
Sampling Optimization and Crop Interface Effects on Lygus lineolaris Populations in Southeastern USA Cotton

 , , , , ,
, , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
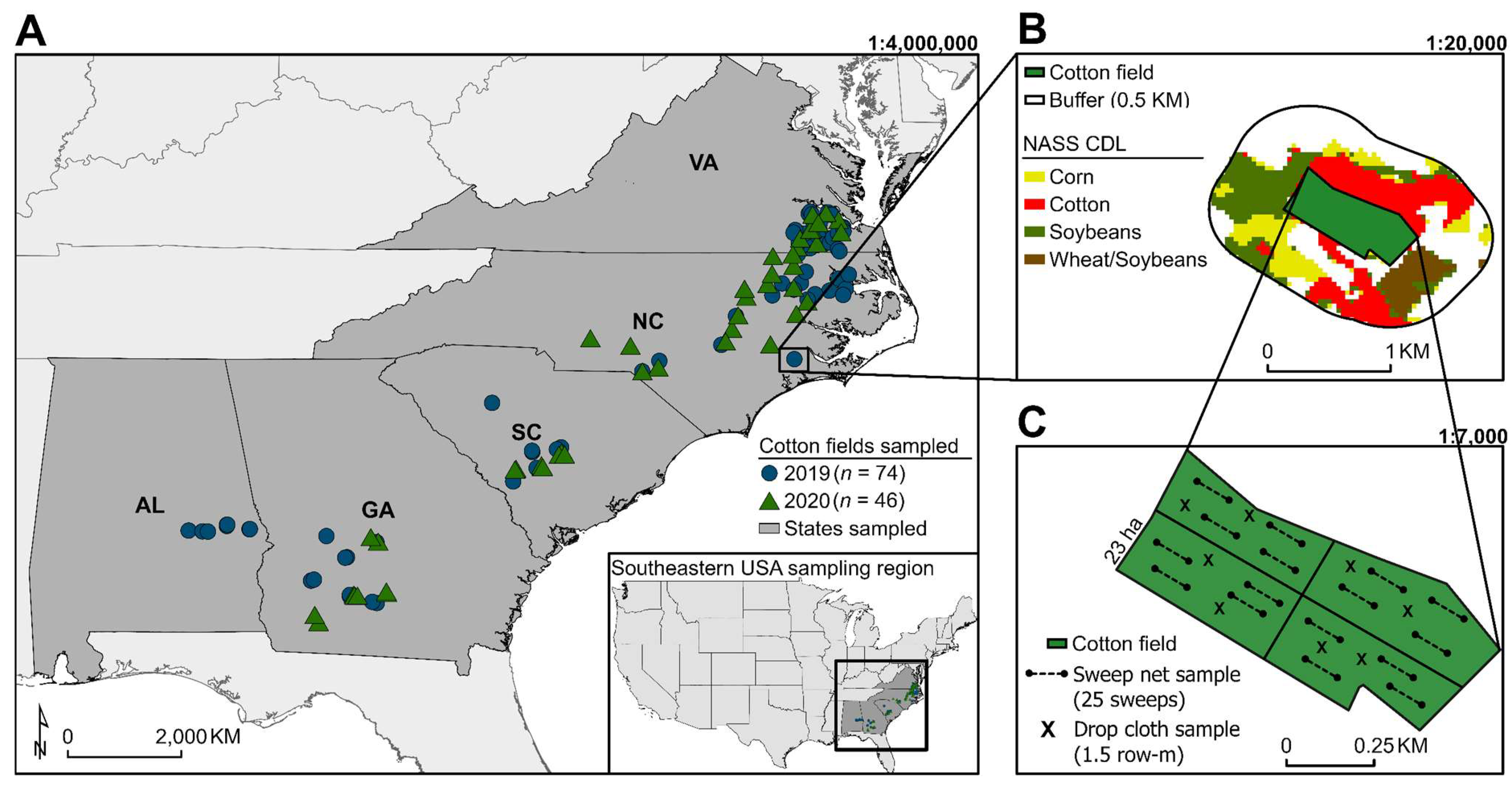
2.1. Field Selection and L. lineolaris Sampling
2.2. Plant Injury Assessments
2.3. Landscape Data Extraction
2.4. Statistical Analysis
2.4.1. Lygus lineolaris Abundance Variation and Fixed Sampling Plan
2.4.2. Correlating L. lineolaris Density to Plant Injury
2.4.3. Local Landscape Effects on L. lineolaris Abundance
3. Results
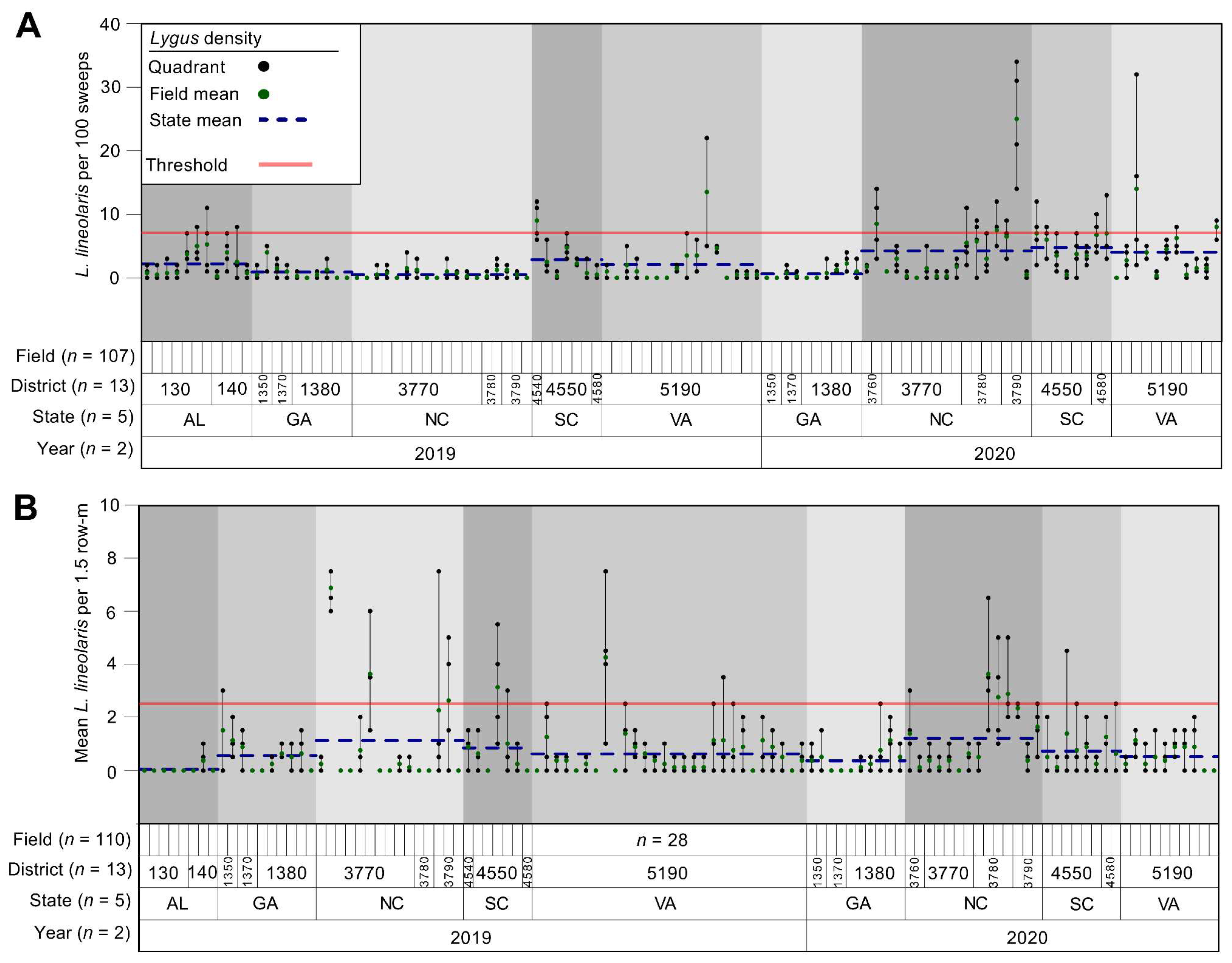
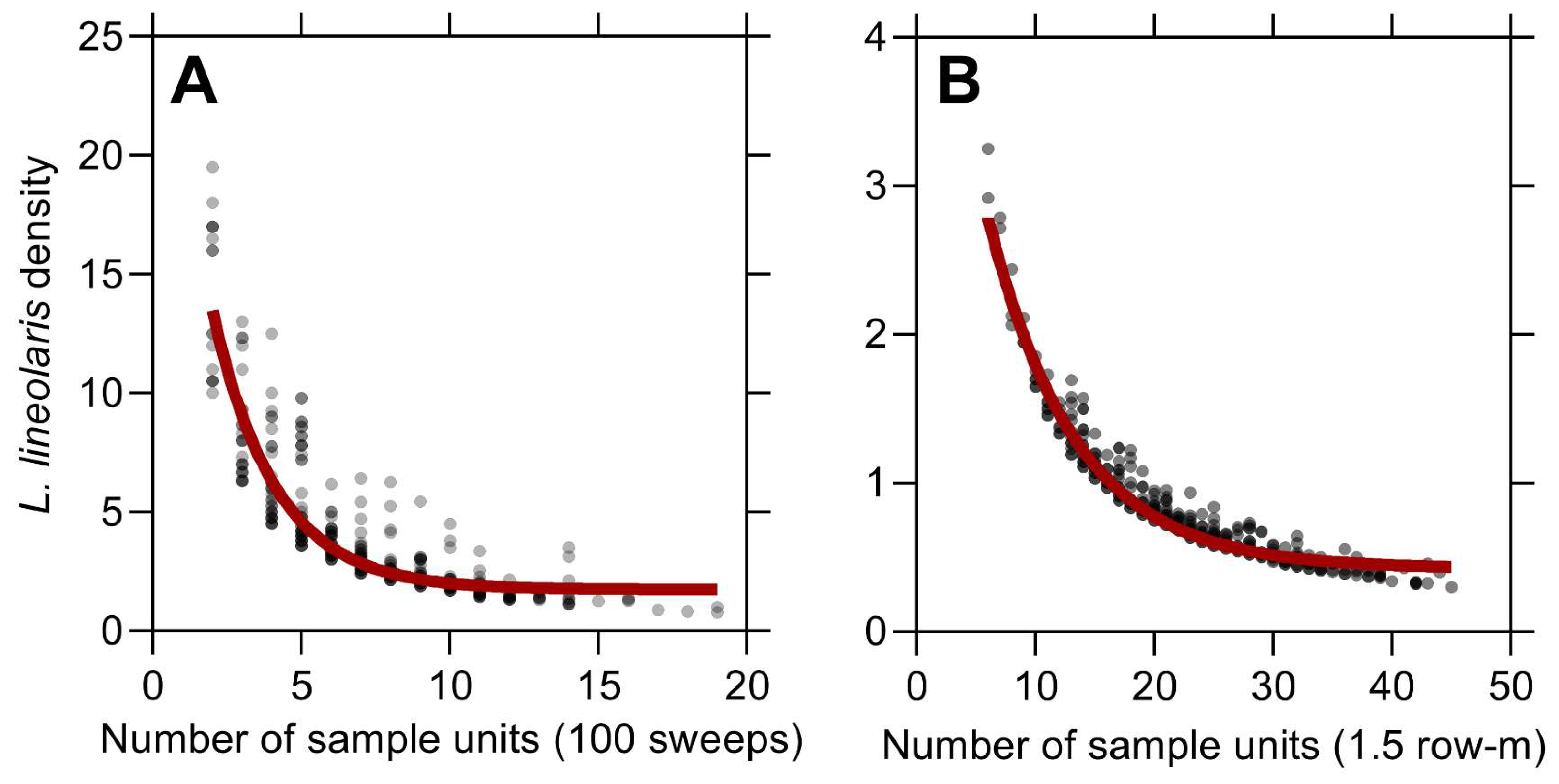
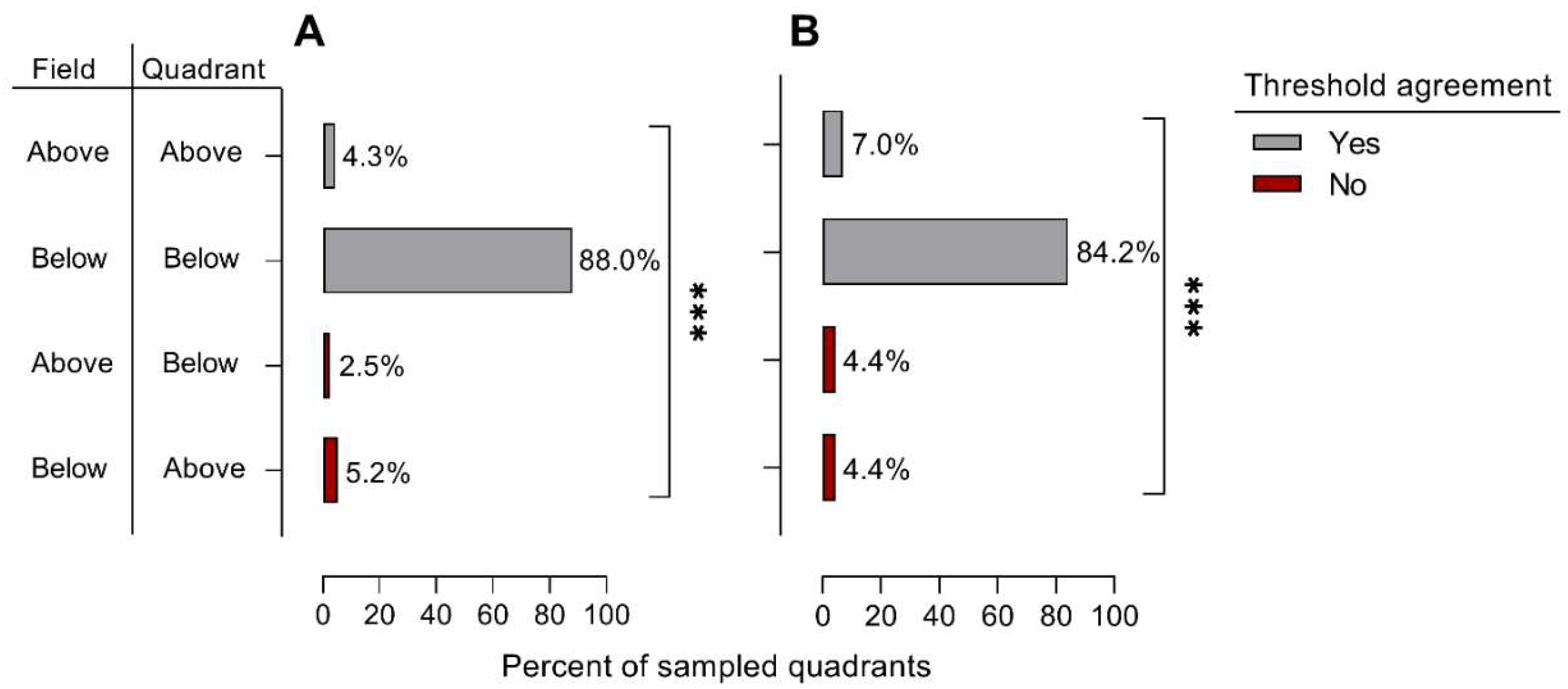
3.1. Lygus lineolaris Abundance Variation and Fixed Sampling Plan
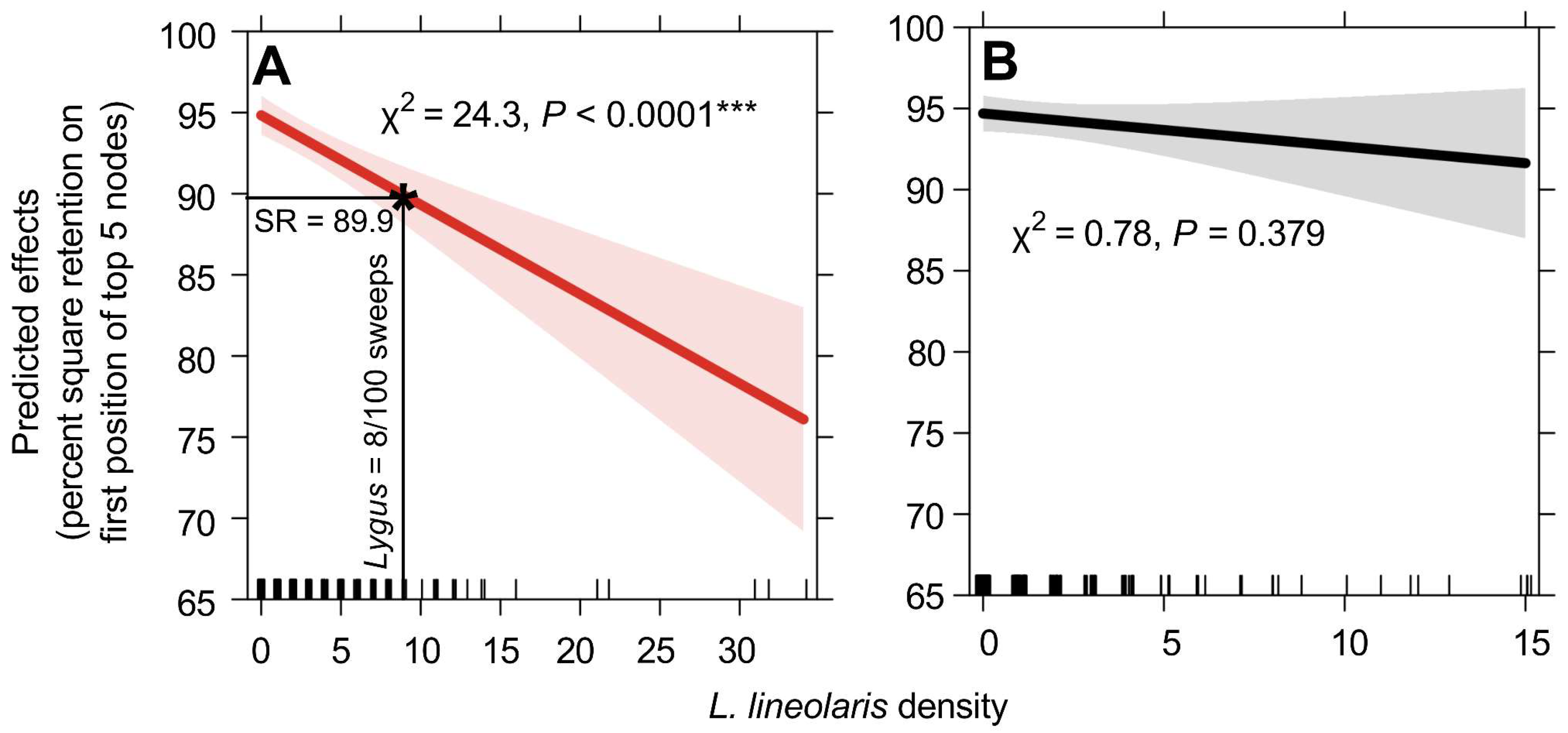
3.2. Correlating L. lineolaris Density to Plant Injury
3.3. Local Landscape Effects on L. lineolaris Abundance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.D.; Park, S.; Park, Y.S.; Chung, Y.J.; Lee, B.Y.; Chon, T.S. Range expansion of forest pest populations by using the lattice model. Ecol. Model. 2007, 203, 157–166. [Google Scholar] [CrossRef]
- Byers, J.A. Simulation of the mate-finding behaviour of pine shoot beetles, Tomicus piniperda. Anim. Behav. 1991, 41, 649–660. [Google Scholar] [CrossRef]
- Vinatier, F.; Tixier, P.; Duyck, P.F.; Lescourret, F. Factors and mechanisms explaining spatial heterogeneity: A review of methods for insect populations. Methods Ecol. Evol. 2011, 2, 11–22. [Google Scholar] [CrossRef]
- Pedigo, L.P. Introduction to sampling arthropod populations. In Arthropods in Agriculture; CRC Press: Boca Raton, FL, USA, 1994; pp. 1–11. [Google Scholar]
- Kogan, M. Integrated pest management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef]
- Taylor, L.R. Aggregation, variance and the mean. Nature 1961, 189, 732–735. [Google Scholar] [CrossRef]
- Perry, J.N. Spatial analysis by distance indices. J. Anim. Ecol. 1995, 64, 303–314. [Google Scholar] [CrossRef]
- Begon, M.; Harper, J.L.; Townsend, C.R. Ecology: Individuals, Populations and Communities, 3rd ed.; Blackwell Science: Oxford, UK, 1996. [Google Scholar]
- Elliott, J.M. Some Methods for the Statistical Analysis of Samples of Benthic Invertebrates, 2nd ed.; Freshwater Biological Association Scientific Publications: Cumbria, UK, 1977. [Google Scholar]
- Davis, P.M. Statistics for describing populations. In Handbook for Sampling Methods for Arthropods in Agriculture; CRC Press: Boca Raton, FL, USA, 1994; pp. 33–54. [Google Scholar]
- Fleischer, S.J.; Weisz, R.; Smilowitz, Z.; Midgarden, D. Spatial variation in insect populations and site-specific integrated pest management. In The State of Site-Specific Management for Agriculture; American Society of America, Inc.: Madison, WI, USA, 1997; pp. 101–130. [Google Scholar]
- Meehan, T.D.; Werling, B.P.; Landis, D.A.; Gratton, C. Agricultural landscape simplification and insecticide use in the Midwestern United States. Proc. Natl. Acad. Sci. USA 2011, 108, 11500–11505. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, M.E.; Rienzo-Stack, K.; Power, A.G. A multi-scale, landscape approach to predicting insect populations in agroecosystems. Ecol. Appl. 2011, 21, 1782–1791. [Google Scholar] [CrossRef]
- Huseth, A.S.; Frost, K.E.; Knuteson, D.L.; Wyman, J.A.; Groves, R.L. Effects of landscape composition and rotation distance on Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) abundance in cultivated potato. Environ. Entomol. 2013, 41, 1553–1564. [Google Scholar] [CrossRef]
- Stack-Whitney, K.; Meehan, T.D.; Kucharik, C.J.; Zhu, J.; Townsend, P.A.; Hamilton, K.; Gratton, C. Explicit modeling of abiotic and landscape factors reveals precipitation and forests associated with aphid abundance. Ecol. Appl. 2016, 26, 2598–2608. [Google Scholar] [CrossRef]
- Meisner, M.H.; Zaviezo, T.; Rosenheim, J.A. Landscape crop composition effects on cotton yield, Lygus hesperus densities and pesticide use. Pest Manag. Sci. 2017, 73, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Jeffris, R.A.; Nault, B.A. Crop spatiotemporal dominance is a better predictor of pest and predator abundance than traditional partial approaches. Agric. Ecosyst. Environ. 2018, 265, 331–339. [Google Scholar] [CrossRef]
- Dorman, S.J.; Schürch, R.; Huseth, A.S.; Taylor, S.V. Landscape and climatic effects driving spatiotemporal abundance of Lygus lineolaris (Hemiptera: Miridae) in cotton agroecosystems. Agric. Ecosyst. Environ. 2020, 295, 106910. [Google Scholar] [CrossRef]
- Cline, A.R.; Backus, E.A. Correlations among AC electronic monitoring waveforms, body postures, and stylet penetration behaviors of Lygus hesperus (Hemiptera: Miridae). Environ. Entomol. 2002, 31, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Shackel, K.A.; de la Paz Celorio-Mancera, M.; Ahmadi, H.; Greve, L.C.; Teuber, L.R.; Backus, E.A.; Labavitch, J.M. Micro-injection of lygus salivary gland proteins to simulate feeding damage in alfalfa and cotton flowers. Arch. Insect Biochem. Physiol. 2005, 58, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.L. Heteroptera as vectors of plant pathogens. Neotrop. Entomol. 2004, 33, 519–545. [Google Scholar] [CrossRef] [Green Version]
- Snodgrass, G.L.; Scott, W.P.; Smith, J.W. Host plants and seasonal distribution of the tarnished plant bug (Hemiptera: Miridae) in the Delta of Arkansas, Louisiana, and Mississippi. Environ. Entomol. 1984, 13, 110–116. [Google Scholar] [CrossRef]
- Young, O.P. Host plants of the tarnished plant bug, Lygus lineolaris (Heteroptera: Miridae). Ann. Entomol. Soc. Am. 1986, 79, 747–762. [Google Scholar] [CrossRef]
- Jackson, R.E.; Allen, K.C.; Snodgrass, G.L.; Krutz, L.J.; Gore, J.; Perera, O.P.; Price, L.D.; Mullen, R.M. Influence of maize and pigweed on tarnished plant bug (Hemiptera: Miridae) populations infesting cotton. Southwest. Entomol. 2014, 39, 391–399. [Google Scholar]
- Carrière, Y.; Ellsworth, P.C.; Dutilleul, P.; Ellers-Kirk, C.; Barkley, V.; Antilla, L. A GIS-based approach for areawide pest management: The scales of Lygus hesperus movements to cotton from alfalfa, weeds, and cotton. Entomol. Exp. Appl. 2006, 118, 203–210. [Google Scholar] [CrossRef]
- Goethe, J.K.; Dorman, S.J.; Huseth, A.S. Local and landscape scale drivers of Euschistus servus and Lygus lineolaris in North Carolina small grain agroecosystems. Agric. For. Entomol. 2021, 23, 441–451. [Google Scholar] [CrossRef]
- D’Ambrosio, D.A.; Peele, W.; Hubers, A.; Huseth, A.S. Seasonal dispersal of Lygus lineolaris (Hemiptera: Miridae) from weedy hosts into differently fragmented cotton landscapes in North Carolina. Crop Prot. 2019, 125, 104898. [Google Scholar] [CrossRef]
- Musser, F.; Stewart, S.; Bagwell, R.; Lorenz, G.; Catchot, A.; Burris, E.; Cook, D.; Robbins, J.; Greene, J.; Studebaker, G.; et al. Comparison of direct and indirect sampling methods for tarnished plant bug (Hemiptera: Miridae) in flowering cotton. J. Econ. Entomol. 2007, 100, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- Musser, F.R.; Lorenz, G.M.; Stewart, S.D.; Bagwell, R.D.; Leonard, B.R.; Catchot, A.L.; Tindall, K.V.; Studebaker, G.E.; Akin, D.S.; Cook, D.R.; et al. Tarnished plant bug (Hemiptera: Miridae) thresholds for cotton before bloom in the midsouth of the United States. J. Econ. Entomol. 2009, 102, 2109–2115. [Google Scholar] [CrossRef] [PubMed]
- Musser, F.; Catchot, A.L.; Stewart, S.D.; Bagwell, R.D.; Lorenz, G.M.; Tindall, K.V.; Studebaker, G.E.; Leonard, B.R.; Akin, D.S.; Cook, D.R.; et al. Tarnished plant bug (Hemiptera: Miridae) thresholds and sampling comparisons for flowering cotton in the midsouthern United States. J. Econ. Entomol. 2009, 102, 1827–1836. [Google Scholar] [CrossRef] [PubMed]
- Gore, J.; Catchot, A.; Musser, F.; Greene, J.; Leonard, B.R.; Cook, D.R.; Snodgrass, G.L.; Jackson, R. Development of a plant-based threshold for tarnished plant bug (Hemiptera: Miridae) in cotton. J. Econ. Entomol. 2012, 105, 2007–2014. [Google Scholar] [CrossRef] [Green Version]
- Aghaee, M.A.; Dorman, S.J.; Taylor, S.V.; Reisig, D.D. Evaluating optimal spray timing, planting date, and current thresholds for Lygus lineolaris (Hemiptera: Miridae) in Virginia and North Carolina cotton. J. Econ. Entomol. 2019, 112, 1207–1216. [Google Scholar] [CrossRef]
- Snodgrass, G.L. Estimating absolute density of nymphs of Lygus lineolaris (Heteroptera: Miridae) in cotton using drop cloth and sweep-net sampling methods. J. Econ. Entomol. 1993, 86, 1116–1123. [Google Scholar] [CrossRef]
- Dorman, S.J.; Reisig, D.D.; Malone, S.; Taylor, S.V. Systems approach to evaluate tarnished plant bug (Hemiptera: Miridae) management practices in Virginia and North Carolina cotton. J. Econ. Entomol. 2020, 113, 2223–2234. [Google Scholar] [CrossRef]
- Rosenheim, J.A. Evaluating the quality of ecoinformatics data derived from commercial agriculture: A repeatability analysis of pest density estimates. J. Econ. Entomol. 2021, 114, 1842–1846. [Google Scholar] [CrossRef]
- Stewart, S.D.; Gaylor, M.J. Effects of age, sex, and reproductive status on flight by the tarnished plant bug (Heteroptera: Miridae). Popul. Ecol. 1994, 23, 80–84. [Google Scholar] [CrossRef]
- Outward, R.; Sorenson, C.E.; Bradley, J.R. Effects of vegetated field borders on arthropods in cotton fields in eastern North Carolina. J. Insect Sci. 2008, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.N.; McCarty, J.C., Jr.; Parott, W.L. Fruiting efficiency in cotton: Bolls size and boll set percentage. Crop Sci. 1990, 30, 857–860. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA) National Agricultural Statistics Service (NASS). Cropland Data Layer (CDL). Published Crop-Specific Data Layer [Online]. USDA-NASS: Washington, DC, USA; 2020. Available online: https://nassgeodata.gmu.edu/CropScape/ (accessed on 1 July 2021).
- Schuetzenmeister, A.; Dufey, F. VCA: Variance Component Analysis. 2020. Available online: https://cran.r-project.org/package=VCA (accessed on 1 July 2021).
- Feltz, C.J.; Miller, G.E. An asymptotic test for the equality of coefficients of variation from k populations. Stat. Med. 1996, 15, 647–658. [Google Scholar] [CrossRef]
- Green, R.H. On fixed precision sequential sampling. Res. Popul. Ecol. 1970, 12, 249–251. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Ecological Methods, with Particular Reference to the Study of Insect Populations; Chapman & Hall: London, UK, 1978. [Google Scholar]
- Hutchison, W.D. Sequential Sampling to Determine Population Density. In Handbook of Sampling Methods for Arthropods in Agriculture, 1st ed.; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Naranjo, S.E.; Hutchison, W.D. Validation of arthropod sampling plans using a resampling approach: Software and analysis. Am. Entomol. 1997, 43, 48–57. [Google Scholar] [CrossRef]
- Bates, D.M.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian inference for latent Gaussian models by using integrated nested Laplace approximations. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2009, 71, 319–392. [Google Scholar] [CrossRef]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; Van Der Linde, A. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2002, 64, 583–639. [Google Scholar] [CrossRef] [Green Version]
- Croissant, Y.; Millo, G. Panel data econometrics in R: The plm package. J. Stat. Softw. 2008, 27, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Sevacherian, V.; Stern, V.M. Movements of Lygus bugs between alfalfa and cotton. Environ. Entomol. 1974, 4, 163–165. [Google Scholar] [CrossRef]
- Brewer, M.J.; Anderson, D.J.; Armstrong, J.S.; Villanueva, R.T. Sampling strategies for square and boll-feeding plant bugs (Hemiptera: Miridae) occurring on cotton. J. Econ. Entomol. 2012, 105, 896–905. [Google Scholar] [CrossRef]
- Reay-Jones, F.P.F. Spatial and temporal patterns of stink bugs (Hemiptera: Pentatomidae) in wheat. Environ. Entomol. 2010, 39, 944–955. [Google Scholar] [CrossRef]
- Pulakkatu-Thodi, I.; Reisig, D.D.; Greene, J.K.; Reay-Jones, F.P.F.; Toews, M.D. Within-field spatial distribution of stink bug (Hemiptera: Pentatomidae)-induced boll injury in commercial cotton fields of the southeastern United States. Environ. Entomol. 2014, 43, 744–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, A.; Reisig, D.D. Developing a sampling plan for brown stink bug (Hemiptera: Pentatomidae) in field corn. J. Econ. Entomol. 2018, 111, 1915–1926. [Google Scholar] [CrossRef]
- Pezzini, D.T.; DiFonzo, C.D.; Finke, D.L.; Hunt, T.E.; Knodel, J.J.; Krupke, C.H.; McCornack, B.; Michel, A.P.; Moon, A.D.; Philips, C.R.; et al. Spatial patterns and sequential sampling plans for estimating densities of stink bugs (Hemiptera: Pentatomidae) in soybean in the north central region of the United States. J. Econ. Entomol. 2019, 112, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Bachman, P.M.; Ahmad, A.; Ahrens, J.E.; Akbar, W.; Baum, J.A.; Brown, S.; Clark, T.L.; Fridley, J.M.; Gowda, A.; Greenplate, J.T.; et al. Characterization of the activity spectrum of MON 88702 and the plant-incorporated protectant Cry51Aa2. PLoS ONE 2017, 12, e0169409. [Google Scholar] [CrossRef] [Green Version]
- Akbar, W.; Gowda, A.; Ahrens, J.E.; Stelzer, J.W.; Brown, R.S.; Bollman, S.L.; Greenplate, J.T.; Gore, J.; Catchot, A.L.; Lorenz, G..; et al. First transgenic trait for control of plant bugs and thrips in cotton. Pest Manag. Sci. 2019, 75, 867–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbin, J.C.; Towles, T.B.; Crow, W.D.; Catchot, A.L.; Cook, D.R.; Dodds, D.M.; Gore, J. Evaluation of Current Tarnished Plant Bug (Hemiptera: Miridae) Thresholds in Transgenic MON 88702 Cotton Expressing the Bt Cry51Aa2. 834_16 Trait. J. Econ. Entomol. 2020, 113, 1816–1822. [Google Scholar] [CrossRef]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martínez-Salinas, A.; et al. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [Green Version]
- Rosenheim, J.A.; Gratton, C. Ecoinformatics (big data) for agricultural entomology: Pitfalls, progress, and promise. Annu. Rev. Entomol. 2017, 62, 399–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Root, R.B. Organization of a plant arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Allen, K.C.; Luttrell, R.G. Spatial and temporal distribution of heliothines and tarnished plant bugs across the landscape of an Arkansas farm. Crop Prot. 2009, 28, 722–727. [Google Scholar] [CrossRef]
- Carrière, Y.; Goodell, P.B.; Ellers-Kirk, C.; Larocque, G.; Dutilleul, P.; Naranjo, S.E.; Ellsworth, P.C. Effects of local and landscape factors on population dynamics of a cotton pest. PLoS ONE 2012, 7, e39862. [Google Scholar] [CrossRef] [Green Version]
- Fleischer, S.J.; Gaylor, M.J. Lygus lineolaris (Heteroptera: Miridae) population dynamics: Nymphal development, life tables, and Leslie matrices on selected weeds and cotton. Environ. Entomol. 1988, 17, 246–253. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technique | Source | df | VC a | % Variation b | CV c |
|---|---|---|---|---|---|
| Sweep net (100 sweeps) | Year | 1 | 1.94 | 9.01 | 0.48 |
| State | - | 0 | - | - | |
| District | 8 | 2.40 | 11.1 | 0.53 | |
| Field | 94 | 9.58 | 44.5 | 1.06 | |
| Field:quadrant | 292 | 6.85 | 31.8 | 0.90 | |
| Error | 3 | 0.79 | 3.67 | 0.30 | |
| Drop cloth (1.5 row-m) | Year | 1 | 314.1 | 1.15 | 34.6 |
| State | 4 | 2725.3 | 9.98 | 10.2 | |
| District | 7 | 1846.3 | 6.76 | 84.0 | |
| Field | 66 | 12,310.8 | 45.1 | 216.8 | |
| Field:quadrant | 236 | 10,098.9 | 37.0 | 196.4 | |
| Error | - | 0 | - | - |
| Covariate | Posterior Mean | 95% CI | Significant |
|---|---|---|---|
| Intercept | −1.08 | [−2.09, −0.09] | Yes |
| Agriculture | 5.32 | [1.25, 9.38] | Yes |
| Corn | −2.12 | [−6.61, 2.27] | No |
| Cotton | −2.28 | [−4.61, −0.10] | Yes |
| Forest | 0.74 | [−1.23, 2.73] | No |
| Soybeans | −1.47 | [−6.45, 3.35] | No |
| Double-crop wheat/soybeans | 10.9 | [1.59, 19.7] | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorman, S.J.; Taylor, S.V.; Malone, S.; Roberts, P.M.; Greene, J.K.; Reisig, D.D.; Smith, R.H.; Jacobson, A.L.; Reay-Jones, F.P.F.; Paula-Moraes, S.; et al. Sampling Optimization and Crop Interface Effects on Lygus lineolaris Populations in Southeastern USA Cotton. Insects 2022, 13, 88. https://doi.org/10.3390/insects13010088
Dorman SJ, Taylor SV, Malone S, Roberts PM, Greene JK, Reisig DD, Smith RH, Jacobson AL, Reay-Jones FPF, Paula-Moraes S, et al. Sampling Optimization and Crop Interface Effects on Lygus lineolaris Populations in Southeastern USA Cotton. Insects. 2022; 13(1):88. https://doi.org/10.3390/insects13010088
Chicago/Turabian StyleDorman, Seth J., Sally V. Taylor, Sean Malone, Phillip M. Roberts, Jeremy K. Greene, Dominic D. Reisig, Ronald H. Smith, Alana L. Jacobson, Francis P. F. Reay-Jones, Silvana Paula-Moraes, and et al. 2022. "Sampling Optimization and Crop Interface Effects on Lygus lineolaris Populations in Southeastern USA Cotton" Insects 13, no. 1: 88. https://doi.org/10.3390/insects13010088
APA StyleDorman, S. J., Taylor, S. V., Malone, S., Roberts, P. M., Greene, J. K., Reisig, D. D., Smith, R. H., Jacobson, A. L., Reay-Jones, F. P. F., Paula-Moraes, S., & Huseth, A. S. (2022). Sampling Optimization and Crop Interface Effects on Lygus lineolaris Populations in Southeastern USA Cotton. Insects, 13(1), 88. https://doi.org/10.3390/insects13010088