
A New Cixiid Fossil Genus of the Tribe Acrotiarini from Mid-Cretaceous Burmese Amber (Insecta, Hemiptera, Fulgoromorpha) †


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
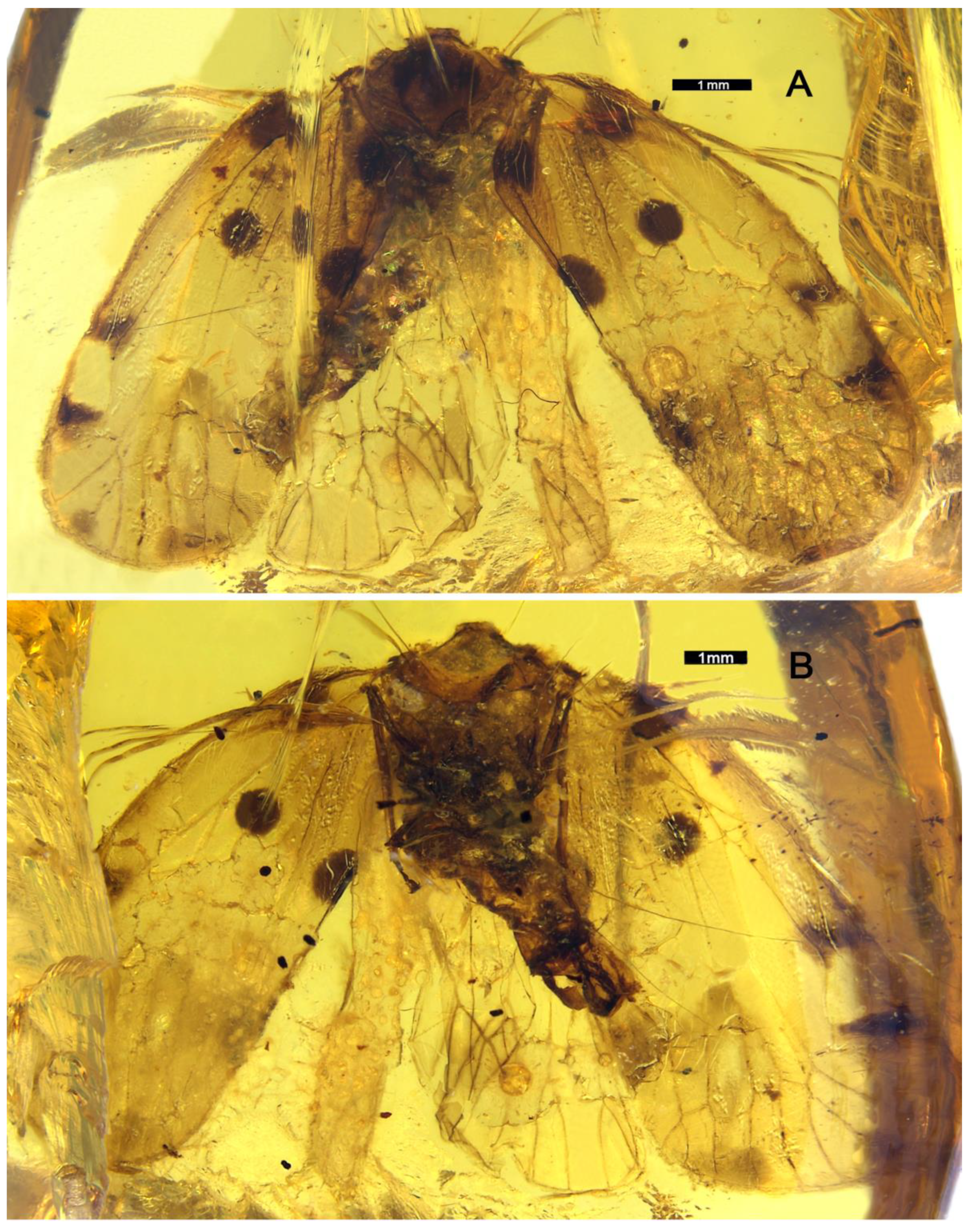
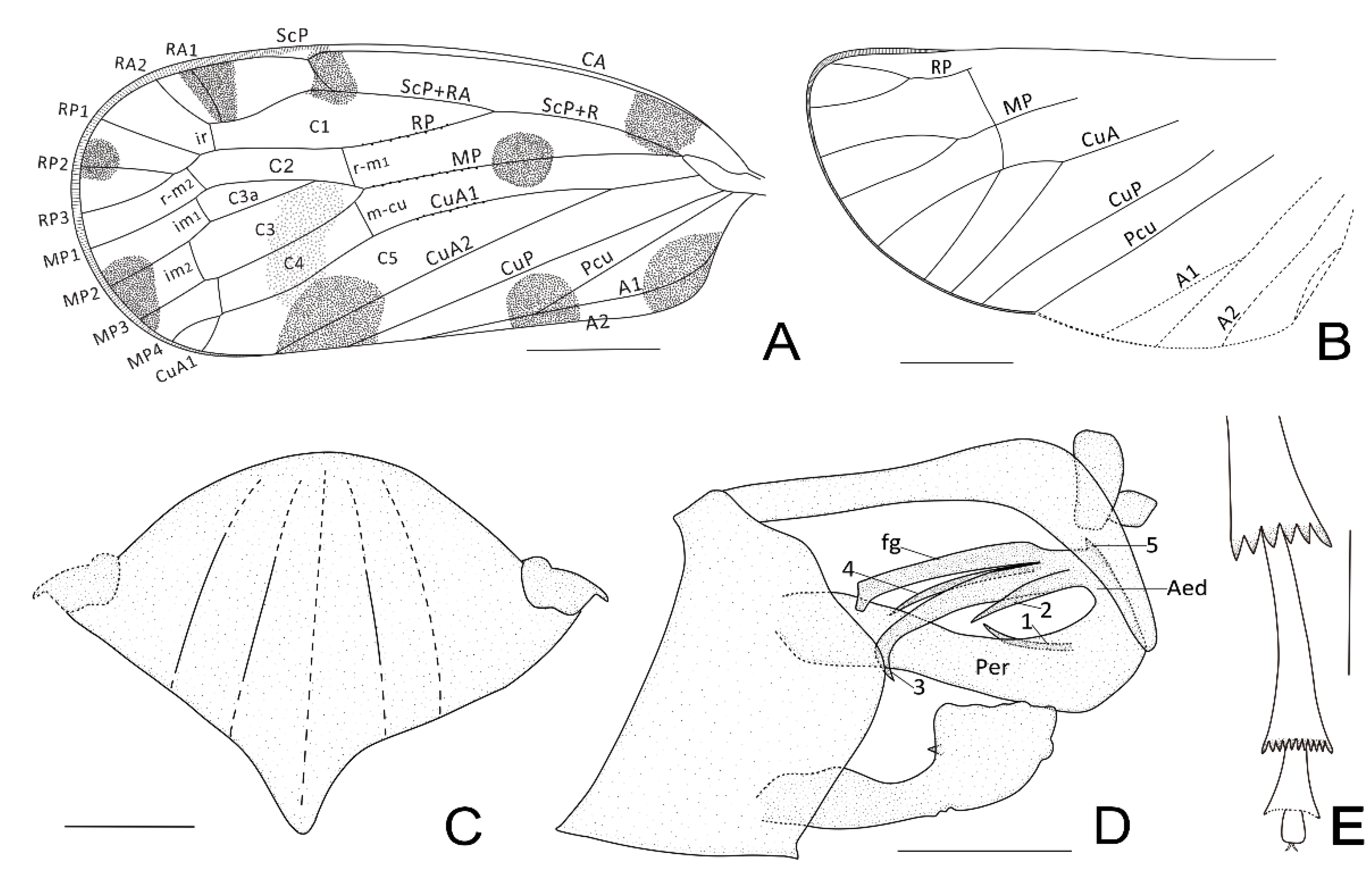
3. Systematic Paleontology
- Order: Hemiptera Linnaeus, 1758
- Suborder: Fulgoromorpha Evans, 1946
- Superfamily: Fulgoroidea Latreille, 1807
- Family: Cixiidae Spinola, 1839
- Subfamily: Cixiinae Spinola, 1839
- Tribe: Acrotiarini Bourgoin & Luo, 2021
- Genus: Maculixius Bourgoin et Wang, gen. nov.
- Type species: Maculixius jiewenae Wang et Bourgoin, sp. nov.
- LSID urn:lsid:zoobank.org:act:C206EB68-FDFF-411D-ADDA-A24D94EE2939
- Maculixius jiewenae Wang et Bourgoin, sp. nov.
- LSID urn:lsid:zoobank.org:act:B85383AC-100D-43B1-A7CB-DD0EFF6F6FDF
4. Discussion
| 1. RP with 3 terminals on forewing. Pcu and A1 merging in the middle of clavus. Stem ScP+R long, forking late after Pcu and A1 merging. CuA forking early, well before Pcu and A1 merging......Maculixius gen. nov. |
| - RP with 2 terminals on forewing. Stem ScP+R short or of medium length......2 |
| 2. Pcu and A1 merging late at 2/3 of the clavus. Stem ScP+R short, forking early before CuA, both well before Pcu and A1 merging......Pentacarinus Bourgoin & Luo 2021 |
| - Pcu and A1 merging in the middle of clavus. ScP+R and CuA forking almost or slightly before the level of Pcu and A1 merging......3 |
| 3. Pedicel of antenna elongated-ovate. Two transverse veinlets distad to RA fork between RA and RP on forewing......Delphitiara Bourgoin & Luo, 2021 |
| - Pedicel of antenna short, conical. One transverse veinlet distad to RA fork between RA and RP on forewing......Acrotiara Bourgoin & Luo, 2021 |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szwedo, J. Niryasaburnia Gen. nov. for “Liburnia” Burmitina Cockerell, 1917, from Cretaceous Myanmar (Burmese) amber (Hemiptera, Fulgoromorpha: Achilidae). J. Syst. Palaeontol. 2004, 2, 105–107. [Google Scholar] [CrossRef]
- Luo, Y.; Bourgoin, T.; Szwedo, J.; Feng, J.N. Acrotiarini trib. nov., in the Cixiidae (Insecta, Hemiptera, Fulgoromorpha) from mid-Cretaceous amber of northern Myanmar, with new insights in the classification of the family. Cretac. Res. 2021, 128, 104959. [Google Scholar] [CrossRef]
- Emeljanov, A.F.; Shcherbakov, D.E. The first Mesozoic Derbidae (Homoptera: Fulgoroidea) from Cretaceous Burmese amber. Russ. Entomol. J. 2020, 29, 237–246. [Google Scholar] [CrossRef]
- Emeljanov, A.F.; Shcherbakov, D.E. The longest-nosed Mesozoic Fulgoroidea (Homoptera): A new family from mid-Cretaceous Burmese amber. Far East. Entomol. 2018, 354, 1–14. [Google Scholar] [CrossRef]
- Song, Z.S.; Zhang, C.L.; Xi, H.Y.; Szwedo, J.; Bourgoin, T. First record of adult Dorytocidae—Dorytocus jiaxiaoae Song, Szwedo & Bourgoin sp. nov. (Hemiptera: Fulgoromorpha: Fulgoroidea) from mid-Cretaceous Kachin amber. Cretac. Res. 2021, 125, 104863. [Google Scholar] [CrossRef]
- Shcherbakov, D.E.; Emeljanov, A.F. Paradoxical derbid-like planthopper (Homoptera: Fulgoroidea) from Cretaceous Burmese amber. Russ. Entomol. J. 2021, 30, 135–139. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, D.; Yao, Y.Z. A new family Jubisentidae fam. nov. (Hemiptera: Fulgoromorpha: Fulgoroidea) from the mid-Cretaceous Burmese amber. Cretac. Res. 2019, 94, 1–7. [Google Scholar] [CrossRef]
- Luo, C.H.; Jiang, T.; Szwedo, J.; Wang, B.; Xiao, C.T. A new planthopper family Katlasidae fam. nov. (Hemiptera: Fulgoromorpha: Fulgoroidea) from mid-Cretaceous Kachin amber. Cretac. Res. 2020, 115, 104532. [Google Scholar] [CrossRef]
- Jiang, T.; Szwedo, J.; Wang, B. A giant fossil Mimarachnidae planthopper from the mid-Cretaceous Burmese amber (Hemiptera, Fulgoromorpha). Cretac. Res. 2018, 89, 183–190. [Google Scholar] [CrossRef]
- Jiang, T.; Szwedo, J.; Wang, B. A unique camouflaged mimarachnid planthopper from mid-Cretaceous Burmese amber. Sci. Rep. 2019, 9, 13112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ren, D.; Yao, Y.Z. A new genus and species of Mimarachnidae (Hemiptera: Fulgoromorpha: Fulgoroidea) from mid-Cretaceous Burmese amber. Cretac. Res. 2018, 90, 168–173. [Google Scholar] [CrossRef]
- Fu, Y.Z.; Szwedo, J.; Azar, D.; Huang, D.Y. A second species of Dachibangus (Hemiptera: Fulgoromorpha: Mimarachnidae) in mid-Cretaceous amber from northern Myanmar. Cretac. Res. 2019, 103, 104170. [Google Scholar] [CrossRef]
- Jiang, T.; Szwedo, J.; Song, Z.S.; Chen, J.; Li, Y.L.; Jiang, H. Ayaimatum trilobatum gen. et sp. nov. of Mimarachnidae (Hemiptera: Fulgoromorpha) from mid-Cretaceous amber from Kachin (northern Myanmar). Acta Palaeontol. Sin. 2020, 59, 70–85. [Google Scholar] [CrossRef]
- Fu, Y.Z.; Huang, D.Y. New mimarachnids in mid-Cretaceous amber from northern Myanmar (Hemiptera, Fulgoromorpha). Cretac. Res. 2021, 119, 104682. [Google Scholar] [CrossRef]
- Shcherbakov, D.E. Mesozoic spider mimics–Cretaceous Mimarachnidae fam. n. (Homoptera: Fulgoroidea). Russ. Entomol. J. 2007, 16, 259–264. [Google Scholar]
- Shcherbakov, D.E. An extraordinary new family of Cretaceous planthoppers (Homoptera: Fulgoroidea). Russ. Entomol. J. 2007, 16, 139–154. [Google Scholar]
- Luo, C.H.; Jiang, T.; Szwedo, J.; Wang, B.; Xiao, C.T. A new genus and species of Perforissidae (Hemiptera: Fulgoromorpha) from mid-Cretaceous Kachin amber. Cretac. Res. 2020, 114, 104518. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, D.; Yao, Y.Z. A new species of Foveopsis Shcherbakov (Hemiptera: Fulgoromorpha: Fulgoroidea: Perforissidae) from mid-Cretaceous Burmese amber. Cretac. Res. 2017, 79, 35–42. [Google Scholar] [CrossRef]
- Song, Z.S.; Xu, G.H.; Liang, A.P.; Szwedo, J.; Bourgoin, T. Still greater disparity in basal planthopper lineage: A new planthopper family Yetkhatidae (Hemiptera, Fulgoromorpha, Fulgoroidea) from mid-Cretaceous Myanmar amber. Cretac. Res. 2019, 101, 47–60. [Google Scholar] [CrossRef]
- Luo, C.H.; Wang, B.; Jarzembowski, E.A. A bizarre planthopper nymph (Hemiptera: Fulgoroidea) from mid-Cretaceous Kachin amber. Insects 2021, 12, 318. [Google Scholar] [CrossRef] [PubMed]
- Bourgoin, T. FLOW (Fulgoromorpha Lists on The Web): A World Knowledge Base Dedicated to Fulgoromorpha Version 8. Updated [10 November 2021]. Available online: http://flow.hemiptera-databases.org/flow/ (accessed on 15 December 2021).
- Shi, G.H.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.C.; Lei, W.Y.; Li, Q.L.; Li, X.H. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Bourgoin, T.; Wang, R.R.; Asche, M.; Hoch, H.; Soulier-Perkins, A.; Stroiński, A.; Yap, S.; Szwedo, J. From micropterism to hyperpterism: Recognition strategy and standardized homology-driven terminology of the forewing venation patterns in planthoppers (Hemiptera: Fulgoromorpha). Zoomorphology 2015, 134, 63–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourgoin, T. A new interpretation of the homologies of the Hemiptera male genitalia, illustrated by the Tettigometridae (Hemiptera, Fulgoromorpha). In Proceedings of the 6th Auchenorrhyncha Meeting, Turin, Italy, 7–11 September 1987. [Google Scholar]
- Löcker, B.; Fletcher, M.J.; Larivière, M.C.; Gurr, G.M. The Australian Pentastirini (Hemiptera: Fulgoromorpha: Cixiidae). Zootaxa 2006, 1290, 1–138. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Liang, F.; Bourgoin, T. A New Cixiid Fossil Genus of the Tribe Acrotiarini from Mid-Cretaceous Burmese Amber (Insecta, Hemiptera, Fulgoromorpha). Insects 2022, 13, 102. https://doi.org/10.3390/insects13010102
Wang M, Liang F, Bourgoin T. A New Cixiid Fossil Genus of the Tribe Acrotiarini from Mid-Cretaceous Burmese Amber (Insecta, Hemiptera, Fulgoromorpha). Insects. 2022; 13(1):102. https://doi.org/10.3390/insects13010102
Chicago/Turabian StyleWang, Menglin, Feiyang Liang, and Thierry Bourgoin. 2022. "A New Cixiid Fossil Genus of the Tribe Acrotiarini from Mid-Cretaceous Burmese Amber (Insecta, Hemiptera, Fulgoromorpha)" Insects 13, no. 1: 102. https://doi.org/10.3390/insects13010102
APA StyleWang, M., Liang, F., & Bourgoin, T. (2022). A New Cixiid Fossil Genus of the Tribe Acrotiarini from Mid-Cretaceous Burmese Amber (Insecta, Hemiptera, Fulgoromorpha). Insects, 13(1), 102. https://doi.org/10.3390/insects13010102