
Insect Rearing Techniques for Biological Control Programs, a Component of Sustainable Agriculture in Brazil


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Biological Control and Insect Rearing
3. Forms/Means of Rearing
4. Successful Cases in Brazil with Different Forms of Rearing the Host and the Natural Enemy
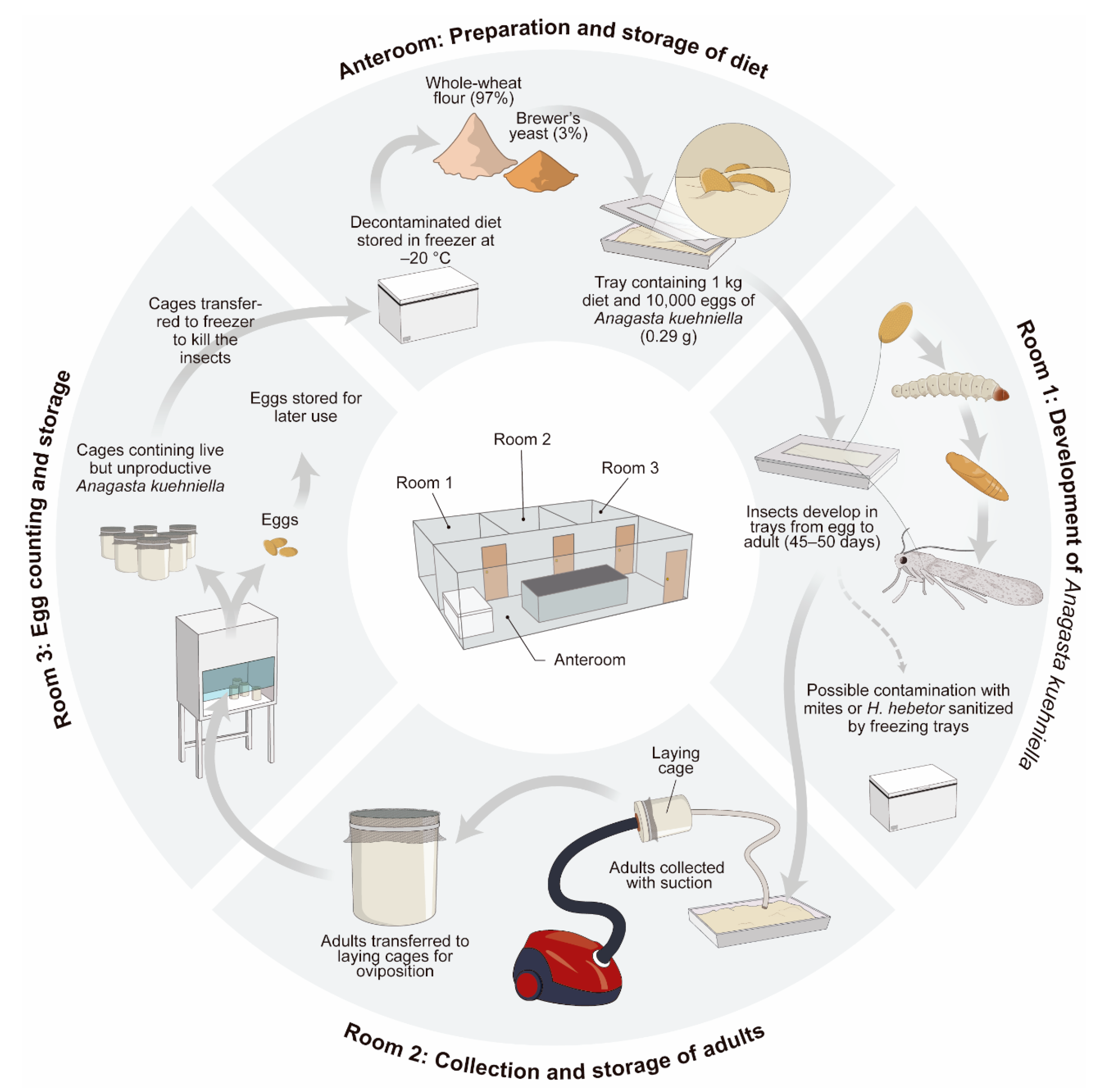
4.1. Use of a Factitious Host to Rear a Parasitoid: Case Study of Trichogramma x Lepidopterans
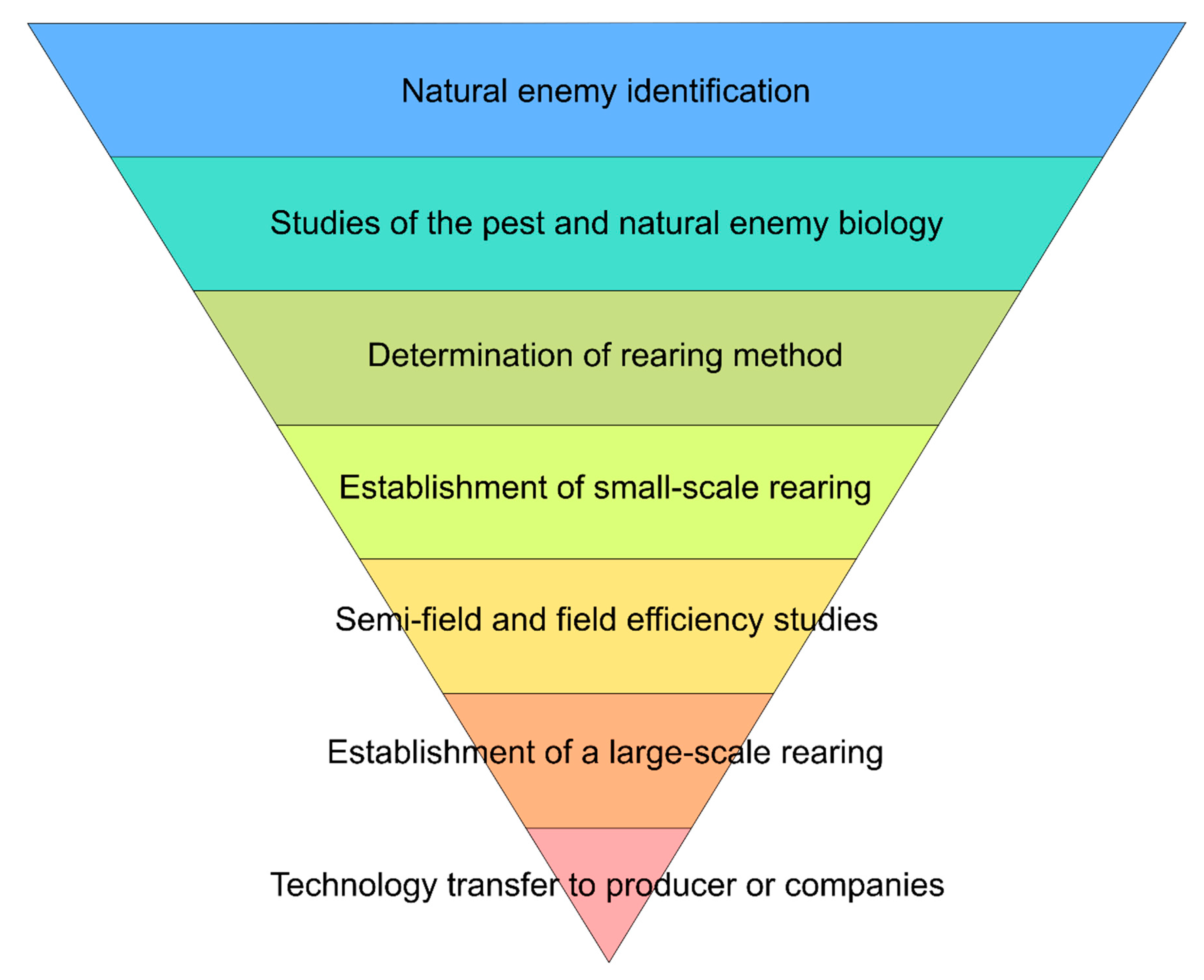
- Collection, identification, and maintenance of Trichogramma strains;
- Selection of a factitious host for mass-rearing the parasitoid;
- Biological and behavioral studies of the parasitoid and the factitious host;
- Population (eggs) dynamics of the target pest;
- Mass rearing and quality control;
- Release technique, number of parasitoids released, and release interval, as a function of plant physiology and number of release points;
- Selectivity of agrochemicals;
- Evaluation of efficiency;
- Cost/benefit ratio.
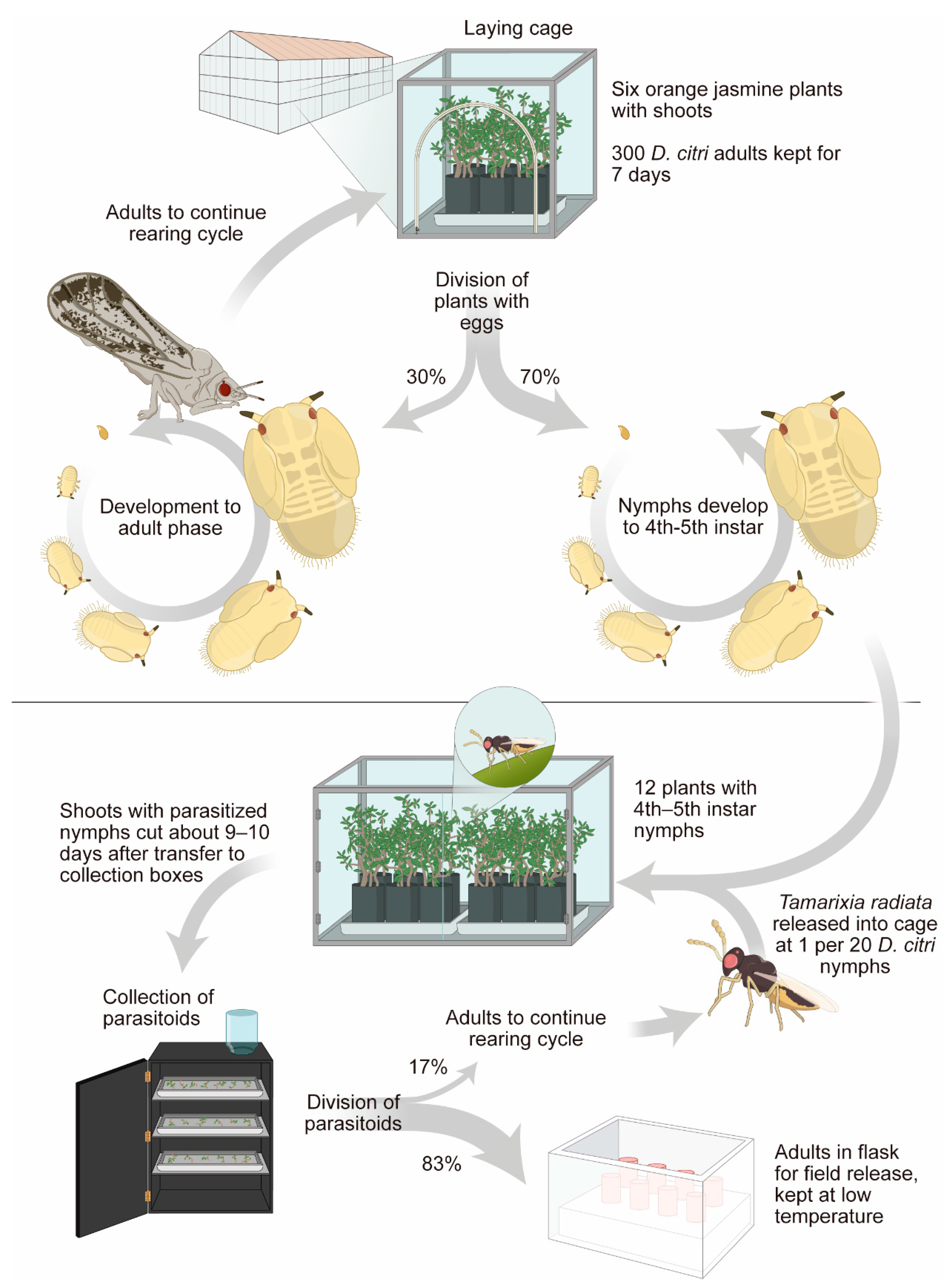
4.2. Use of a Non-Target Plant to Rear the Natural Host of Parasitoids: Example of Diaphorina citri x Tamarixia radiata
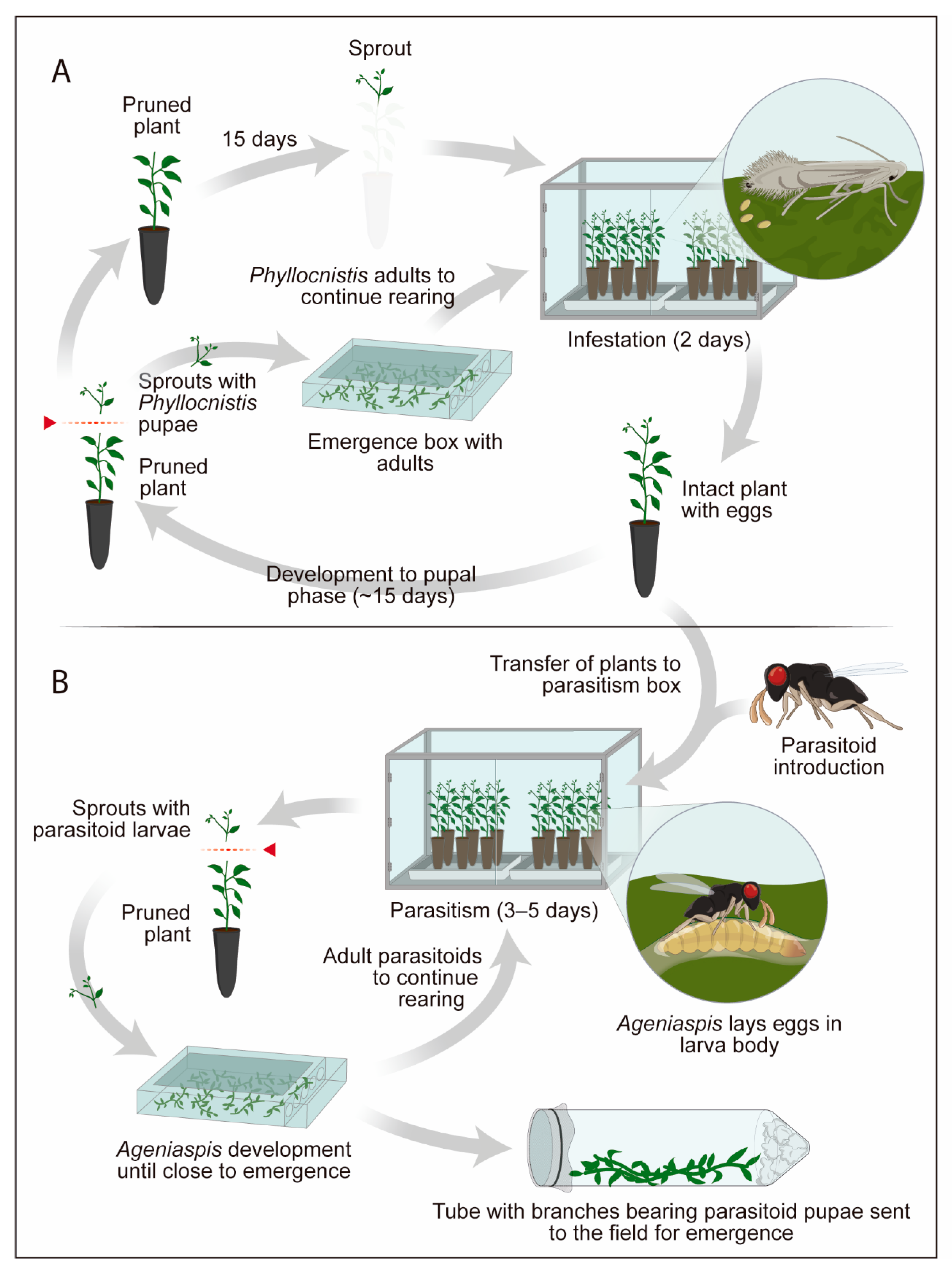
4.3. Use of Plant (Natural Host) to Rear the Pest and Produce the Parasitoid: Example of Phyllocnistis citrella x Ageniaspis citricola
4.4. Use of Natural Host Reared on Artificial Diet to Rear a Parasitoid: Example of Cotesia flavipes x Diatraea saccharalis
4.5. Use of Natural Host Reared on Artificial Diet to Rear a Parasitoid: Example of Telenomus podisi x Euchistus heros
5. Final Considerations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CONAB-Companhia Nacional de Abastecimento. 2021. Available online: https://www.conab.gov.br/info-agro/safras/graos/monitoramento-agricola (accessed on 3 November 2021).
- Parra, J.R.P. Biological control in Brazil: An overview. Sci. Agric. 2014, 71, 420–429. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Coelho, A., Jr. Applied biological control in Brazil: From laboratory assays to field application. J. Insect Sci. 2019, 19, 5. [Google Scholar]
- Smith, R.F.; Apple, J.L.; Bottrell, D.G. The origins of Integrated Pest Management concepts for agricultural crops. In Integrated Pest Management; Apple, J.L., Smith, R.F., Eds.; Springer: Boston, MA, USA, 1976; pp. 1–16. [Google Scholar]
- Mendoza, A.C.; da Rocha, A.C.; Parra, J.R. Lyophilized artificial diet for rearing the Neotropical Euschistus heros (Hemiptera: Pentatomidae). J. Insect Sci. 2016, 16, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. Biocontrol 2018, 63, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Crop Life Brazil, São Paulo, SP, Brazil. Group of Several Associations, including the Former ABCBio, the Brazilian Association of Biological Control Companies. 2021; Unpublished data.
- Li, L.Y.; Liu, W.H.; Chen, C.S.; Han, S.C.; Shin, J.C.; Du, H.S.; Feng, S.Y. In vitro rearing of Trichogramma spp. and Anastatus sp. in artificial “eggs” and the methods of mass production. Colloq. INRA 1988, 43, 339–352. [Google Scholar]
- Cônsoli, F.L.; Grenier, S. In vitro rearing of egg parasitoids. In Egg Parasitoids in Agroecosystems with Emphasis on Trichogramma. Progress in Biological Control; Cônsoli, F.L., Parra, J.R.P., Zucchi, R.A., Eds.; Springer: Dordrecht, Germany, 2009; Volume 9, pp. 293–313. [Google Scholar]
- Coudron, T.A.; Yocum, G.D.; Brandt, S.L. Nutrigenomics: A case study in the measurement of insect response to nutritional quality. Entomol. Exp. Appl. 2006, 121, 1–14. [Google Scholar] [CrossRef]
- Huynh, M.P.; Meihls, L.N.; Hibbard, B.E.; Lapointe, S.L.; Niedz, R.P.; Ludwick, D.C.; Coudron, T.A. Diet improvement for western corn rootworm (Coleoptera: Chrysomelidae) larvae. PLoS ONE 2017, 12, e0187997. [Google Scholar] [CrossRef] [Green Version]
- Huynh, M.P.; Hibbard, B.E.; Lapointe, S.L.; Niedz, R.P.; French, B.W.; Pereira, A.E.; Finke, D.L.; Shelby, K.S.; Coudron, T.A. Multidimensional approach to formulating a specialized diet for northern corn rootworm larvae. Sci. Rep. 2019, 9, 3709. [Google Scholar] [CrossRef] [Green Version]
- Diniz, A.J.F.; Garcia, A.G.; Alves, G.R.; Reigada, C.; Vieira, J.M.; Parra, J.R.P. The enemy is outside: Releasing the parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in external sources of HLB inocula to control the Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae). Neotrop. Entomol. 2020, 49, 250–257. [Google Scholar] [CrossRef]
- Smith, C.N. Insect Colonization and Mass Production; Academic Press: San Diego, CA, USA, 1966; 612p. [Google Scholar]
- Singh, P. Artificial Diets for Insects, Mites and Spiders; Plenum Press: Chicago, IL, USA, 1977; 594p. [Google Scholar]
- Singh, P.; Moore, R.F. Handbook of Insect Rearing, Vol. I and II; Elsevier: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Cohen, A.C. Insect Diets: Science and Technology, 1st ed.; CRS Press: Cleveland, IL, USA, 2003. [Google Scholar]
- Cohen, A.C. Insect Diets: Science and Technology, 2nd ed.; CRS Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Schneider, J.C. Principles and Procedures for Rearing High Quality Insects; Mississippi State University: Starkville, MS, USA, 2009. [Google Scholar]
- Panizzi, A.R.; Parra, J.R.P. Insect Bioecology and Nutrition for Integrated Pest Management; CRC Press: New York, NY, USA, 2012; 750p. [Google Scholar]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-Ilan, D.I. Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens; Academic Press: San Diego, CA, USA, 2014; 711p. [Google Scholar]
- Parra, J.R.P.; Coelho, A., Jr.; Cuervo-Rugno, J.B.; Garcia, A.G.; Moral, R.A.; Specht, A.; Dourado Neto, D. Important pest species of the Spodoptera complex: Biology, thermal requirements and ecological zoning. J. Pest Sci. 2022, 95, 169–186. [Google Scholar] [CrossRef]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. Velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- Parra, J.R.P. Mass rearing of natural enemies. In Encyclopedia of Entomology, 2nd ed.; Capinera, J.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 4, pp. 2301–2305. [Google Scholar]
- Finney, G.L.; Fisher, T.W. Culture of entomophagous insects and their hosts. In Biological Control of Pests and Weeds; de Bach, P., Schingler, E.I., Eds.; Chapman & Hall: London, UK, 1964; pp. 328–353. [Google Scholar]
- Mackauer, M. Genetic aspects of insect production. Entomophaga 1972, 17, 27–48. [Google Scholar] [CrossRef]
- Chambers, D.L. Quality control in mass rearing. Annu. Rev. Entomol. 1977, 22, 289–308. [Google Scholar] [CrossRef]
- Leppla, N.C.; Adams, F. Insect mass-rearing technology. In Principles and Applications; University of Florida: Gainesville, FL, USA, 1986; 20p. [Google Scholar]
- Ridgway, R.L.; Inscoe, M.N. Mass-reared natural enemies for pest control: Trends and challenges. In Mass-Reared Natural Enemies: Application, Regulation, and Needs; Ridgway, R.L., Hoffmann, M.P., Inscoe, M.N., Glenister, C.S., Eds.; Thomas Say Publications in Entomology: Lanham, MD, USA, 1998; pp. 1–27. [Google Scholar]
- Parra, J.R.P.; Botelho, P.S.M.; Corrêa-Ferreira, B.S.; Bento, J.M.S. Controle Biológico: Terminologia. In Controle Biológico No Brasil—Parasitóides e Predadores; Parra, J.R.P., Botelho, P.S.M., Corrêa-Ferreira, B.S., Bento, J.M.S., Eds.; Manole: Barueri, Brazil, 2002; pp. 1–13. [Google Scholar]
- Parra, J.R.P. Elaboração de programas de controle biológico: Uma visão inter e multidisciplinar. In Controle Biológico com Parasitoides e Predadores na Agricultura Brasileira; Parra, J.R.P., Pinto, A.S., Nava, D.E., Oliveira, R.C., Diniz, A.J.F., Eds.; FEALQ: Piracicaba, Brazil, 2021; pp. 39–54. [Google Scholar]
- Flanders, S.E. Biological control of the codling moth (Carpocapsa pomonella). J. Econ. Entomol. 1927, 20, 644. [Google Scholar]
- Querino, R.B.; Zucchi, R.A. Annotated checklist and illustrated key to the species of Trichogramma Westwood (Hymenoptera: Trichogrammatidae) from South America. Zootaxa 2019, 4656, 201–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.A.; Nishimura, G.; Querino, R.B.; Foerster, L.A. An integrative taxonomy of a new species of Trichogramma Westwood (Hymenoptera: Trichogrammatidae) with high reproductive capacity. Neotrop. Entomol. 2021, 50, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Parra, J.R.P.; Zucchi, R.A.; Coelho, A., Jr.; Geremias, L.D.; Cônsoli, F.L. Trichogramma as a tool for IPM in Brazil. In Augmentative Biological Control Using Trichogramma spp.: Current Status and Perspectives; Vinson, B., Greenberg, S.M., Liu, T., Rao, A., Volosciuk, L.F., Eds.; Northwest A&F University Press: Xianyang, China, 2015; pp. 472–496. [Google Scholar]
- Moraes, G.W.W.; Brun, P.G.; Soares, L.A. Insetos x Insetos: Nova alternativa para o controle de pragas. Ciênc. Hoje 1983, 1, 70–73. [Google Scholar]
- Zucchi, R.A. New species of Trichogramma (Hymenoptera: Trichogrammatidae) associated with sugar cane borer Diatraea saccharalis (F.) (Lepidoptera: Pyralidae) in Brazil. Colloq. INRA 1988, 43, 133–140. [Google Scholar]
- Botelho, P.S.M. Tabela de Vida Ecológica e Simulação da Fase Larval da Diatraea saccharalis (Fabr., 1794) (Lep.: Pyralidae). Ph.D. Thesis, Universidade de São Paulo, Piracicaba, Brazil, 1985. [Google Scholar]
- Lewis, W.J.; Nordlund, D.A.; Gross, H.R., Jr.; Perkins, W.D.; Voegelé, J. Production and performance of Trichogramma reared in eggs of Heliothis zea and other hosts. Environ. Entomol. 1976, 5, 449–452. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Coelho, A., Jr.; Geremias, L.D.; Bertin, A.; Ramos, C.J. Criação de Anagasta Kuehniella, em Pequena Escala, Para Produção de Trichogramma; Occasio: Piracicaba, Brazil, 2014; 32p. [Google Scholar]
- Gomes, S.M.; Parra, J.R.P. The parasitization as a tool for factitious host selection for Trichogramma galloi Zucchi and T. pretiosum Riley. Mitt. Biol. Bundesanst. Land Forstwirtsch. 1998, 356, 13–23. [Google Scholar]
- Strong, R.G.; Partida, G.J.; Warner, D.B. Rearing stored-product insects for laboratory studies: Six species of moths. J. Econ. Entomol. 1968, 61, 1237–1249. [Google Scholar] [CrossRef]
- Bournier, J.P.; Peyrelongue, J.Y. Introduction élevage et lachers de Trichogramma brasiliensis Ashm. (Hymenoptera: Trichogrammatidae) en vue de lutter contre Heliothis armigera Hbn. (Lepidoptera: Noctuidae) a Madagascar. Coton Fibres Trop. 1973, 28, 231–237. [Google Scholar]
- Daumal, J.; Marconi, D.; Chassain, C. Dispositif d’élevage miniaturisé et automatisé d’ Ephestia kuehniella Zeller (Lepidoptera—Pyralidae). Bull. Soc. Linn. Lyon 1985, 54, 7–12. [Google Scholar] [CrossRef]
- Daumal, J.; Voegelé, J.; Brun, P. Les Trichogrammes: II. Unité de production massive et quotidienne d’un hôte de substitution Ephestia kuehniella Zell. (Lepidoptera, Pyralidae). Ann. Zool. Ecol. Anim. 1975, 7, 45–49. [Google Scholar]
- Parra, J.R.P.; Lopes, J.R.S.; Serra, H.J.P.; Sales, O., Jr. Metodologia de criação de Anagasta kuehniella (Zeller, 1879) para produção massal de Trichogramma spp. An. Soc. Entomol. Bras. 1989, 18, 403–415. [Google Scholar] [CrossRef]
- Stein, C.P.; Parra, J.R.P. Aspectos biológicos de Anagasta kuehniella (Zeller, 1879) criada em dois substratos alimentares. An. Soc. Entomol. Bras. 1987, 16, 173–185. [Google Scholar] [CrossRef]
- Coelho, A., Jr. Otimização da Criação de Anagasta kuehniella (Zeller, 1879), Hospedeiro Alternativo de Trichogramma spp., Baseando-Se na Temperatura, Densidade Larval e Concentração de Dióxido de Carbono. Master’s Thesis, Universidade de São Paulo, Piracicaba, Brazil, 2011. [Google Scholar]
- Coelho, A., Jr.; Parra, J.R.P. The effect of rearing in different temperature regimes on the reproduction of Anagasta kuehniella (Lepidoptera: Pyralidae). Environ. Entomol. 2013, 42, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, F.; Bigler, F.; Eden, G.; Bosshart, S. Optimal larval density and quality control aspects in mass rearing of the Mediterranean flour moth, Ephestia kuehniella Zeller (Lepidoptera: Phycitidae). J. Appl. Entomol. 1992, 114, 353–361. [Google Scholar] [CrossRef]
- Coelho, A., Jr.; Reigada, C.; Haddad, M.L.; Parra, J.R.P. Effects of temperature increase caused by larval metabolism on the reproduction of Anagasta kuehniella (Lepidoptera: Pyralidae), a factitious host for Trichogramma. Biocontrol Sci. Technol. 2016, 26, 630–639. [Google Scholar] [CrossRef]
- Coelho, A., Jr.; Parra, J.R.P. Effect of carbon dioxide (CO2) on mortality and reproduction of Anagasta kuehniella (Zeller, 1879), in mass rearing, aiming at the production of Trichogramma spp. An. Acad. Bras. Ciênc. 2013, 85, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Parra, J.R.P.; Coelho, A., Jr. Trichogramma spp. um caso de sucesso de controle biológico aplicado no Brasil. In Controle Biológico com Parasitoides e Predadores na Agricultura Brasileira; Parra, J.R.P., Pinto, A.S., Nava, D.E., Oliveira, R.C., Diniz, A.J.F., Eds.; FEALQ: Piracicaba, Brazil, 2021; pp. 159–202. [Google Scholar]
- Querino, R.B.; Zucchi, R.A. Guia de Identificação de Trichogramma Para o Brasil; Embrapa Informação Tecnológica: Brasília, Brazil, 2011; 103p. [Google Scholar]
- Ciociola, A.I., Jr.; Almeida, R.P.; Zucchi, R.A.; Stouthamer, R. Detecção de Wolbachia em uma população telítoca de Trichogramma atopovirilia Oatman & Platner (Hymenoptera: Trichogrammatidae) via PCR com o primer específico wsp. Neotrop. Entomol. 2001, 30, 489–491. [Google Scholar]
- Nansen, C.; Coelho, A., Jr.; Vieira, J.M.; Parra, J.R.P. Reflectance-based identification of parasitized host eggs and adult Trichogramma specimens. J. Exp. Biol. 2014, 217, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Bleicher, E.; Parra, J.R.P. Espécies de Trichogramma parasitóides de Alabama argillacea II: Tabela de vida de fertilidade e parasitismo de três populações. Pesqui. Agropecu. Bras. 1990, 25, 207–214. [Google Scholar]
- Cerutti, F.; Bigler, F. Quality assessment of Trichogramma brassicae in the laboratory. Entomol. Exp. Appl. 1995, 75, 19–26. [Google Scholar] [CrossRef]
- Sorati, M.; Newman, M.; Hoffmann, A.A. Inbreeding and incompatibility in Trichogramma brassicae: Evidence and implications for quality control. Entomol. Exp. Appl. 1996, 78, 283–290. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Laboratory fecundity as predictor of field success in Trichogramma carverae (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2002, 95, 912–917. [Google Scholar] [CrossRef]
- Guzmán-Larralde, A.; Cerna-Chávez, E.; Rodríguez-Campos, E.; Loyola-Licea, J.C.; Stouthamer, R. Genetic variation and the performance of a mass-reared parasitoid, Trichogramma pretiosum (Hymenoptera: Trichogrammatidae), in laboratory trials. J. Appl. Entomol. 2014, 138, 315–391. [Google Scholar] [CrossRef]
- Coelho, A., Jr.; Rugman-Jones, P.F.; Reigada, C.; Stouthamer, R.; Parra, J.R. Laboratory performance predicts the success of field releases in inbred lines of the egg parasitoid Trichogramma pretiosum (Hymenoptera: Trichogrammatidae). PLoS ONE 2016, 11, e0146153. [Google Scholar]
- Coelho, A., Jr.; Stouthamer, R.; Parra, J.R.P. Flight Propensity of Isofemale Lines of Trichogramma pretiosum Riley in Two Relative Humidity Levels. Fla. Entomol. 2018, 101, 364–368. [Google Scholar] [CrossRef]
- Prezotti, L.; Parra, J.R.P.; Vencovsky, R.; Coelho, A.S.G.; Cruz, I. Effect of the size of the founder population on the quality of sexual populations of Trichogramma pretiosum in laboratory. Biol. Control 2004, 30, 174–180. [Google Scholar] [CrossRef]
- Dutton, A.; Bigler, F. Flight activity assessment of the egg parasitoid Trichogramma brassicae (Hym.: Trichogrammatidae) in laboratory and field conditions. Entomophaga 1995, 40, 223–233. [Google Scholar] [CrossRef]
- Prezotti, L.; Parra, J.R.P.; Vencovsky, R.; Dias, C.T.D.S.; Cruz, I.; Chagas, M.C.M. Teste de vôo como critério de avaliação da qualidade de Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae): Adaptação de metodologia. Neotrop. Entomol. 2002, 31, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Danks, H.V. Systematics in support of entomology. Annu. Rev. Entomol. 1988, 33, 271–294. [Google Scholar] [CrossRef]
- Coelho, A., Jr.; Geremias, L.D.; Alves, G.R.; Da Rocha, A.C.P.; Parra, J.R.P. The biology of Trichogramma pretiosum as atmospheric O2 becomes depleted and CO2 accumulates. Biol. Control 2017, 105, 1–5. [Google Scholar] [CrossRef]
- Parra, J.R.P. Técnicas de criação de Anagasta kuehniella, hospedeiro alternativo para a produção de Trichogramma. In Trichogramma e o Controle Biológico Aplicado; Parra, J.R.P., Zucchi, R.A., Eds.; FEALQ: Piracicaba, Brazil, 1997; pp. 121–150. [Google Scholar]
- Molina, R.M.S.; Fronza, V.; Parra, J.R.P. Seleção de Trichogramma spp., para o controle de Ecdytolopha aurantiana, com base na biologia e exigências térmicas. Rev. Bras. Entomol. 2005, 49, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Parra, J.R.P.; Geremias, L.D.; Bertin, A.; Colmenarez, Y.C.; Coelho, A., Jr. Small-Scale Rearing of Anagasta Kuehniella for Trichogramma Production; CABI: Wallingford, UK, 2021. [Google Scholar]
- Lopes, J.R.S.; Parra, J.R.P. Efeito da idade do ovo do hospedeiro natural e alternativo no desenvolvimento e parasitismo de duas espécies de Trichogramma. Rev. Agric. 1991, 66, 221–244. [Google Scholar]
- Pak, G.A.; Oatman, E.R. Biology of Trichogramma brevicapillum. Entomol. Exp. Appl. 1982, 32, 61–67. [Google Scholar] [CrossRef]
- Sá, L.A.N. Bioecologia de Trichogramma Pretiosum Riley, 1879, Visando Avaliar o Seu Potencial Para Controle de Spodoptera frugiperda (J. E. Smith, 1797) e Helicoverpa zea (Boddie, 1850) em Milho. Ph.D. Thesis, Universidade de São Paulo, Piracicaba, Brazil, 1991. [Google Scholar]
- Neil, K.A.; Specht, H.B. Field releases of Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae) for suppression of corn earworm, Heliothis zea (Lepidoptera: Noctuidae), egg populations on sweet corn in Nova Scotia. Can. Entomol. 1990, 122, 1259–1266. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Alves, G.R.; Diniz, A.J.F.; Vieira, J.M. Tamarixia radiata (Hymenoptera: Eulophidae) × Diaphorina citri (Hemiptera: Liviidae): Mass rearing and potential use of the parasitoid in Brazil. J. Integr. Pest Manag. 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Marin, D.R. Densidade e Raio de Dispersão de Tamarixia Radiata (Waterston) (Hymenoptera: Eulophidae) Para o Controle de Diaphorina Citri Kuwayama (Hemiptera: Liviidae) em Pomares Cítricos. Ph.D. Thesis, Fundo de defesa da Citricultura (FUNDECITRUS), Araraquara, Brazil, 2021. [Google Scholar]
- Diniz, A.J.F.; Alves, G.R.; Vieira, J.M.; Parra, J.R.P. Manejo do psilídeo dos citros com uso do parasitoide Tamarixia radiata: Uma nova abordagem em controle biológico. In Controle Biológico com Parasitoides e Predadores na Agricultura Brasileira; Parra, J.R.P., Pinto, A.S., Nava, D.E., Oliveira, R.C., Diniz, A.J.F., Eds.; FEALQ: Piracicaba, Brazil, 2021; pp. 127–158. [Google Scholar]
- Nava, D.E.; Torres, M.L.G.; Rodrigues, M.D.L.; Bento, J.M.S.; Parra, J.R.P. Biology of Diaphorina citri (Hem.: Psyllidae) on different hosts and at different temperatures. J. Appl. Entomol. 2007, 131, 709–715. [Google Scholar] [CrossRef]
- Dal Pogetto, L. Potencial Hospedeiro de Duas Espécies de Murraya Para Diaphorina citri (Kuwayama, 1908) (Hemiptera: Liviidae) com Vistas à Produção de Tamarixia Radiata Waterston, 1922 (Hymenoptera: Eulophidae). Master’s Thesis, Universidade de São Paulo, Piracicaba, Brazil, 2018. [Google Scholar]
- Diniz, A.J.F. Otimização da Criação de Diaphorina citri Kuwayama, 1908 (Hemiptera: Liviidae) e de Tamarixia Radiata (Waterston, 1922) (Hymenoptera: Eulophidae), Visando a Produção em Larga Escala do Parasitoide e Avaliação do seu Estababelecimento em Campo. Ph.D. Thesis, Universidade de São Paulo, Piracicaba, Brazil, 2013. [Google Scholar]
- Parra, J.R.P.; Diniz, A.J.F.; Vieira, J.M.; Alves, G.R. Utilização do Parasitoide Tamarixia Radiata como Componente do Manejo Integrado do Huanglongbing; Fundecitrus: Ararquara, Brazil, 2017. [Google Scholar]
- Alves, G.R.; Diniz, A.J.F.; Parra, J.R.P. Biology of the huanglongbing vector Diaphorina citri (Hemiptera: Liviidae) on different host plants. J. Econ. Entomol. 2014, 107, 691–696. [Google Scholar] [CrossRef]
- Feichtenberger, E.; Raga, A. First report of citrus leafminer Phyllocnistis citrella (Lep.: Gracillariidae) in Brazil. Proc. Reun. Interam. De Hortic. Trop. 1996, 42, 445. [Google Scholar]
- Parra, J.R.P. Casos de sucesso de CB, com parasitoides e predadores no Brasil, em diferentes culturas. In Controle Biológico com Parasitoides e Predadores na Agricultura Brasileira; Parra, J.R.P., Pinto, A.S., Nava, D.E., Oliveira, R.C., Diniz, A.J.F., Eds.; FEALQ: Piracicaba, Brazil, 2021; pp. 77–98. [Google Scholar]
- Chagas, M.C.M.; Parra, J.R.P.; Milano, P.; Nascimento, A.M.; Parra, A.L.G.C.; Yamamoto, P.T. Ageniaspis citricola: Criação e estabelecimento no Brasil. In Controle Biológico no Brasil: Parasitóides e Predadores; Parra, J.R.P., Botelho, P.S.M., Corrêa-Ferreira, B.S., Bento, J.M.S., Eds.; Manole: São Paulo, Brazil, 2002; pp. 377–391. [Google Scholar]
- Hensley, S.D.; Hammond, A.M., Jr. Laboratory techniques for rearing the sugarcane borer on an artificial diet. J. Econ. Entomol. 1968, 61, 1742–1743. [Google Scholar] [CrossRef]
- Gallo, D.; Windle, F.M.; Willians, R.N.; Berti Filho, E. Método de criação artificial da broca de cana de açúcar para emprego seu controle. In Anais, Proceedings of the Reunião Anual de Entomologia; Sociedade Entomológica do Brasil: Recife, PE, Brazil, 1969; pp. 4–5. [Google Scholar]
- Botelho, P.S.M.; Macedo, N. Cotesia flavipes para o controle de Diatraea saccharalis. In Controle Biológico No Brasil: Parasitóides e Predadores; Parra, J.R.P., Botelho, P.S.M., Corrêa-Ferreira, B.S., Bento, J.M.S., Eds.; Manole: São Paulo, Brazil, 2002; pp. 409–429. [Google Scholar]
- Sgrillo, R.B.; Walder, J.M.M.; Wiendl, F.M. Progressos na criação da broca da cana-de-açúcar, Diatraea saccharalis (F.) realizados no Centro de Energia Nuclear na Agricultura—CENA. O Solo 1977, 69, 58–60. [Google Scholar]
- Mendonça Filho, A. Criação artificial em laboratório dos parasitos da broca da cana-de-açúcar (Diatraea spp.) (Lep., Crambidae). Brasil Açuc. 1973, 81, 47–80. [Google Scholar]
- Araújo, J.R.; Botelho, P.S.M.; Araújo, S.M.S.S.; Almeida, L.C.; Degaspari, N. Nova dieta para criação da Diatraea saccharalis (Fabr.). Saccharum APC, Ver. Tecnol. Indúst. Açuc. Alcool. 1985, 36, 45–48. [Google Scholar]
- Parra, J.R.P.; Mihsfeldt, L.H. Comparison of artificial diets for rearing the sugarcane borer. In Advances in Insect Rearing for Research & Pest Management; Anderson, T.E., Leppla, N.C., Eds.; Westview Press: San Francisco, CA, USA, 1992; pp. 195–209. [Google Scholar]
- Poitout, S.; Bues, R. Elevage de plusieurs espèces de Lépidoptères Noctuidae sur milieu artificiel riche et sur milieu artificiel simplifié. Ann. Zool. Ecol. Anim. 1970, 2, 79–91. [Google Scholar]
- Parra, J.R.P. Técnicas de Criação de Insetos Para Programas de Controle Biológico; University of São Paulo-Luiz de Queiroz College of Agriculture: Piracicaba, SP, Brazil, 2013; Unpublished work. [Google Scholar]
- Alves, S.B.; Botelho, P.S.M.; Macedo, N.; Riscado, G.M. The occurrence of some pathogens on sugar cane borer Diatraea saccharalis in field and laboratory conditions. Entomol. News 1985, 17, 4–5. [Google Scholar]
- Simões, R.A.; Reis, L.G.; Bento, J.M.; Solter, L.F.; Delalibera, I., Jr. Biological and behavioral parameters of the parasitoid Cotesia flavipes (Hymenoptera: Braconidae) are altered by the pathogen Nosema sp. (Microsporidia: Nosematidae). Biol. Control 2012, 63, 164–171. [Google Scholar] [CrossRef]
- Corrêa-Ferreira, B.S. Trissolcus basalis para o controle de percevejos da soja. In Controle Biológico no Brasil: Parasitóides e Predadores; Parra, J.R.P., Botelho, P.S.M., Corrêa-Ferreira, B.S., Bento, J.M.S., Eds.; Manole: São Paulo, Brazil, 2002; pp. 449–471. [Google Scholar]
- Corrêa-Ferreira, B.S.; Moscardi, F. Seasonal occurrence and host spectrum of egg parasitoids associated with soybean stink bugs. Biol. Control 1995, 5, 196–202. [Google Scholar] [CrossRef]
- Panizzi, A.R.; Parra, J.R.P.; Santos, C.H.; Carvalho, D.R. Rearing the southern green stink bug using artificial dry diet and artificial plant. Pesqui. Agropecu. Bras. 2000, 35, 1709–1715. [Google Scholar] [CrossRef] [Green Version]
- Kamano, S. Artificial diet for rearing bean bug, Riptortus clavatus Thunberg. Jpn. J. Appl. Entomol. 1980, 24, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Fortes, P.; Magro, S.R.; Panizzi, A.R.; Parra, J.R.P. Development of a dry artificial diet for Nezara viridula (L.) and Euschistus heros (Fabricius) (Heteroptera: Pentatomidae). Neotrop. Entomol. 2006, 35, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, F. Adição de Frutos de Ligustrum Lucidum Ait. (Oleaceae) em Dieta Artificial Seca Para a Criação do Percevejo-marrom Euschistus heros (F.) (Heteroptera: Pentatomidae). Master’s Thesis, Universidade Federal do Paraná, Curitiba, Brazil, 2007. [Google Scholar]
- Rocha, A.C.P.D. Dinâmica de Interações Tróficas de Euschistus heros (Hemiptera: Pentatomidae) com Telenomus podisi e Trissolcus basalis (Hymenoptera: Platygastridae). Ph.D. Thesis, Universidade de São Paulo, Piracicaba, Brazil, 2016. [Google Scholar]
- Garcia, M.S.; Parra, J.R. Comparison of several artificial diets with different protein sources for massal rearing of Ecdytolopha aurantiana (Lima) (Lepidoptera: Tortricidae). An. Soc. Entomol. Bras. 1999, 28, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Parra, J.R.P.; Bento, J.M.S.; Garcia, M.S.; Yamamoto, P.T.; Vilela, E.F.; Leal, W.S. Development of a control alternative for the citrus fruit borer, Ecdytolopha aurantiana (Lepidoptera, Tortricidae): From basic research to the grower. Rev. Bras. Entomol. 2004, 48, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Bento, J.M.S.; Parra, J.R.P.; de Miranda, S.H.; Adami, A.C.; Vilela, E.F.; Leal, W.S. How much is a pheromone worth? F1000Research 2016, 5, 1763. [Google Scholar] [CrossRef] [Green Version]
- Bolfe, É.L.; Jorge, L.A.D.C.; Sanches, I.D.A.; Luchiari, A., Jr.; da Costa, C.C.; Victoria, D.D.C.; Inamasu, R.Y.; Grego, C.R.; Ferreira, V.R.; Ramirez, A.R. Precision and digital agriculture: Adoption of technologies and perception of Brazilian farmers. Agriculture 2020, 10, 653. [Google Scholar] [CrossRef]
- Nansen, C.; Geremias, L.D.; Xue, Y.; Huang, F.; Parra, J.R. Agricultural case studies of classification accuracy, spectral resolution, and model over-fitting. Appl. Spectrosc. 2013, 67, 1332–1338. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra, J.R.P.; Coelho, A., Jr. Insect Rearing Techniques for Biological Control Programs, a Component of Sustainable Agriculture in Brazil. Insects 2022, 13, 105. https://doi.org/10.3390/insects13010105
Parra JRP, Coelho A Jr. Insect Rearing Techniques for Biological Control Programs, a Component of Sustainable Agriculture in Brazil. Insects. 2022; 13(1):105. https://doi.org/10.3390/insects13010105
Chicago/Turabian StyleParra, José Roberto Postali, and Aloisio Coelho, Jr. 2022. "Insect Rearing Techniques for Biological Control Programs, a Component of Sustainable Agriculture in Brazil" Insects 13, no. 1: 105. https://doi.org/10.3390/insects13010105
APA StyleParra, J. R. P., & Coelho, A., Jr. (2022). Insect Rearing Techniques for Biological Control Programs, a Component of Sustainable Agriculture in Brazil. Insects, 13(1), 105. https://doi.org/10.3390/insects13010105