
How Did Seal Lice Turn into the Only Truly Marine Insects?



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
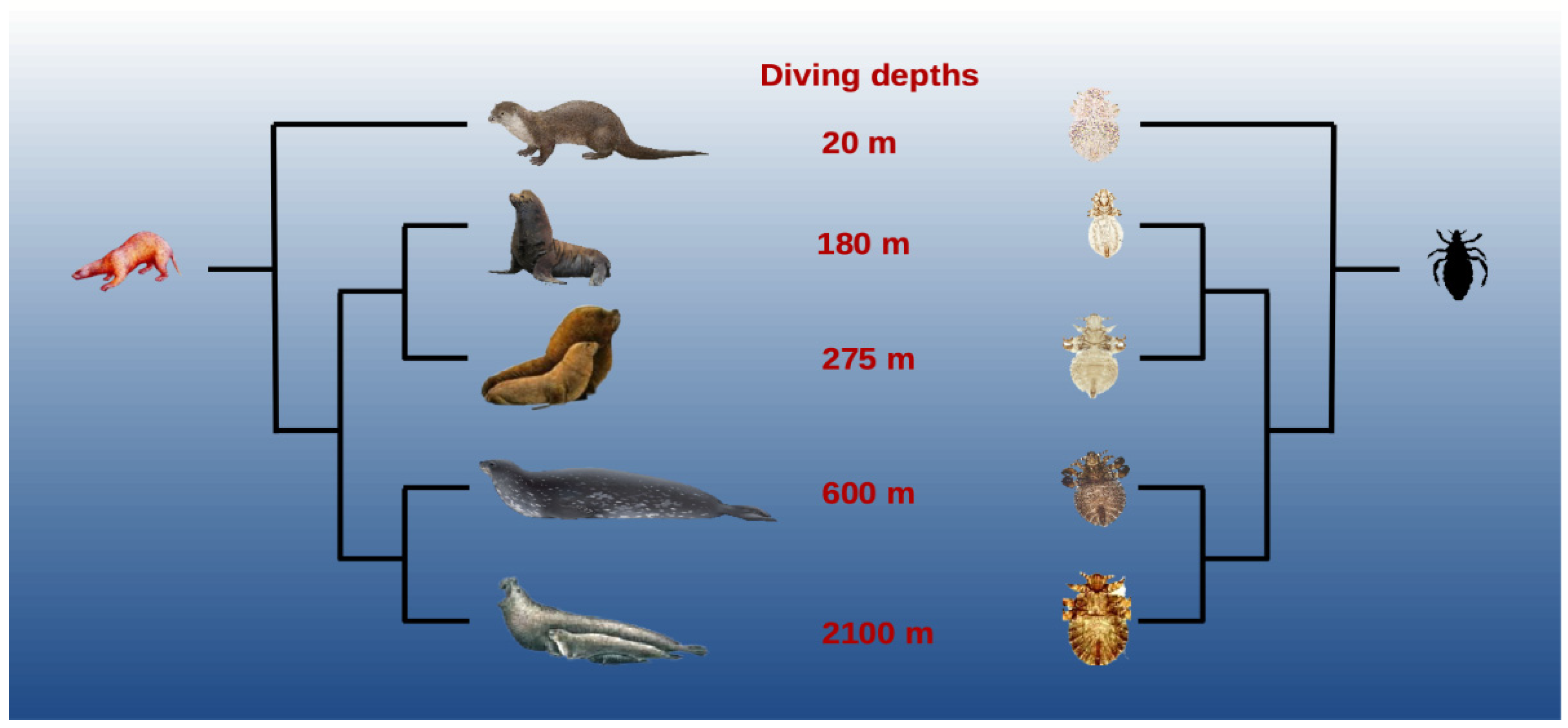
2.1. Evolution
2.2. Morphological Adaptations
2.3. Reproductive Synchronization with Hosts
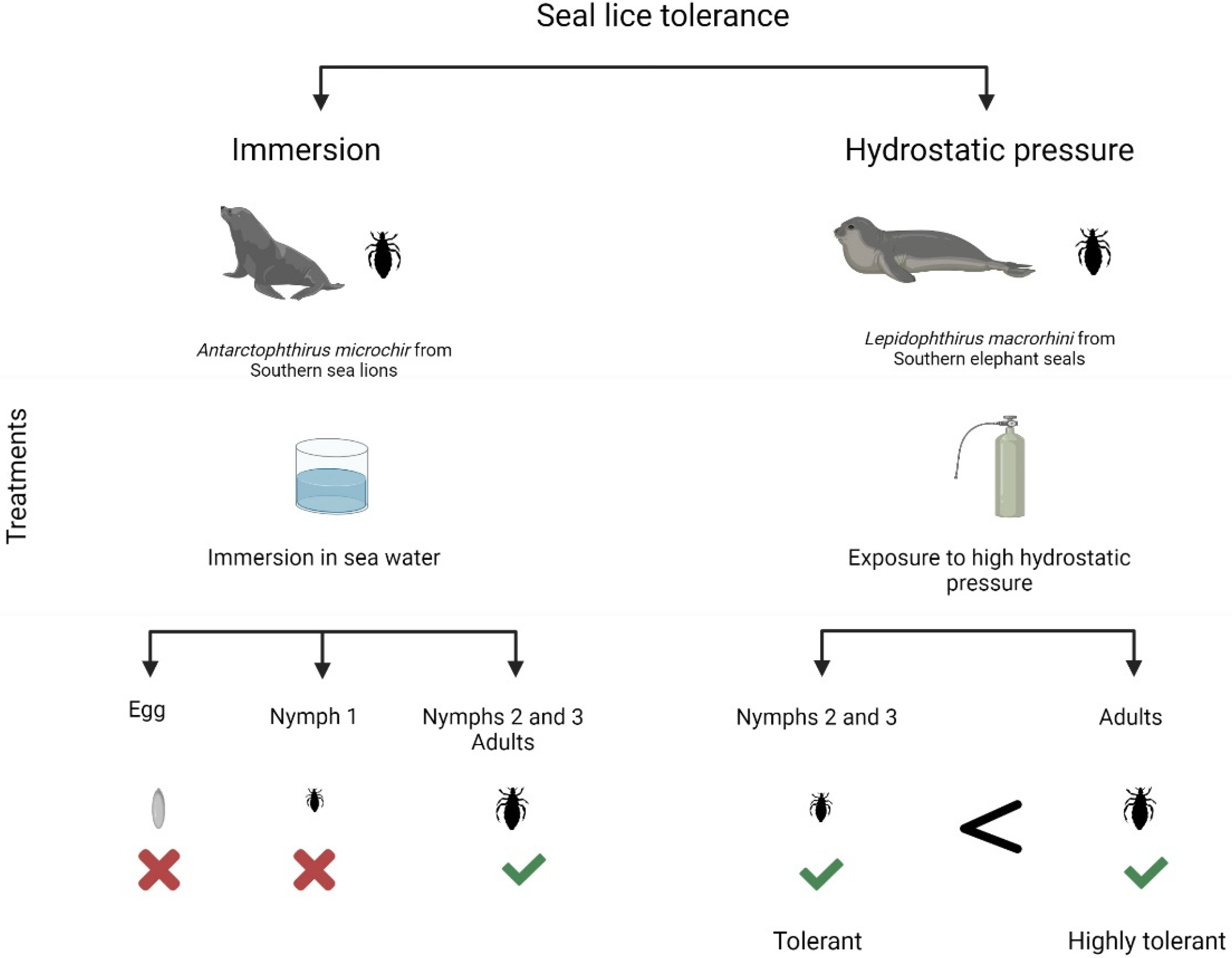
2.4. Tolerance to Immersion
2.5. Tolerance to Hydrostatic Pressure
2.6. Ecology
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Spence, J.R.; Anderson, N.M. Biology of water striders: Interactions between systematics and ecology. Annu. Rev. Entomol. 1994, 39, 101–128. [Google Scholar] [CrossRef]
- Ikawa, T.; Okabe, H.; Cheng, L. Skaters of the seas–comparative ecology of nearshore and pelagic Halobates species (Hemiptera: Gerridae), with special reference to Japanese species. Mar. Biol. Res. 2012, 8, 915–936. [Google Scholar] [CrossRef] [Green Version]
- Ruxton, G.D.; Humphries, S. Can ecological and evolutionary arguments solve the riddle of the missing marine insects? Mar. Ecol. 2008, 29, 72–75. [Google Scholar] [CrossRef]
- Kim, K.C. Coevolution of Parasitic Arthropods and Mammals; Wiley: New York, NY, USA, 1985. [Google Scholar]
- Bush, A.O.; Fernández, J.C.; Esch, G.W.; Seed, J.R. Parasitism: The Diversity and Ecology of Animal Parasites; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Durden, L.A.; Musser, G.G. The sucking lice (Insecta, Anoplura) of the world: A taxonomic checklist with records of mammalian hosts and geographical distributions. Bull. Am. Mus. Nat. 1994, 218, 1–90. [Google Scholar]
- Leonardi, M.S.; Palma, R.L. Review of the systematics, biology and ecology of lice from pinnipeds and river otters (Insecta: Phthiraptera: Anoplura: Echinophthiriidae). Zootaxa 2013, 3630, 445–466. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B. Diving behavior. In Encyclopedia of Marine Mammals; Perrin, W.F., Würsig, B., Thewissen, J.G.M., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 321–327. [Google Scholar]
- McIntyre, T.; de Bruyn, P.J.N.; Ansorge, I.J.; Bester, M.N.; Bornemann, H.; Plötz, J.; Tosh, C.A. A lifetime at depth: Vertical distribution of Southern elephant seals in the water column. Polar Biol. 2010, 33, 1037–1048. [Google Scholar] [CrossRef]
- Teilmann, J.; Born, E.W.; Aquarone, M. Behaviour of ringed seals tagged with satellite transmitters in the North Water polynya. Can. J. Zool. 1999, 77, 1934–1946. [Google Scholar] [CrossRef]
- Leonardi, M.S.; Virrueta Herrera, S.; Sweet, A.; Negrete, J.; Johnson, K.P. Phylogenomic analysis of seal lice reveals codivergence with their hosts. Syst. Entomol. 2019, 44, 699–708. [Google Scholar] [CrossRef]
- Leonardi, M.S.; Soto, F.; Negrete, J. Lousy big guys: Lepidophthirus macrorhini infesting seals from Antarctica. Polar Biol. 2018, 41, 481–485. [Google Scholar] [CrossRef]
- Hassanin, A.; Veron, G.; Ropiquet, A.; Jansen van Vuuren, B.; Lécu, A.; Goodman, S.M.; Haider, J.; Nguyen, T.T. Evolutionary history of Carnivora (Mammalia, Laurasiatheria) inferred from mitochondrial genomes. PLoS ONE 2021, 16, e0240770. [Google Scholar]
- Arnason, U.; Gullberg, A.; Janke, A.; Kullberg, M.; Lehman, N.; Petrov, E.A.; Väinölä, R. Pinniped phylogeny and a new hypothesis for their origin and dispersal. Mol. Phylogenet. Evol. 2006, 41, 345–354. [Google Scholar] [CrossRef]
- Aznar, F.J.; Balbuena, J.A.; Fernández, M.; Raga, J.A. Living Together: The parasites of marine mammals. In Marine Mammals; Evans, P.G.H., Raga, J.A., Eds.; Springer: Boston, MA, USA, 2002. [Google Scholar]
- Ménier, K. Origin and evolution of parasites in marine mammals: Pinnipedia as an example. Rev. Med. Vet. 2000, 151, 275–280. [Google Scholar]
- Leonardi, M.S. Clave para el reconocimiento de las especies de Echinophthiriidae (Phthiraptera: Anoplura) de Argentina y Antártida. Rev. Argent. Parasitol. 2014, 3, 24–30. [Google Scholar]
- Kim, K.C. Evolutionary parallelism in Anoplura and eutherian mammals. In Biosystematics of Haematophagous Insects, Systematics Association; Service, M.W., Ed.; Clarendon Press: Oxford, UK, 1988; pp. 91–114. [Google Scholar]
- Mehlhorn, B.; Mehlhorn, H.; Plötz, J. Light and scanning electron microscopical study on Antarctophthirus ogmorhini lice from the Antarctic seal Leptonychotes Weddellii. Parasitol. Res. 2002, 88, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.S.; Crespo, E.A.; Raga, J.A.; Fernández, M. Scanning electron microscopy of Antarctophthirus microchir (Phthiraptera: Anoplura: Echinophthiriidae): Studying morphological adaptations to aquatic life. Micron 2012, 43, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.S.; Krmpotic, C.; Barbeito, C.; Soto, F.; Loza, C.M.; Vera, R.; Negrete, J. I’ve got you under my skin: Inflammatory response to elephant seal’s lice. Med. Vet. Entomol. 2021, 35, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.C. Ecology and morphological adaptation of the sucking lice (Anoplura, Echinophthiriidae) on the Northern Fur seal. Rapport et Procès verbaux des Réunions du conseil Permanent International pour lExploration de la Mer 1975, 169, 504–515. [Google Scholar]
- Leonardi, M.S.; Crespo, J.E.; Soto, F.A.; Vera, R.B.; Rua, J.C.; Lazzari, C.R. Under pressure: The extraordinary survival of seal lice in the deep sea. J. Exp. Biol. 2020, 223, jeb226811. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.S.; Crespo, E.A.; Raga, J.A.; Fernández, M. Redescription of Antarctophthirus microchir (Anoplura: Echinophthiriidae) from the South American sea lion, Otaria flavescens, from Patagonia, Argentina. J. Parasitol. 2009, 95, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.C. The sucking lice (Anoplura: Echinophthiriidae) of the northern fur seal; descriptions and morphological adaptation. Ann. Entomol. Soc. Am. 1971, 64, 280–292. [Google Scholar] [CrossRef]
- Murray, M.D. Insect parasite of marine bird and mammals. In Marine Insects; Cheng, L., Ed.; American Elsevier Publishing Company Inc.: New York, NY, USA, 1976; pp. 78–96. [Google Scholar]
- Aznar, F.J.; Leonardi, M.S.; Berón-Vera, B.; Vales, D.G.; Ameghino, S.; Raga, J.A.; Crespo, E.A. Population dynamics of Antarctophthirus microchir (Anoplura: Echinophthiriidae) in pups from South American sea lion, Otaria flavescens, in Northern Patagonia. Parasitology 2009, 136, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, M.S.; Crespo, E.A.; Raga, J.A.; Aznar, F.J. Lousy mums: Patterns of vertical transmission of an amphibious louse. Parasitol. Res. 2013, 112, 3315–3323. [Google Scholar] [CrossRef] [PubMed]
- Hinton, H.E. Respiratory adaptations of marine insects. In Marine Insects; Cheng, L., Ed.; American Elsevier Publishing Company Inc.: New York, NY, USA, 1976; pp. 43–78. [Google Scholar]
- Leonardi, M.S.; Lazzari, C.R. Uncovering deep mysteries: The underwater life of an amphibious louse. J. Insect Physiol. 2014, 71, 164–169. [Google Scholar] [CrossRef]
- Murray, M.D.; Smith, M.S.R.; Soucek, Z. Studies on the ectoparasites of seals and penguins. II. The ecology of the louse Antarctophthirus ogmorhini Enderlein on the Weddell seal, Leptonychotes weddelli Lesson. Aust. J. Zool. 1965, 13, 761–771. [Google Scholar] [CrossRef]
- Soto, F.A.; Klaich, M.J.; Negrete, J.; Leonardi, M.S. So happy together: Juvenile crabeater seal behavior improves lice transmission. Parasitol. Res. 2020, 119, 2059–2065. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.D.; Nicholls, D.G. Studies on the ectoparasites of seals and penguins I. The ecology of the louse Lepidophthirus macrorhini Enderlein on the Southern Elephant seal, Mirounga leonina. Aust. J. Zool. 1965, 13, 437–454. [Google Scholar] [CrossRef]
- Schorr, G.S.; Falcone, E.A.; Moretti, D.J.; Andrews, R.D. First longterm behavioral records from Cuvier’s beaked whales (Ziphius cavirostris) reveal record-breaking dives. PLoS ONE 2014, 9, e92633. [Google Scholar]
- Murray, M.D. Ecology of the ectoparasites of seals and penguins. In Proceedings of the 1st SCAR Symposium of Antarctic Biology, Paris, France, 2–8 September 1962; Carrick, R., Prevost, J., Holdgate, M.W., Eds.; Hermann: Paris, France, 1964; pp. 241–245. [Google Scholar]
- Murray, M.D. Ecology of the louse Lepidophthirus macrorhini Enderlein 1904 on the elephant seal Mirounga leonina (L.). Nature 1958, 182, 404–405. [Google Scholar] [CrossRef]
- Murray, M.D. Ectoparasites of Antarctic seals and birds. Jpn. Antarct. Res. Exped. Sci. Rep. 1967, 1, 185–191. [Google Scholar]
- Kim, K.C. Louse populations of the northern fur seal (Callorhinus ursinus). Am. J. Vet. Res. 1972, 33, 2027–2036. [Google Scholar]
- Leidenberger, S.; Harding, K.; Härkönen, T. Phocid seals, seal lice and heartworms: A terrestrial host-parasite system conveyed to the marine environment. Dis. Aquat. Org. 2007, 77, 235–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddrell, S.H.P. Why are there no insects in the open sea? J. Exp. Biol. 1998, 201, 2461–2464. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.F.; Woods, H.A.; Roberts, S.P. Ecological and Environmental Physiology of Insects; Oxford University Press: Oxford, UK, 2012. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Louse Genus | Species | Host |
|---|---|---|
| Antarctophthirus | A. callorhini | Northern fur seal |
| A. carlinii | Weddell seal | |
| A. lobodontis | Crabeater seal | |
| A. mawsoni | Ross seal | |
| A. microchir | Steller, Californian, South American, Australian, and New Zealand sea lion | |
| A. ogmorhini | Leopard seal | |
| A. trichechi | Walrus | |
| Latagophthirus | La. rauschi | North American river otter |
| Lepidophthirus | Le. macrorhini | Elephant seals |
| Le. piriformis | Monk seals | |
| Echinophthirius | E. horridus | Northern true seals |
| Proechinophthirus | P. fluctus | Northern fur seal |
| P. zumpti | Southern fur seals |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leonardi, M.S.; Crespo, J.E.; Soto, F.; Lazzari, C.R. How Did Seal Lice Turn into the Only Truly Marine Insects? Insects 2022, 13, 46. https://doi.org/10.3390/insects13010046
Leonardi MS, Crespo JE, Soto F, Lazzari CR. How Did Seal Lice Turn into the Only Truly Marine Insects? Insects. 2022; 13(1):46. https://doi.org/10.3390/insects13010046
Chicago/Turabian StyleLeonardi, María Soledad, José E. Crespo, Florencia Soto, and Claudio R. Lazzari. 2022. "How Did Seal Lice Turn into the Only Truly Marine Insects?" Insects 13, no. 1: 46. https://doi.org/10.3390/insects13010046
APA StyleLeonardi, M. S., Crespo, J. E., Soto, F., & Lazzari, C. R. (2022). How Did Seal Lice Turn into the Only Truly Marine Insects? Insects, 13(1), 46. https://doi.org/10.3390/insects13010046