Proteins

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Protein Extraction
2.3. Sodium Dodecyl Sulphate–Polyacrylamide Gel Electrophoresis (SDS–PAGE)
2.4. In-Gel Protein Digestion
2.5. LC–ESI–MS/MS
2.6. Protein Identification Using Automated De Novo Sequencing (PEAKS Studio)
2.7. Prediction of Protein–Protein Interaction (PPI) Network and Functional Ontology Enrichment
2.8. Statistical Analysis for LFQ
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muktar, Y.; Tamerat, N.; Shewafera, A. Aedes aegypti as a vector of Flavivirus. J. Trop. Dis. 2016, 4, 2. [Google Scholar] [CrossRef]
- Whiten, S.R.; Keith Ray, W.; Helm, R.F.; Adelman, Z.N. Characterisation of the adult Aedes aegypti early midgut peritrophic matrix proteome using LC-MS. PLoS ONE 2018, 13, e0194734. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Fustec, B.; Cattel, J.; Chonephetsarath, S.; Thammavong, P.; Phommavanh, N.; David, J.-P.; Corbel, V.; Sutherland, L.W.; Hertz, J.C.; et al. Distribution of insecticide resistance and mechanisms involved in the arbovirus vector Aedes aegypti in Laos and implication for vector control. PLoS Negl. Trop. Dis. 2019, 13, e0007852. [Google Scholar] [CrossRef] [Green Version]
- Beserra, E.B.; Fernandes, C.R.M.; de, O.; Silva, S.A.; de Silva, L.A.; dos Santos, J.W. Efeitos da temperatura no ciclo de vida, exigências térmicas e estimativas do número de gerações anuais de Aedes aegypti (Diptera, Culicidae). Iheringia Série Zool. 2009, 99, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Hopp, M.J.; Foley, J.A. Global-scale relationships between climate and the dengue fever vector, Aedes aegypti. Clim. Chang. 2001, 48, 441–463. [Google Scholar] [CrossRef]
- Dhawan, R.; Kumar, M.; Mohanty, A.K.; Dey, G.; Advani, J.; Prasad, T.S.K.; Kumar, A. Mosquito-borne diseases and omics: Salivary gland proteome of the female Aedes aegypti mosquito. Omi. J. Integr. Biol. 2017, 21, 45–54. [Google Scholar] [CrossRef]
- Dhawan, R.; Mohanty, A.K.; Kumar, M.; Dey, G.; Advani, J.; Prasad, T.S.K.; Kumar, A. Data from salivary gland proteome analysis of female Aedes aegypti. Linn. Data Br. 2017, 13, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Springer Science & Business Media: Berlin, Germany, 2010. [Google Scholar]
- Cutwa, M.M.; O’Meara, G.F. Photographic Guide to Common Mosquitoes of Florida; Florida Medical Entomology Laboratory, University of Florida: Jacksonville, FL, USA, 2006. [Google Scholar]
- Valenzuela, J.G.; Francischetti, I.M.B.; Pham, V.M.; Garfield, M.K.; Ribeiro, J.M.C. Exploring the salivary gland transcriptome and proteome of the Anopheles stephensi mosquito. Insect Biochem. Mol. Biol. 2003, 33, 717–732. [Google Scholar] [CrossRef]
- Guerrero, D.; Cantaert, T.; Missé, D. Aedes mosquito salivary components and their effect on the immune response to arboviruses. Front. Cell. Infect. Microbiol. 2020, 10, 407. [Google Scholar] [CrossRef]
- Rawal, R.; Vijay, S.; Kadian, K.; Singh, J.; Pande, V.; Sharma, A. Towards a proteomic catalogue and differential annotation of salivary gland proteins in blood-fed malaria vector Anopheles culicifacies by mass spectrometry. PLoS ONE 2016, 11, e0161870. [Google Scholar] [CrossRef] [PubMed]
- De Lourdes Muñoz, M.; Limón-Camacho, G.; Tovar, R.; Diaz-Badillo, A.; Mendoza-Hernández, G.; Black, W.C. Proteomic identification of dengue virus binding proteins in Aedes aegypti mosquitoes and Aedes albopictus cells. Biomed. Res. Int. 2013, 2013, 875958. [Google Scholar]
- Mano, C.; Jariyapan, N.; Sor-Suwan, S.; Roytrakul, S.; Kittisenachai, S.; Tippawangkosol, P.; Somboon, P. Protein expression in female salivary glands of pyrethroid-susceptible and resistant strains of Aedes aegypti mosquitoes. Parasit. Vectors 2019, 12, 111. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.T.; Brito, N.F.; Oliveira, D.S.; Araujo, G.D.T.; Nogueira, F.C.S.; Domont, G.B.; Moreira, M.F.; Moreira, L.M.; Soares, M.R.; Melo, A.C.A. Comparative proteome analysis reveals that blood and sugar meals induce differential protein expression in Aedes aegypti female heads. Proteomics 2016, 16, 2582–2586. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Ramos, L.F.C.; Murillo, J.R.; Torres, A.; de Carvalho, S.S.; Domont, G.B.; de Oliveira, D.M.P.; Mesquita, R.D.; Nogueira, F.C.S.; Maciel-de-Freitas, R.; et al. Comprehensive quantitative proteome analysis of Aedes aegypti identifies proteins and pathways involved in Wolbachia pipientis and Zika virus interference phenomenon. BioRxiv 2020, 12, 199. [Google Scholar]
- Epelboin, Y.; Wang, L.; Giai Gianetto, Q.; Choumet, V.; Gaborit, P.; Issaly, J.; Guidez, A.; Douché, T.; Chaze, T.; Matondo, M.; et al. CYP450 core involvement in multiple resistance strains of Aedes aegypti from French Guiana highlighted by proteomics, molecular and biochemical studies. PLoS ONE 2021, 16, e0243992. [Google Scholar] [CrossRef] [PubMed]
- Rosilawati, R.; Nabila, R.; Siti Futri Farahininajua, F.; Nazni, W.A.; Lee, H.L. A preliminary proteomic study of permethrin resistant and susceptible Aedes aegypti (L.). Trop. Biomed. 2019, 36, 855–865. [Google Scholar]
- Wang, W.; Lv, Y.; Fang, F.; Hong, S.; Guo, Q.; Hu, S.; Zou, F.; Shi, L.; Lei, Z.; Ma, K.; et al. Identification of proteins associated with pyrethroid resistance by iTRAQ-based quantitative proteomic analysis in Culex pipiens pallens. Parasit. Vectors. 2015, 8, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl. Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Liu, H.; Zhang, L.; Liu, N. Resistance in the mosquito, Culex quinquefasciatus, and possible mechanisms for resistance. Pest. Manag. Sci. Former Pestic. Sci. 2005, 61, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Pando-Robles, V.; Batista, C.V. Aedes-Borne Virus–Mosquito Interactions: Mass Spectrometry Strategies and Findings. Vector-Borne Zoonotic Dis. 2017, 17, 361–375. [Google Scholar] [CrossRef]
- Tree, M.O.; Londono-Renteria, B.; Troupin, A.; Clark, K.M.; Colpitts, T.M.; Conway, M.J. Dengue virus reduces expression of low-density lipoprotein receptor-related protein 1 to facilitate replication in Aedes aegypti. Sci. Rep. 2019, 9, 6352. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Liu, P.; Mooney, B.P.; Franz, A.W.E. Quantitative proteomic analysis of chikungunya virus-infected aedes aegypti reveals proteome modulations indicative of persistent infection. J. Proteome Res. 2020, 19, 2443–2456. [Google Scholar] [CrossRef]
- Fernandes, K.M.; de Magalhães-Júnior, M.J.; Baracat-Pereira, M.C.; Martins, G.F. Proteomic analysis of Aedes aegypti midgut during post-embryonic development and of the female mosquitoes fed different diets. Parasitol. Int. 2016, 65. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.C.; Arcà, B.; Lombardo, F.; Calvo, E.; Phan, V.M.; Chandra, P.K.; Wikel, S.K. An annotated catalogue of salivary gland transcripts in the adult female mosquito, Aedes aegypti. BMC Genomics 2007, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chisenhall, D.M.; Christofferson, R.C.; McCracken, M.K.; Johnson, A.M.F.; Londono-Renteria, B.; Mores, C.N. Infection with dengue-2 virus alters proteins in naturally expectorated saliva of Aedes aegypti mosquitoes. Parasites Vectors 2014, 7, 252. [Google Scholar] [CrossRef] [Green Version]
- Almeras, L.; Fontaine, A.; Belghazi, M.; Bourdon, S.; Boucomont-Chapeaublanc, E.; Orlandi-Pradines, E.; Baragatti, M.; Corre-Catelin, N.; Reiter, P.; Pradines, B.; et al. Salivary gland protein repertoire from Aedes aegypti mosquitoes. Vector-Borne Zoonotic Dis. 2010, 10, 391–402. [Google Scholar] [CrossRef]
- Degner, E.C.; Ahmed-Braimah, Y.H.; Borziak, K.; Wolfner, M.F.; Harrington, L.C.; Dorus, S. Proteins, transcripts, and genetic architecture of seminal fluid and sperm in the mosquito Aedes aegypti. Mol. Cell. Proteomics. 2019, 18, S6–S22. [Google Scholar] [CrossRef]
- Camargo, C.; Ahmed-Braimah, Y.H.; Amaro, I.A.; Harrington, L.C.; Wolfner, M.F.; Avila, F.W. Mating and blood-feeding induce transcriptome changes in the spermathecae of the yellow fever mosquito. Aedes aegypti. Sci. Rep. 2020, 10, 14899. [Google Scholar] [CrossRef] [PubMed]
- Clemons, A.; Mori, A.; Haugen, M.; Severson, D.; Duman-Scheel, M. Aedes aegypti culturing and egg collection. Cold Spring Harb. Protoc. 2010, 2010, pdb–prot5507. [Google Scholar] [CrossRef]
- Kauffman, E.; Payne, A.; Franke, M.A.; Schmid, M.A.; Harris, E.; Kramer, L.D. Rearing of Culex spp. and Aedes spp. mosquitoes. Bio-Protocol 2017, 7, e2542. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Garver, L.; Dimopoulos, G. Protocol for mosquito rearing (An. gambiae). JoVE J. Vis. Exp. 2007, 5, e221. [Google Scholar] [CrossRef]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteomics 2012, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Shettima, A.; Ishak, I.H.; Rais, S.H.A.; Hasan, H.A.; Othman, N. Evaluation of female Aedes aegypti proteome via LC-ESI-MS/MS using two protein extraction methods. PeerJ 2021, 9, e10863. [Google Scholar] [CrossRef]
- Sun, P.; Nie, K.; Zhu, Y.; Liu, Y.; Wu, P.; Liu, Z.; Du, S.; Fan, H.; Chen, C.-H.; Zhang, R. A mosquito salivary protein promotes flavivirus transmission by activation of autophagy. Nat. Commun. 2020, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Wasinpiyamongkol, L.; Patramool, S.; Luplertlop, N.; Surasombatpattana, P.; Doucoure, S.; Mouchet, F.; Séveno, M.; Remoue, F.; Demettre, E.; Brizard, J.-P.; et al. Blood-feeding and immunogenic Aedes aegypti saliva proteins. Proteomics 2010, 10, 1906–1916. [Google Scholar] [CrossRef]
- Diakov, T.T.; Kane, P.M. Regulation of vacuolar proton-translocating ATPase activity and assembly by extracellular pH. J. Biol. Chem. 2010, 285, 23771–23778. [Google Scholar] [CrossRef] [Green Version]
- Sandiford, S.L.; Dong, Y.; Pike, A.; Blumberg, B.J.; Bahia, A.C.; Dimopoulos, G. Cytoplasmic actin is an extracellular insect immune factor which is secreted upon immune challenge and mediates phagocytosis and direct killing of bacteria, and is a Plasmodium Antagonist. PLoS Pathog. 2015, 11, e1004631. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Pridgeon, J.W.; Becnel, J.J.; Clark, G.G.; Linthicum, K.J. Mitochondrial gene cytochrome b developmental and environmental expression in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2009, 46, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Graifer, D.; Malygin, A.; Zharkov, D.O.; Karpova, G. Eukaryotic ribosomal protein S3: A constituent of translational machinery and an extraribosomal player in various cellular processes. Biochimie 2014, 99, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Champagne, D.E.; Smartt, C.T.; Ribeiro, J.M.C.; James, A.A. The salivary gland-specific apyrase of the mosquito Aedes aegypti is a member of the 5′-nucleotidase family. Proc. Natl. Acad. Sci. USA 1995, 92, 694–698. [Google Scholar] [CrossRef] [Green Version]
- Oktarianti, R.; Senjarini, K.; Hayano, T.; Fatchiyah, F.; Aulanni’am. Proteomic analysis of immunogenic proteins from salivary glands of Aedes aegypti. J. Infect Public Health 2015, 8, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Calvo, E.; Mans, B.J.; Andersen, J.F.; Ribeiro, J.M.C. Function and evolution of a mosquito salivary protein family. J. Biol. Chem. 2006, 281, 1935–1942. [Google Scholar] [CrossRef] [Green Version]
- Habibi, J.; Goodman, C.L.; Stuart, M.K. Distribution of elongation factor-1α in larval tissues of the fall armyworm, Spodoptera frugiperda. J. Insect Sci. 2006, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Nag, J.K.; Shrivastava, N.; Chahar, D.; Gupta, C.L.; Bajpai, P.; Misra-Bhattacharya, S. Wolbachia transcription elongation factor “Wol GreA” interacts with α2ββ′ σ subunits of RNA polymerase through its dimeric c-terminal domain. PLoS Negl. Trop. Dis. 2014, 8, e2930. [Google Scholar] [CrossRef] [Green Version]
- Salas-Benito, J.S.; Nova-Ocampo, D. Viral interference and persistence in mosquito-borne flaviviruses. J. Immunol. Res. 2015, 2015, 873404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colpitts, T.M.; Cox, J.; Nguyen, A.; Feitosa, F.; Krishnan, M.N.; Fikrig, E. Use of a tandem affinity purification assay to detect interactions between West Nile and dengue viral proteins and proteins of the mosquito vector. Virology 2011, 417, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zheng, X.; Wu, Y.; Gan, M.; He, A.; Li, Z.; Zhang, D.; Wu, X.; Zhan, X. Differential proteomics of Aedes albopictus salivary gland, midgut and C6/36 cell induced by dengue virus infection. Virology 2013, 444, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, A.S.; Feitosa-Suntheimer, F.; Araujo, R.V.; Hekman, R.M.; Asad, S.; Londono-Renteria, B.; Emili, A.; Colpitts, T.M. Dengue Virus Infection of Aedes aegypti Alters Extracellular Vesicle Protein Cargo to Enhance Virus Transmission. Int. J. Mol. Sci. 2020, 21, 6609. [Google Scholar] [CrossRef]
- Clayton, A.M.; Dong, Y.; Dimopoulos, G. The Anopheles innate immune system in the defense against malaria infection. J. Innate Immun. 2014, 6, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-mediated endocytosis. J. Gen. Virol. 2008, 89, 474–484. [Google Scholar] [CrossRef] [PubMed]







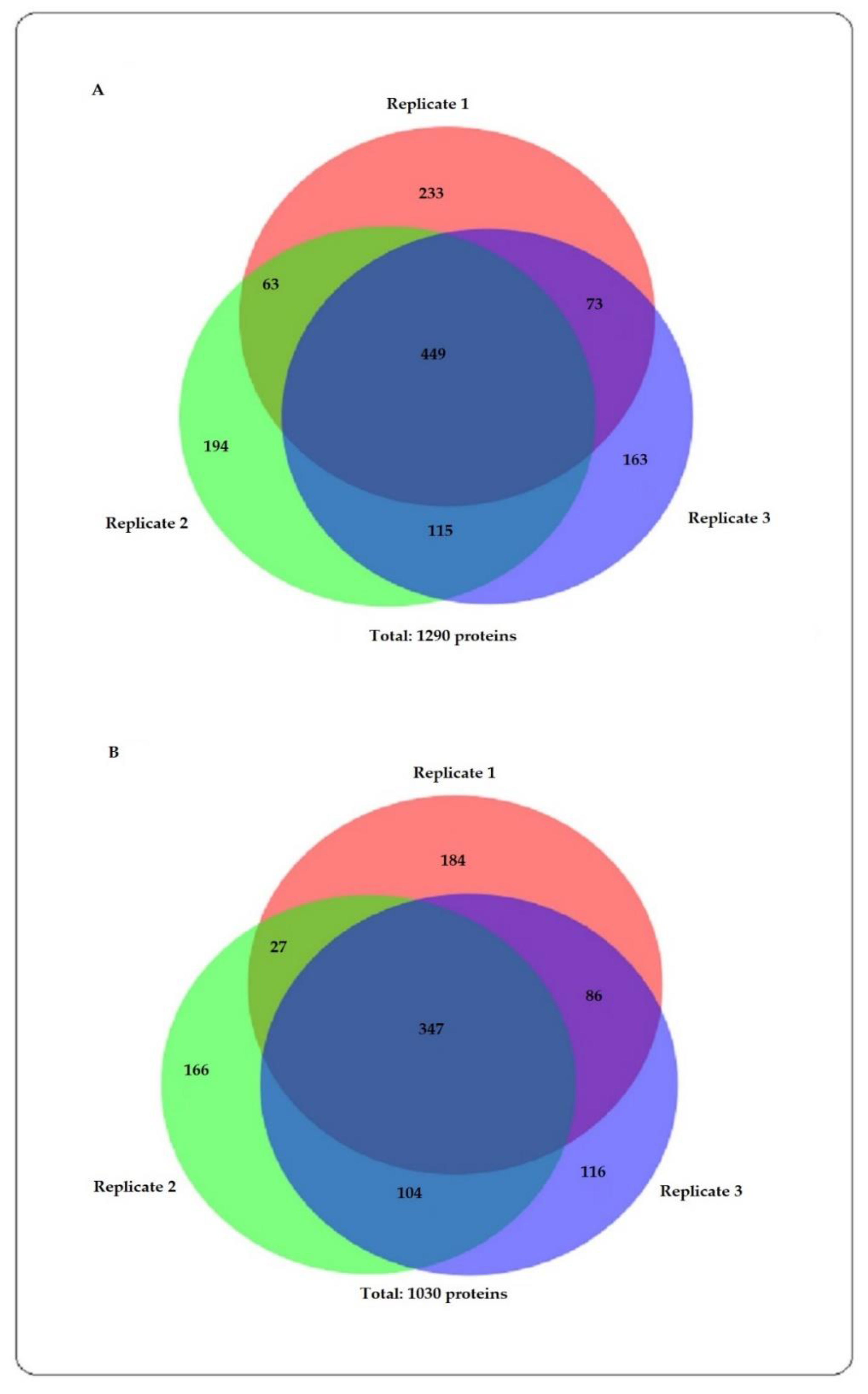{kind=link}
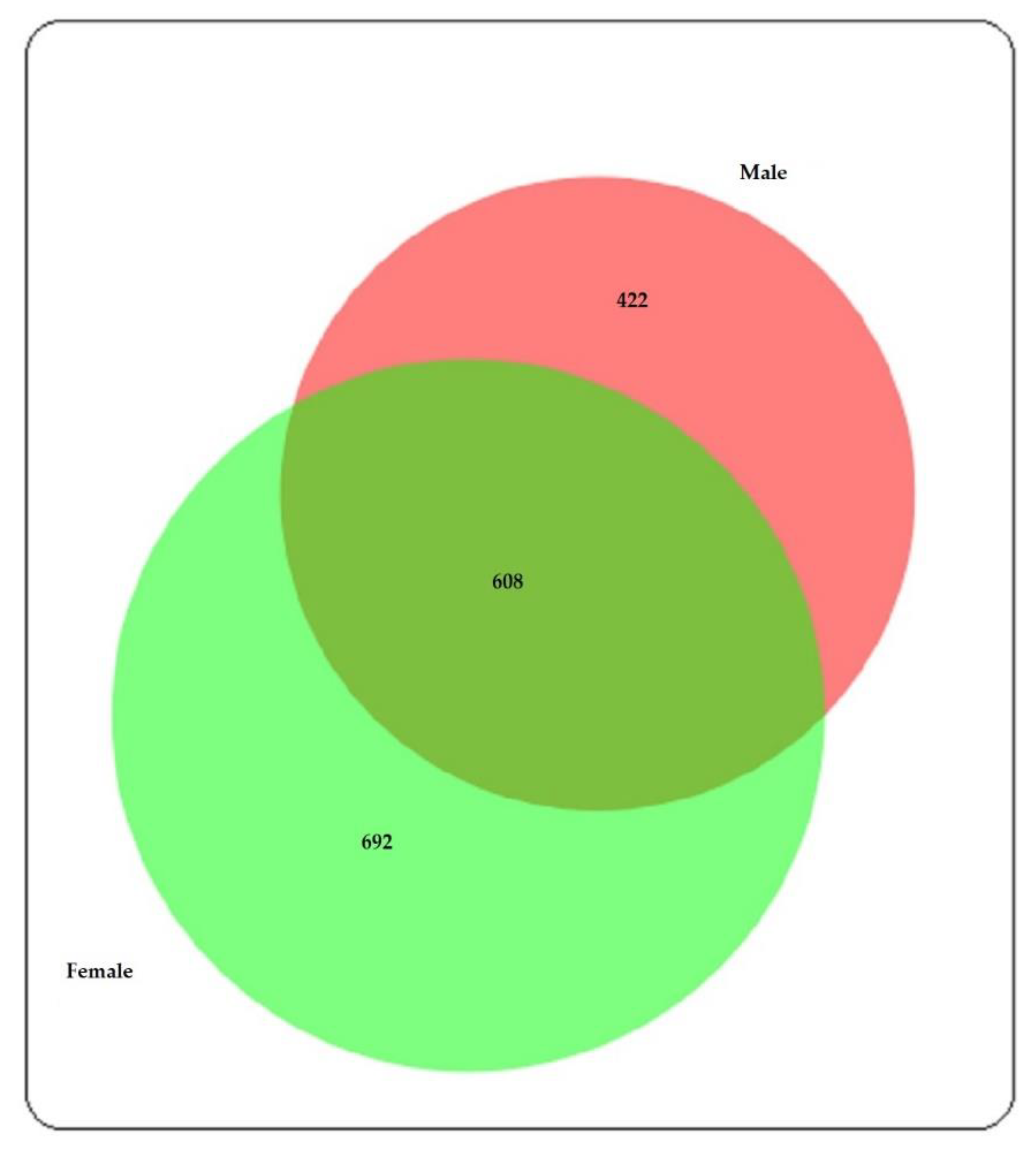{kind=link}
{kind=link}
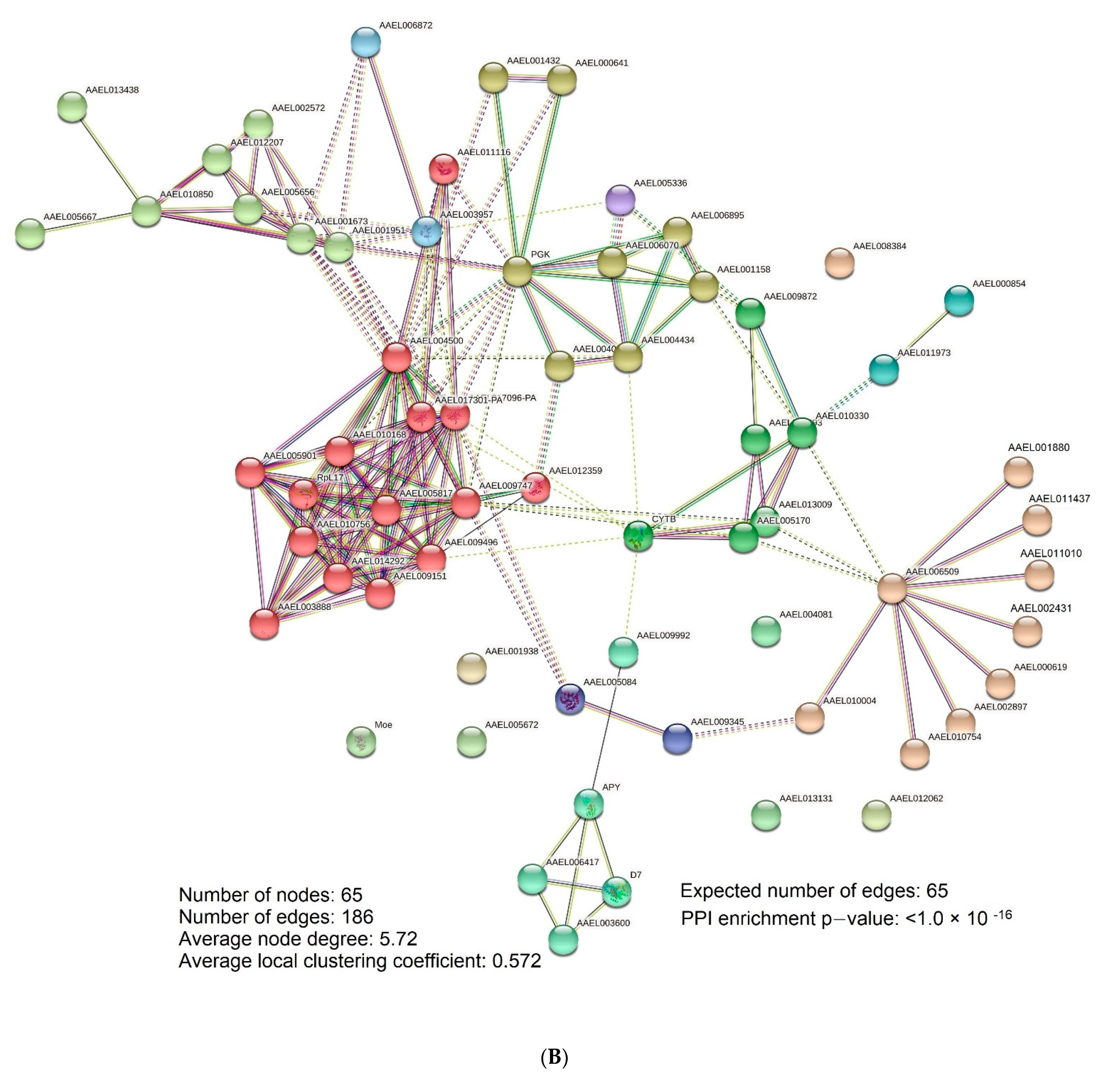{kind=link}
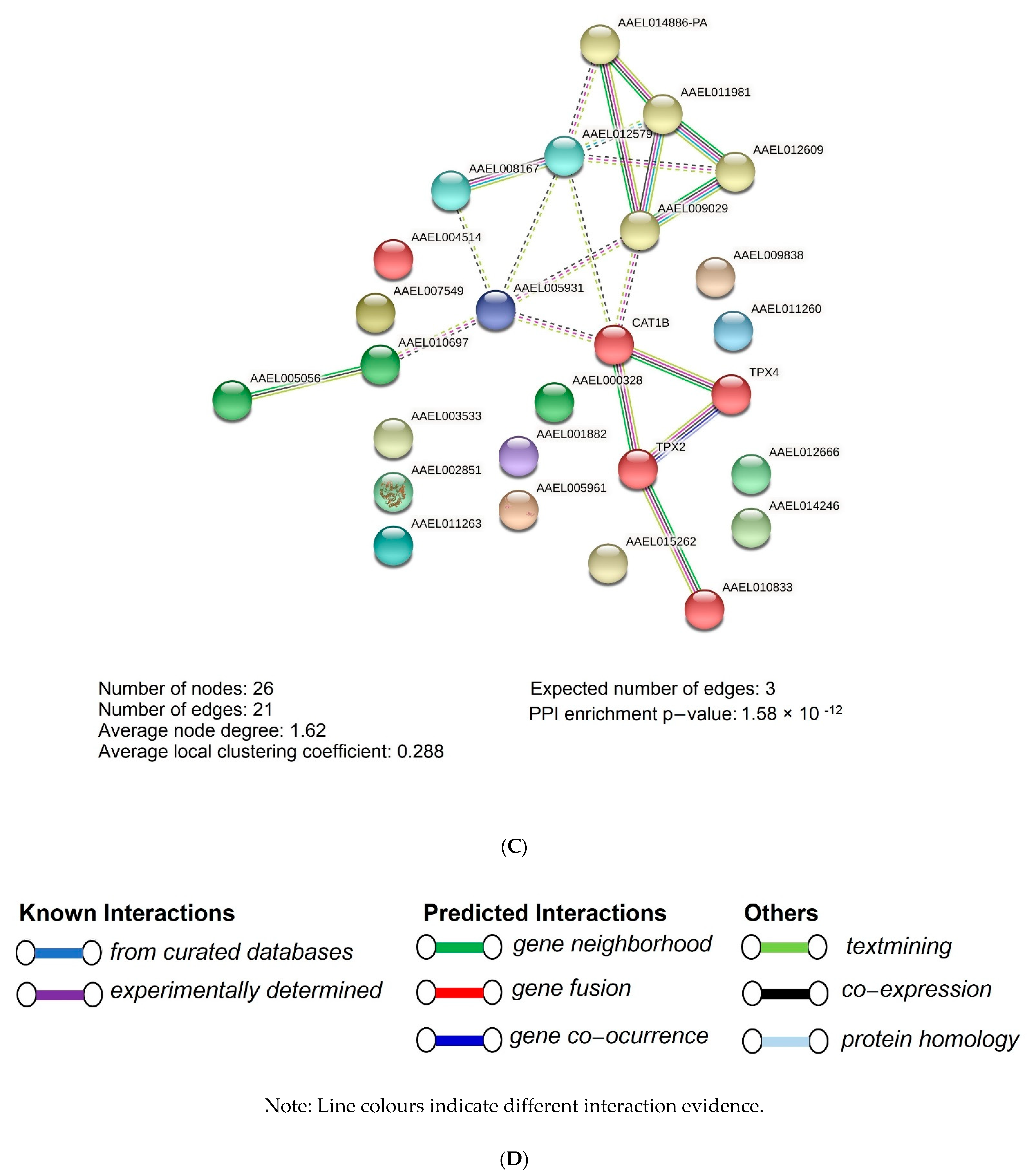{kind=link}
{kind=link}
| (A) Biological Process | ||||||
| Term ID | TermDescription | ObservedGene Count | BackgroundGene Count | Strength | FalseDiscovery Rate | MatchingProteinsin theNetwork (Labels) |
| GO:0005975 | Carbohydrate metabolic process | 2 | 5 | 1.83 | 0.0126 | AAEL002956AAEL011789 |
| GO:0006099 | Tricarboxylic acid cycle | 2 | 2 | 2.23 | 0.0126 | AAEL002956 AAEL011789 |
| GO:0006101 | Citrate metabolic process | 2 | 2 | 2.23 | 0.0126 | AAEL002956AAEL011789 |
| GO:0016999 | Antibiotic metabolic process | 2 | 3 | 2.05 | 0.0126 | AAEL002956AAEL011789 |
| GO:0017144 | Drug metabolic process | 3 | 21 | 1.38 | 0.0126 | AAEL002956 VhaA AAEL011789 |
| GO:0044281 | Small-molecule metabolic process | 3 | 31 | 1.21 | 0.0126 | AAEL002956 VhaA AAEL011789 |
| (B) Molecular Function | ||||||
| GO:0004108 | Citrate (Si)-synthase activity | 2 | 2 | 2.23 | 0.0101 | AAEL002956 AAEL011789 |
| (C) Cellular Component | ||||||
| GO:0005759 | Mitochondrial matrix | 2 | 4 | 1.92 | 0.0097 | AAEL002956 AAEL011789 |
| Term ID | Term Description | Observed Gene Count | Background Gene Count | Strength | False Discovery Rate | Matching Proteins in the Network (Labels) |
|---|---|---|---|---|---|---|
| GO:0005737 | Cytoplasm | 4 | 119 | 0.91 | 0.0211 | AAEL005901 Moe RpL17 CYTB |
| GO:0005840 | Ribosome | 2 | 14 | 1.54 | 0.0211 | AAEL005901 RpL17 |
| GO:0032991 | Protein-containing complex | 3 | 77 | 0.98 | 0.0211 | AAEL005901 RpL17 CYTB |
| GO:0043229 | Intracellular organelle | 4 | 135 | 0.86 | 0.0211 | AAEL005901 Moe RpL17 CYTB |
| GO:0043232 | Intracellular non-membrane-bounded organelle | 3 | 36 | 1.31 | 0.0211 | AAEL005901 Moe RpL17 |
| GO:1990904 | Ribonucleoprotein complex | 2 | 16 | 1.48 | 0.0211 | AAEL005901 RpL17 |
| GO:0005576 | Extracellular region | 2 | 24 | 1.31 | 0.0219 | APY D7 |
| GO:0005623 | Cell | 4 | 186 | 0.72 | 0.0271 | AAEL005901 Moe RpL17 CYTB |
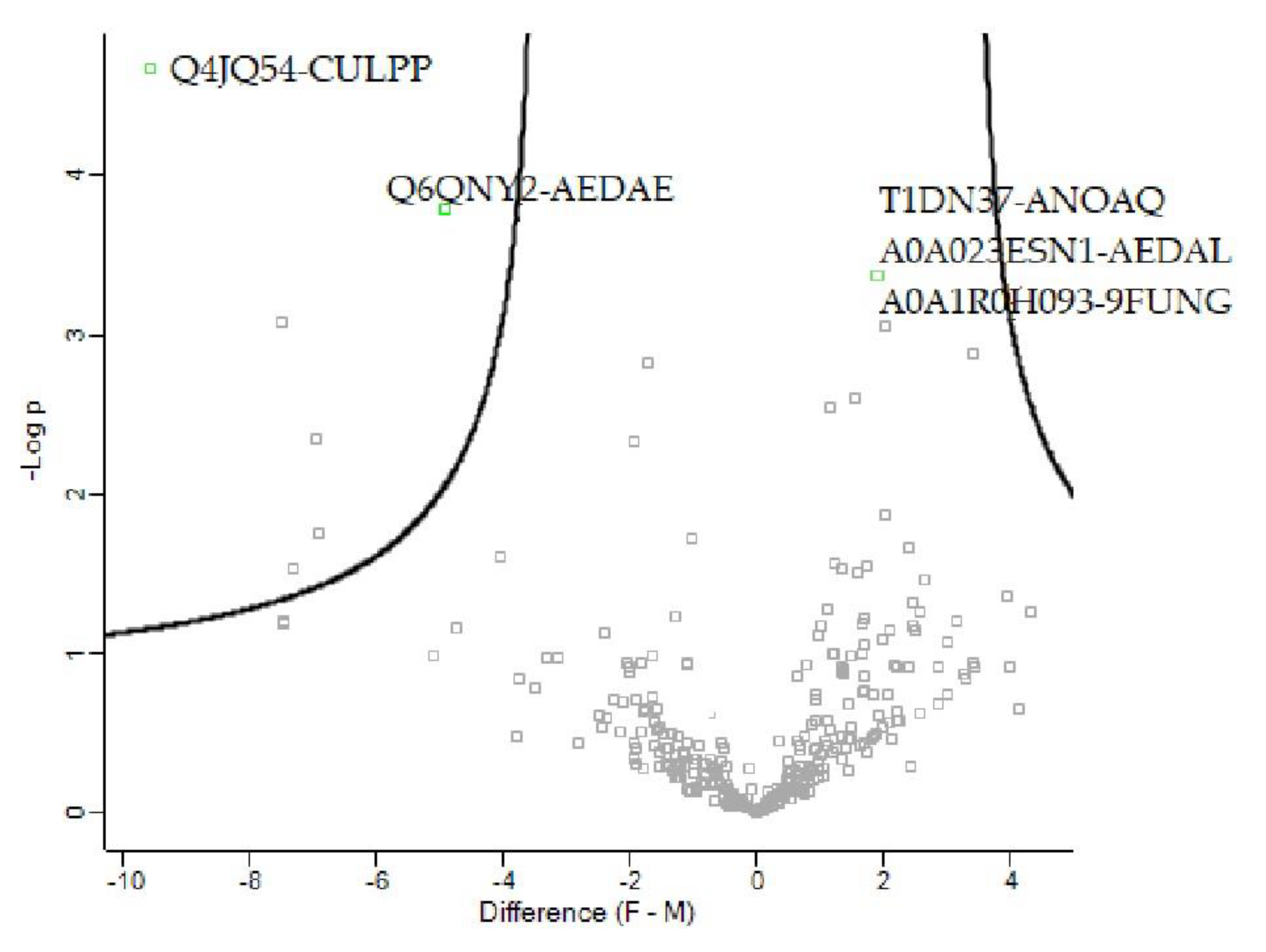
| Name | Accession Number | Fold Change (t-Test Difference) | q-Value | Abundance |
|---|---|---|---|---|
| Elongation factor 1α (fragment) | A0A023ESN1_AEDAL | 1.8433 | 0.017 | Increased |
| AAEL001951-PA | Q6QNY2_AEDAE | 4.91199 | 0.034 | Decreased |
| ACTIN | Q4JQ54_CULPP | 9.55168 | 0.00 | Decreased |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shettima, A.; Joseph, S.; Ishak, I.H.; Abdul Raiz, S.H.; Abu Hasan, H.; Othman, N. Evaluation of Total Female and Male Aedes aegypti Proteomes Reveals Significant Predictive Protein–Protein Interactions, Functional Ontologies, and Differentially Abundant Proteins. Insects 2021, 12, 752. https://doi.org/10.3390/insects12080752
Shettima A, Joseph S, Ishak IH, Abdul Raiz SH, Abu Hasan H, Othman N. Evaluation of Total Female and Male Aedes aegypti Proteomes Reveals Significant Predictive Protein–Protein Interactions, Functional Ontologies, and Differentially Abundant Proteins. Insects. 2021; 12(8):752. https://doi.org/10.3390/insects12080752
Chicago/Turabian StyleShettima, Abubakar, Shaleni Joseph, Intan H. Ishak, Syahirah Hanisah Abdul Raiz, Hadura Abu Hasan, and Nurulhasanah Othman. 2021. "Evaluation of Total Female and Male Aedes aegypti Proteomes Reveals Significant Predictive Protein–Protein Interactions, Functional Ontologies, and Differentially Abundant Proteins" Insects 12, no. 8: 752. https://doi.org/10.3390/insects12080752
APA StyleShettima, A., Joseph, S., Ishak, I. H., Abdul Raiz, S. H., Abu Hasan, H., & Othman, N. (2021). Evaluation of Total Female and Male Aedes aegypti Proteomes Reveals Significant Predictive Protein–Protein Interactions, Functional Ontologies, and Differentially Abundant Proteins. Insects, 12(8), 752. https://doi.org/10.3390/insects12080752