
Use of Pyriproxyfen to Induce Oogenesis in Diapausing Megacopta cribraria (Heteroptera: Plataspidae), and Evaluation of Pyriproxyfen-Induced Eggs for Rearing the Parasitoid Paratelenomus saccharalis (Hymenoptera: Scelionidae)

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Megacopta cribraria Collection
2.2. Pyriproxyfen and Photoperiod Treatments
2.3. Megacopta cribraria Survival, Oviposition and Egg Viability
2.4. Parasitism of Eggs from Treated Females of Megacopta cribraria
2.5. Parasitoid Yield Estimates
3. Results
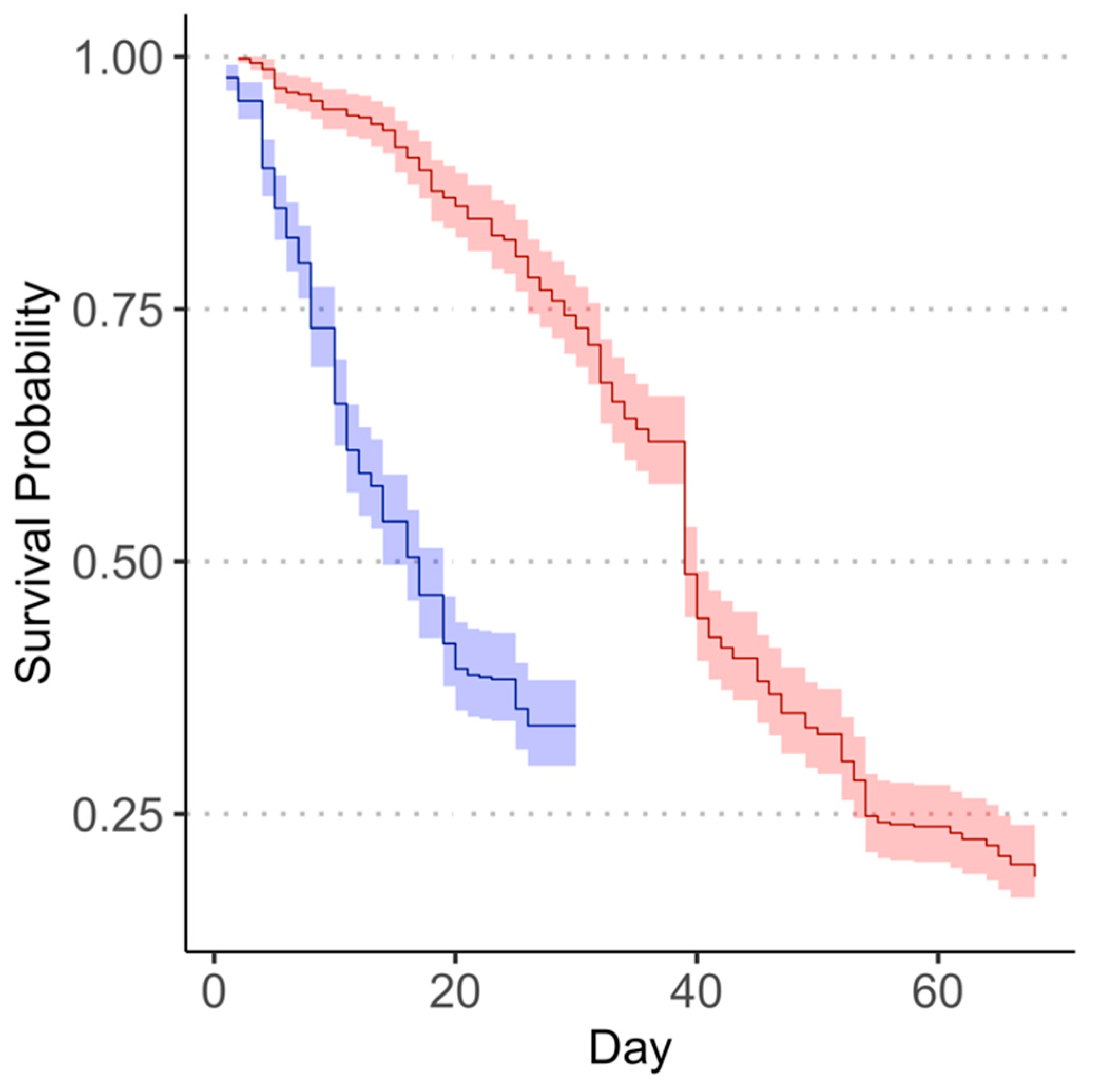
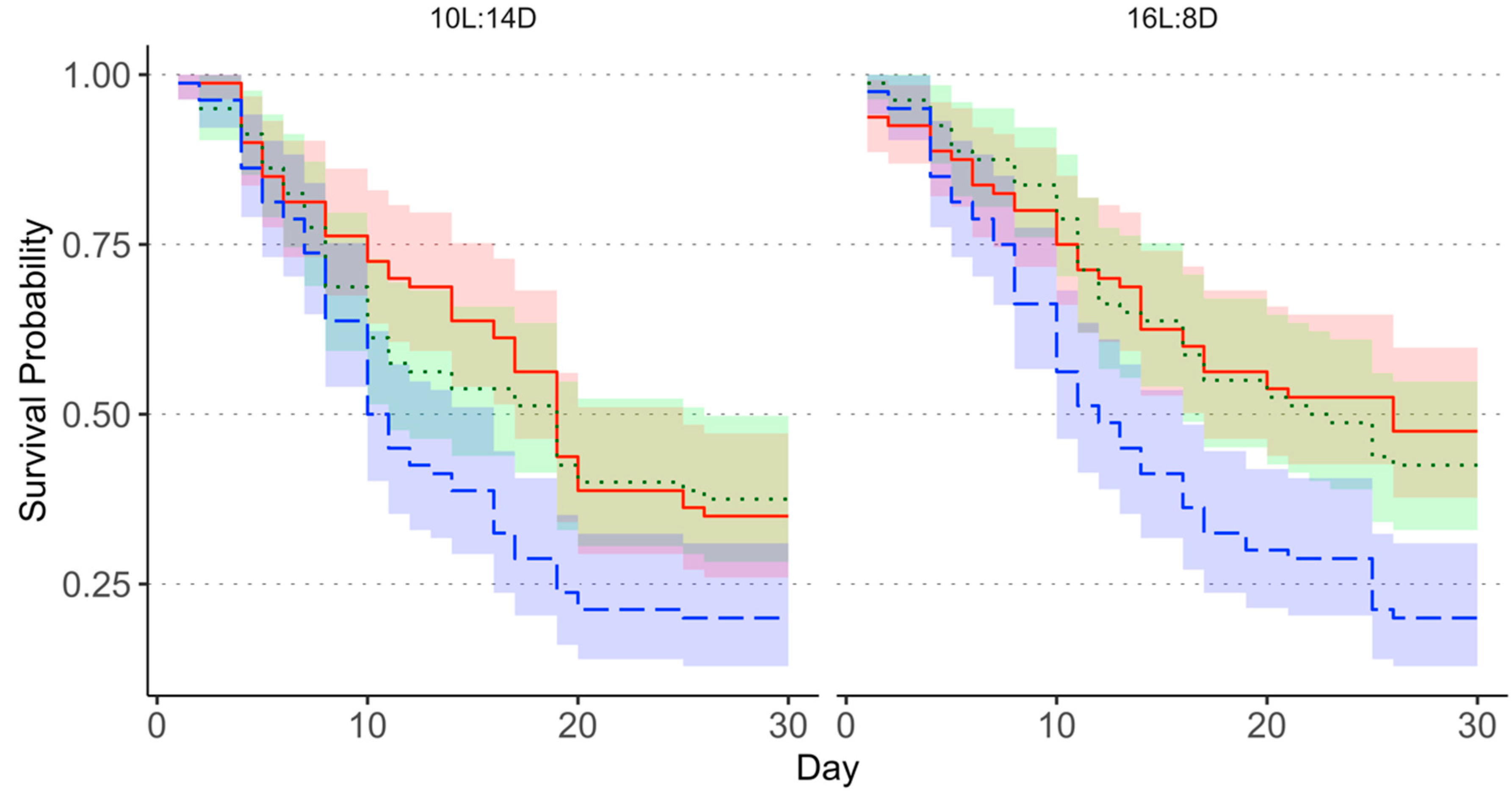
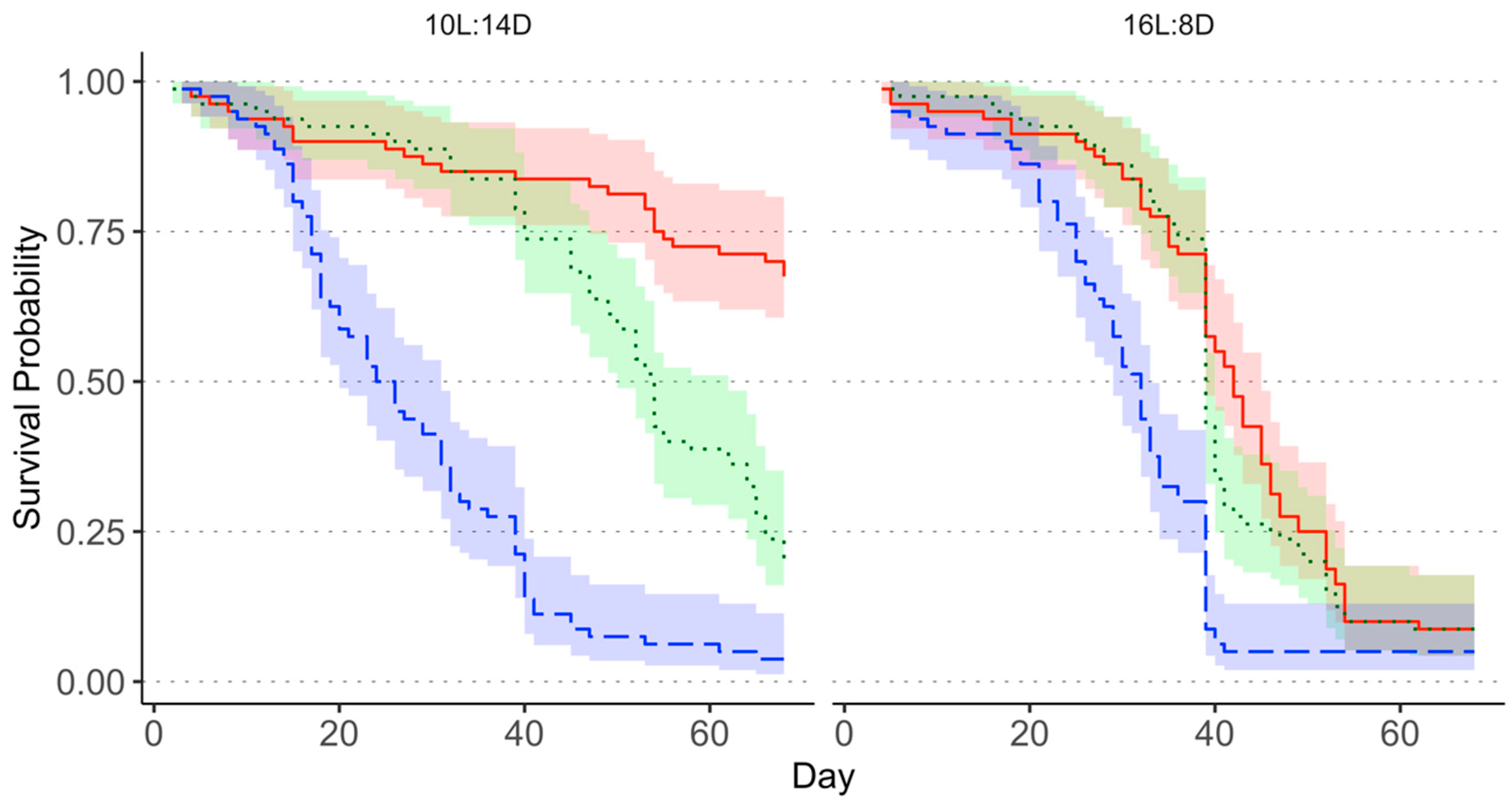
3.1. Survival of Adult Megacopta cribraria
3.2. Egg Production and Eclosion of Megacopta cribraria and Paratelenomus saccharalis
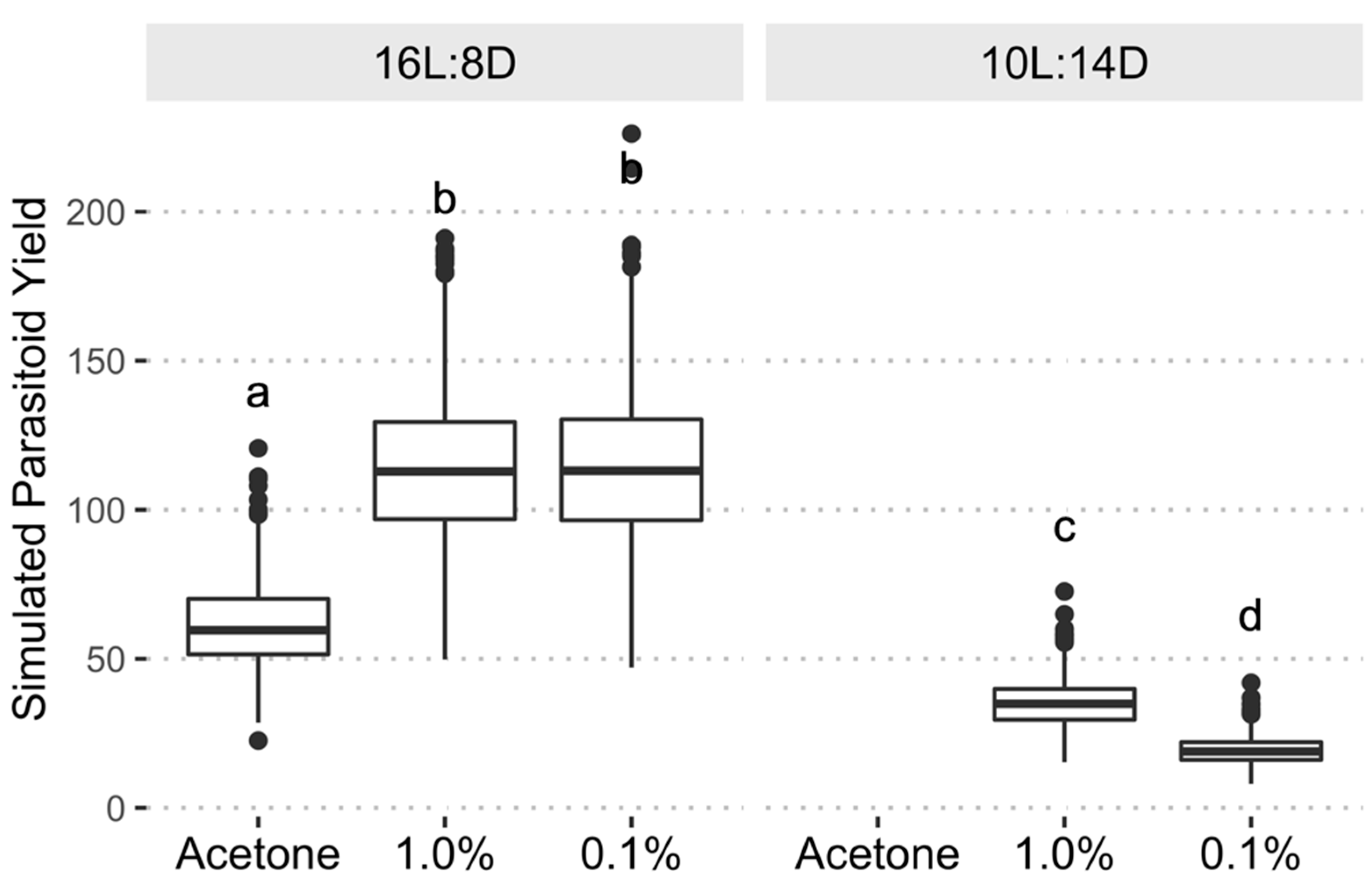
3.3. Parasitoid Yield
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corrêa-Ferreira, B.S.; Moscardi, F. Biological control of soybean stink bugs by inoculative releases of Trissolcus basalis. Entomol. Exp. Appl. 1996, 79, 1–7. [Google Scholar] [CrossRef]
- Smith, S.M. Biological control with Trichogramma: Advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41, 375–406. [Google Scholar] [CrossRef]
- Waage, J.K.; Hassell, M.P. Parasitoids as biological control agents–a fundamental approach. Parasitology 1982, 84, 241. [Google Scholar] [CrossRef]
- Mills, N. Egg parasitoids in biological control and integrated pest management. In Egg Parasitoids in Agroecosystems with Emphasis on Trichogramma; Springer: Dordrecht, The Netherlands, 2009; pp. 389–411. [Google Scholar]
- Mills, N. Selecting effective parasitoids for biological control introductions: Codling moth as a case study. Biol. Control 2005, 34, 274–282. [Google Scholar] [CrossRef]
- Gardner, W.A.; Blount, J.L.; Golec, J.R.; Jones, W.A.; Hu, X.P.; Talamas, E.J.; Evans, R.M.; Dong, X.; Ray, C.H.; Buntin, G.D.; et al. Discovery of Paratelenomus saccharalis (Dodd) (Hymenoptera: Platygastridae), an egg parasitoid of Megacopta cribraria F. (Hemiptera: Plataspidae) in its expanded North American range. J. Entomol. Sci. 2013, 48, 355–359. [Google Scholar] [CrossRef] [Green Version]
- Ruberson, J.R.; Takasu, K.; David Buntin, G.; Eger, J.E.; Gardner, W.A.; Greene, J.K.; Jenkins, T.M.; Jones, W.A.; Olson, D.M.; Roberts, P.M.; et al. From Asian curiosity to eruptive American pest: Megacopta cribraria (Hemiptera: Plataspidae) and prospects for its biological control. Appl. Entomol. Zool. 2013, 48, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.Q.; Yao, Y.X.; Qiu, L.F.; Li, Z.X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Entomol. Soc. Am. 2009, 102, 39–47. [Google Scholar] [CrossRef]
- Sforza, R.F.H.; Bon, M.-C.; Martel, G.; Augé, M.; Roche, M.; Mahmood, R.; Smith, L. Initial evaluation of two native egg parasitoids for the control of Bagrada hilaris, and invasive stink bug in Western USA. In Proceedings of the 5th International Symposium on Biological Control of Arthropods, Langkawi, Malaysia, 11–15 September 2017; Mason, P.G., Gillespie, D.R., Vincent, C., Eds.; CAB International: Langkawi, Malaysia, 2017; p. 221. [Google Scholar]
- Leskey, T.C.; Hamilton, G.C.; Nielsen, A.L.; Polk, D.F.; Rodriguez-Saona, C.; Christopher Bergh, J.; Ames Herbert, D.; Kuhar, T.P.; Pfeiffer, D.; Dively, G.P.; et al. Pest status of the brown marmorated stink bug, Halyomorpha halys in the USA. Outlooks Pest Manag. 2012, 23, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Davey, K.G. Hormonal controls on reproduction in female heteroptera. Arch. Insect Biochem. Physiol. 1997, 35, 443–453. [Google Scholar] [CrossRef]
- Saulich, A.K.; Musolin, D.L. Diapause in the seasonal cycle of stink bugs (Heteroptera, Pentatomidae) from the Temperate Zone. Entomol. Rev. 2012, 92, 1–26. [Google Scholar] [CrossRef]
- Musolin, D.L. Insects in a warmer world: Ecological, physiological and life-history responses of true bugs (Heteroptera) to climate change. Glob. Chang. Biol. 2007, 13, 1565–1585. [Google Scholar] [CrossRef]
- Numata, H.; Hidaka, T. Termination of adult diapause by a juvenile hormone analogue in the bean bug, Riptortus clavatus. Zool. Sci. 1984, 1, 751–754. [Google Scholar]
- Amiri, A.; Bandani, A.R.; Darvishzadeh, A. Effects of the insect growth regulators methoxyfenozide and pyriproxyfen on adult diapause in sunn pest Eurygaster integriceps (Hemiptera: Scutelleridae). J. Agric. Sci. Tech. 2012, 14, 1205–1218. [Google Scholar]
- Penca, C.; Hodges, A.C. Pyriproxyfen treatment terminates Halyomorpha halys reproductive diapause, with an indirect mortality effect on its egg parasitoid Trissolcus japonicus. J. Pest. Sci. 2017, 90, 1269–1276. [Google Scholar] [CrossRef]
- Hoshino, K.; Adati, T.; Olson, D.M.; Takasu, K. Seasonal occurrence and interspecific interactions of egg parasitoids of megacopta cribraria (heteroptera: Plataspidae) in Japan. Environ. Entomol. 2017, 46, 487–493. [Google Scholar] [CrossRef]
- Suiter, D.R.; Eger, J.E.; Gardner, W.A.; Kemerait, R.C.; All, J.N.; Roberts, P.M.; Greene, J.K.; Ames, L.M.; Buntin, G.D.; Jenkins, T.M.; et al. Discovery and distribution of Megacopta cribraria (Hemiptera: Heteroptera: Plataspidae) in northeast Georgia. J. Integr. Pest Manag. 2010, 1, F1–F4. [Google Scholar] [CrossRef]
- Liang, W.; Tran, L.; Washington-Allen, R.; Wiggins, G.; Stewart, S.; Vogt, J.; Grant, J. Predicting the potential invasion of kudzu bug, Megacopta cribraria (Heteroptera: Plataspidae), in North and South America and determining its climatic preference. Biol. Invasions 2018, 20, 2899–2913. [Google Scholar] [CrossRef]
- Eger, J.E., Jr.; Ames, L.M.; Suiter, D.R.; Jenkins, T.M.; Rider, D.A.; Halbert, S.E. Occurrence of the Old World bug Megacopta cribraria (Fabricius) (Heteroptera: Plataspidae) in Georgia: A serious home invader and potential legume pest. Insecta Mundi 2010, 0121, 1–11. [Google Scholar]
- Blount, J.L.; Buntin, G.D.; Sparks, A.N. Host preference of Megacopta cribraria (Hemiptera: Plataspidae) on selected edible beans and soybean. J. Econ. Entomol. 2015, 108, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hanula, J.L.; Horn, S. The biology and preliminary host range of Megacopta cribraria (Heteroptera: Plataspidae) and its impact on kudzu growth. Environ. Entomol. 2012, 41, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Seiter, N.J.; Greene, J.K.; Reay-Jones, F.P.F. Reduction of soybean yield components by Megacopta cribraria (Hemiptera: Plataspidae). J. Econ. Entomol. 2013, 106, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Medal, J.; Cruz, A.S.; Williams, K.; Fraser, S.; Wolaver, D.; Smith, T.; Davis, B.J. First Record of Paratelenomus saccharalis (Hymenoptera: Platygastridae) on Kudzu Bug Megacopta cribraria (Heteroptera: Plataspidae) in Florida. Fla. Entomol. 2015, 98, 1250–1251. [Google Scholar] [CrossRef]
- Tillman, G.; Gaskin, J.; Endale, D.; Johnson, C.; Schomberg, H. Parasitism of Megacopta cribraria (Hemiptera: Plataspidae) by Paratelenomus saccharalis (Hymenoptera: Platygastridae) in Organic Soybean Plots in Georgia, USA. Fla. Entomol. 2016, 99, 300–302. [Google Scholar] [CrossRef]
- Knight, I.A.; Roberts, P.M.; Gardner, W.A.; Oliver, K.M.; Reay-Jones, F.P.F.; Reisig, D.D.; Toews, M.D. Spatial distribution of Megacopta cribraria (Hemiptera: Plataspidae) adults, eggs and parasitism by Paratelenomus saccharalis (Hymenoptera: Platygastridae) in soybean. Environ. Entomol. 2017, 46, 1292–1298. [Google Scholar] [CrossRef]
- Xu, Z.; Cui, J.; Bi, R.; Shi, S.; Gao, Y. Effect of photoperiod on development and reproduction of Megacopta cribraria (Hemiptera: Plataspidae). Oil Crop. Sci. 2019, 4, 90–99. [Google Scholar]
- Hodek, I. Termination of adult diapause in Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae) in the field. Entomol. Exp. Appl. 1971, 14, 212–222. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.2-13. 2015. Available online: https://cran.r-project.org/package=survival (accessed on 28 November 2021).
- Searle, S.R.; Speed, F.M.; Milliken, G.A. Population marginal means in the linear model: An alternative to least squares means. Am. Stat. 1980, 34, 216–221. [Google Scholar] [CrossRef]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.5.3. 2020. Available online: https://cran.r-project.org/package=emmeans (accessed on 28 November 2021).
- Shi, S.S.; Cui, J.; Zang, L.S. Development, survival, and reproduction of Megacopta cribraria (Heteroptera: Plataspidae) at different constant temperatures. J. Econ. Entomol. 2014, 107, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.E.; de Bortoli, S.A.; Takahashi, R. Development and silk production by silkworm larvae after topical application of methoprene. Sci. Agric. 2002, 59, 585–588. [Google Scholar] [CrossRef] [Green Version]
- Murakoshi, S.; Chang, C.-F.; Tamura, S. Increase in silk production by the silkworm, Bombyx mori L., due to oral administration of a juvenile hormone analog. Agric. Biol. Chem. 1972, 36, 695–696. [Google Scholar] [CrossRef]
- Lasa, R.; Caballero, P.; Williams, T. Juvenile hormone analogs greatly increase the production of a nucleopolyhedrovirus. Biol. Control 2007, 41, 389–396. [Google Scholar] [CrossRef]
- Hayes, D.K.; Sullivan, W.N.; Oliver, M.Z.; Schechter, M.S. Photoperiod manipulation of insect diapause: A method of pest control? Science 1970, 169, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.-P.; Iwema, T.; Epa, V.C.; Takaki, K.; Rynes, J.; Jindra, M. Ligand-binding properties of a juvenile hormone receptor, methoprene-tolerant. Proc. Natl. Acad. Sci. USA 2011, 108, 21128–21133. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
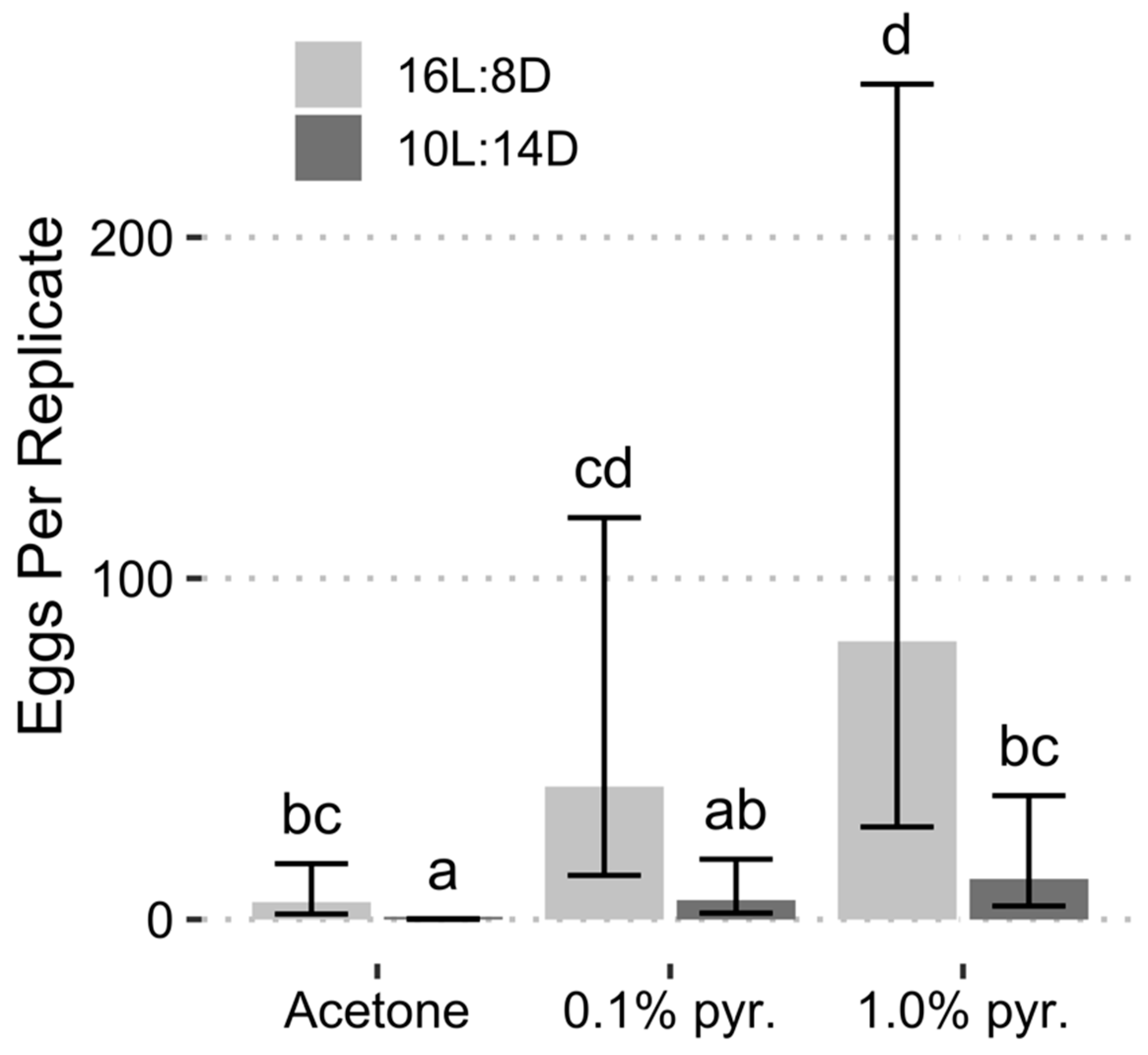{kind=link}
{kind=link}
{kind=link}
| Collection | Factor | Coefficient | SE | Z | p |
|---|---|---|---|---|---|
| August | Intercept | 3.375 | 0.099 | 33.92 | <0.0001 |
| 1.0% pyr. | −0.546 | 0.117 | −4.68 | <0.0001 | |
| 0.1% pyr. | −0.045 | 0.123 | −0.37 | 0.7130 | |
| Long-day | 0.154 | 0.096 | −1.61 | 0.1072 | |
| Log(scale) | −0.1620 | 0.049 | −3.33 | <0.0001 | |
| September | Intercept | 4.307 | 0.050 | 86.07 | <0.0001 |
| 1.0% pyr. | −0.621 | 0.059 | −10.48 | <0.0001 | |
| 0.1% pyr. | −0.226 | 0.059 | −3.85 | 0.0001 | |
| Long-day | −0.273 | 0.046 | −5.92 | <0.0001 | |
| Log(scale) | −0.815 | 0.042 | −19.35 | <0.0001 |
| Collection | Factor | Coefficient | SE | Z | p |
|---|---|---|---|---|---|
| August | Intercept | 1.619 | 0.599 | 2.70 | 0.0069 |
| 1.0% pyr. | 2.782 | 0.738 | 3.77 | 0.0002 | |
| 0.1% pyr. | 2.045 | 0.741 | 2.76 | 0.0058 | |
| Short-day | −1.921 | 0.588 | −3.27 | 0.0011 | |
| September | Intercept | 5.172 | 0.107 | 48.20 | <0.0001 |
| 1.0% pyr. | 1.299 | 0.140 | 9.31 | <0.0001 | |
| 0.1% pyr. | 0.841 | 0.140 | 6.01 | <0.0001 | |
| Short-day | −0.666 | 0.104 | −6.396 | <0.0001 |
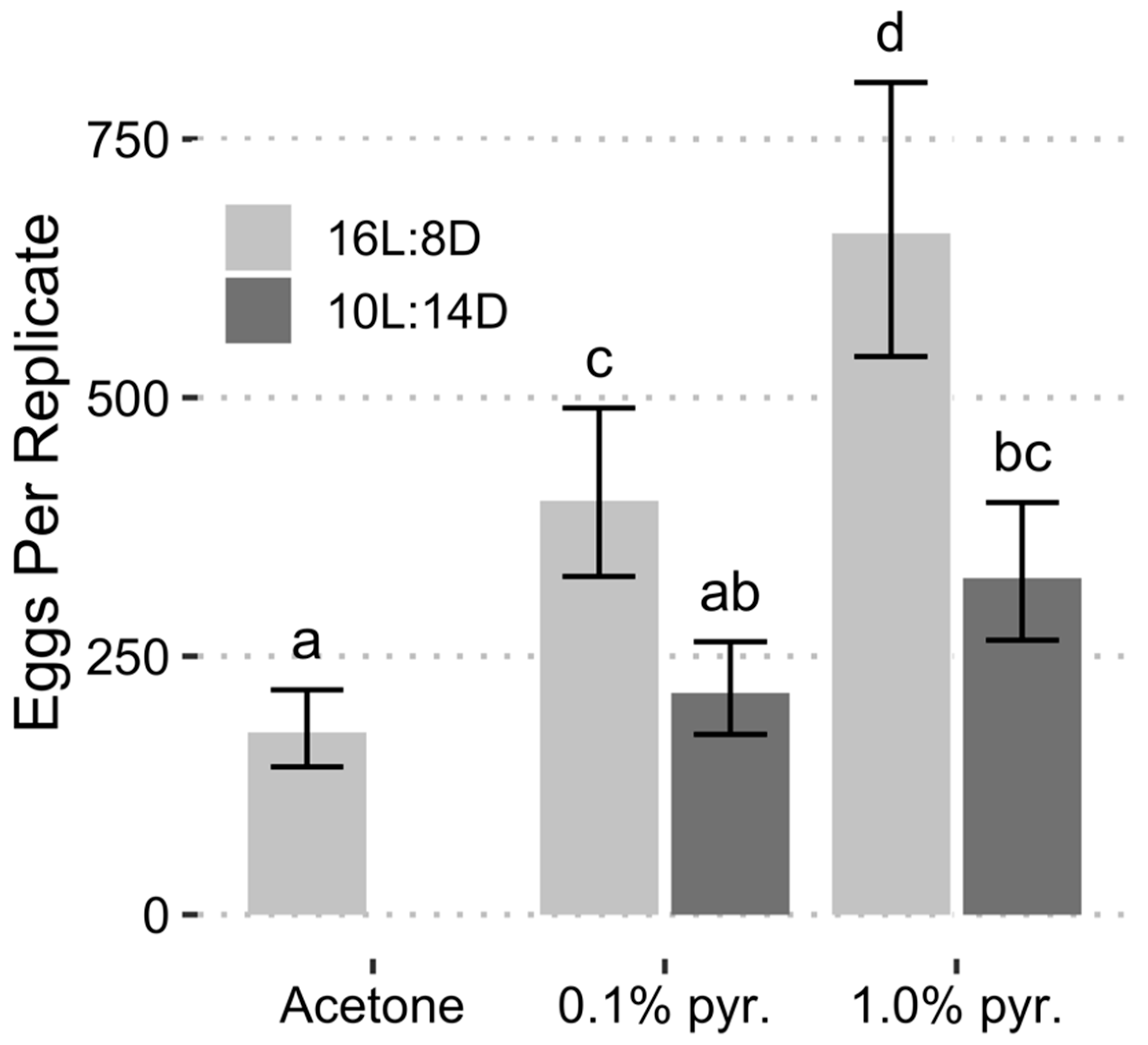
| Treatment | Megacopta cribraria1 | Paratelenomus saccharalis | |||
|---|---|---|---|---|---|
| Chemical | Photoperiod | Eggs | % Eclosion ± SE | Eggs | % Emergence ± SE |
| Acetone | 10L:14D | - | - | - | - |
| Acetone | 16L:8D | 400 | 28.06 ± 1.91 a | 128 | 34.69 ± 3.34 a |
| 0.1% pyr. | 10L:14D | 183 | 0.54 ± 0.54 b | 366 | 8.95 ± 1.42 c |
| 0.1% pyr. | 16L:8D | 665 | 2.21 ± 0.56 b | 321 | 28.82 ± 2.13 a |
| 1.0% pyr. | 10L:14D | 177 | 0.00 ± 0.00 b | 384 | 10.90 ± 1.50 c |
| 1.0% pyr. | 16L:8D | 1065 | 1.48 ± 0.37 b | 414 | 17.36 ± 1.69 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penca, C.; Goltz, N.C.; Hodges, A.C.; Leppla, N.C.; Eger, J.E.; Smith, T.R. Use of Pyriproxyfen to Induce Oogenesis in Diapausing Megacopta cribraria (Heteroptera: Plataspidae), and Evaluation of Pyriproxyfen-Induced Eggs for Rearing the Parasitoid Paratelenomus saccharalis (Hymenoptera: Scelionidae). Insects 2022, 13, 89. https://doi.org/10.3390/insects13010089
Penca C, Goltz NC, Hodges AC, Leppla NC, Eger JE, Smith TR. Use of Pyriproxyfen to Induce Oogenesis in Diapausing Megacopta cribraria (Heteroptera: Plataspidae), and Evaluation of Pyriproxyfen-Induced Eggs for Rearing the Parasitoid Paratelenomus saccharalis (Hymenoptera: Scelionidae). Insects. 2022; 13(1):89. https://doi.org/10.3390/insects13010089
Chicago/Turabian StylePenca, Cory, Nicholas C. Goltz, Amanda C. Hodges, Norman C. Leppla, Joseph E. Eger, and Trevor R. Smith. 2022. "Use of Pyriproxyfen to Induce Oogenesis in Diapausing Megacopta cribraria (Heteroptera: Plataspidae), and Evaluation of Pyriproxyfen-Induced Eggs for Rearing the Parasitoid Paratelenomus saccharalis (Hymenoptera: Scelionidae)" Insects 13, no. 1: 89. https://doi.org/10.3390/insects13010089
APA StylePenca, C., Goltz, N. C., Hodges, A. C., Leppla, N. C., Eger, J. E., & Smith, T. R. (2022). Use of Pyriproxyfen to Induce Oogenesis in Diapausing Megacopta cribraria (Heteroptera: Plataspidae), and Evaluation of Pyriproxyfen-Induced Eggs for Rearing the Parasitoid Paratelenomus saccharalis (Hymenoptera: Scelionidae). Insects, 13(1), 89. https://doi.org/10.3390/insects13010089