
Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. DNA Extraction, PCR Conditions, and Sequencing
2.3. Multilocus Sequence Analysis (MLSA) of the EPF Strains
2.4. In Vitro Insecticide Activity of Buffer Suspension (B) of the Wettable Powder (WP) and Inverse Emulsion (IE) Formulations of EPF against Pupae with Silk Cocoons of L. botrana
2.5. Field Efficacy Assays
2.6. Statistical Analyses
3. Results
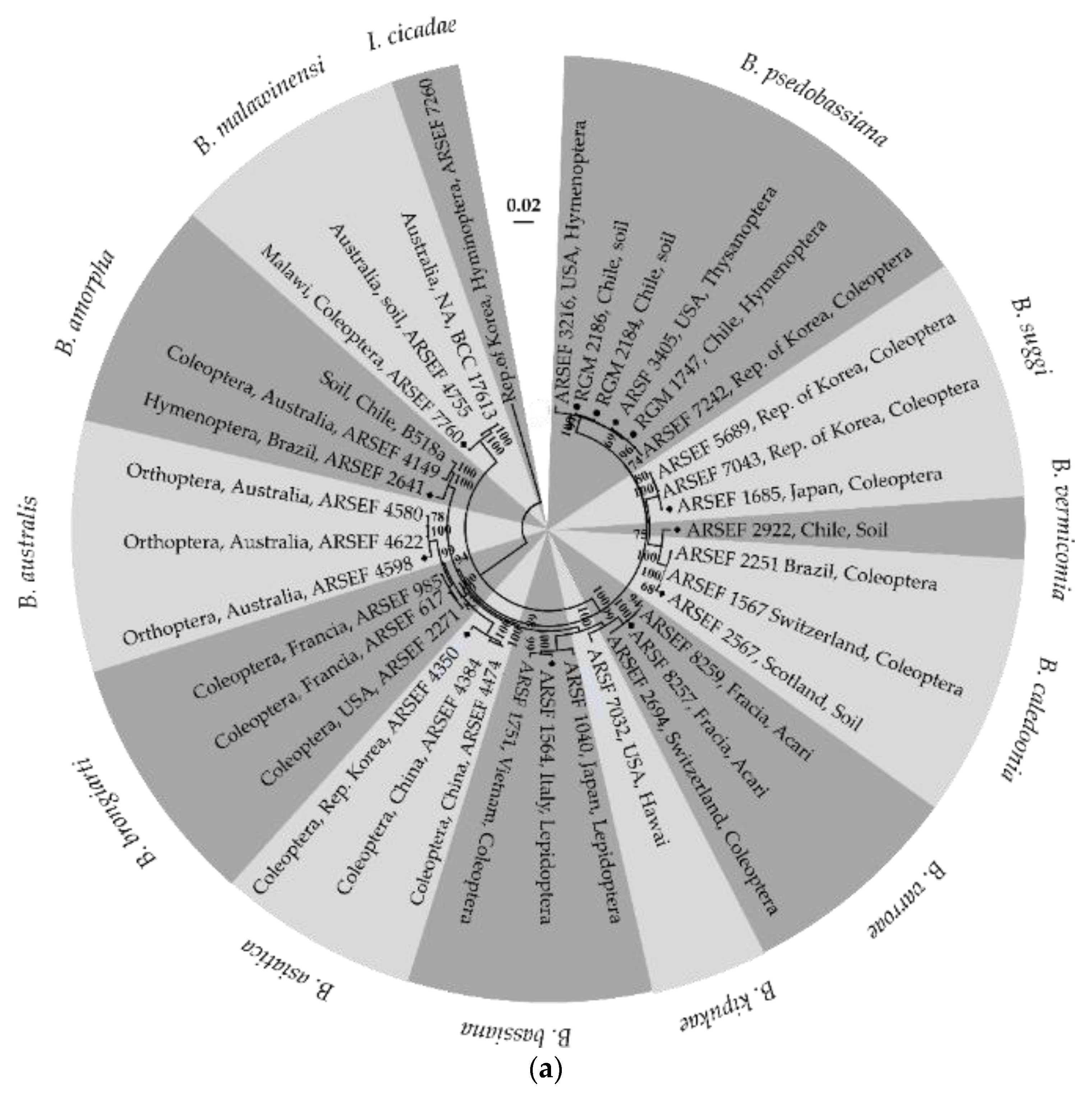
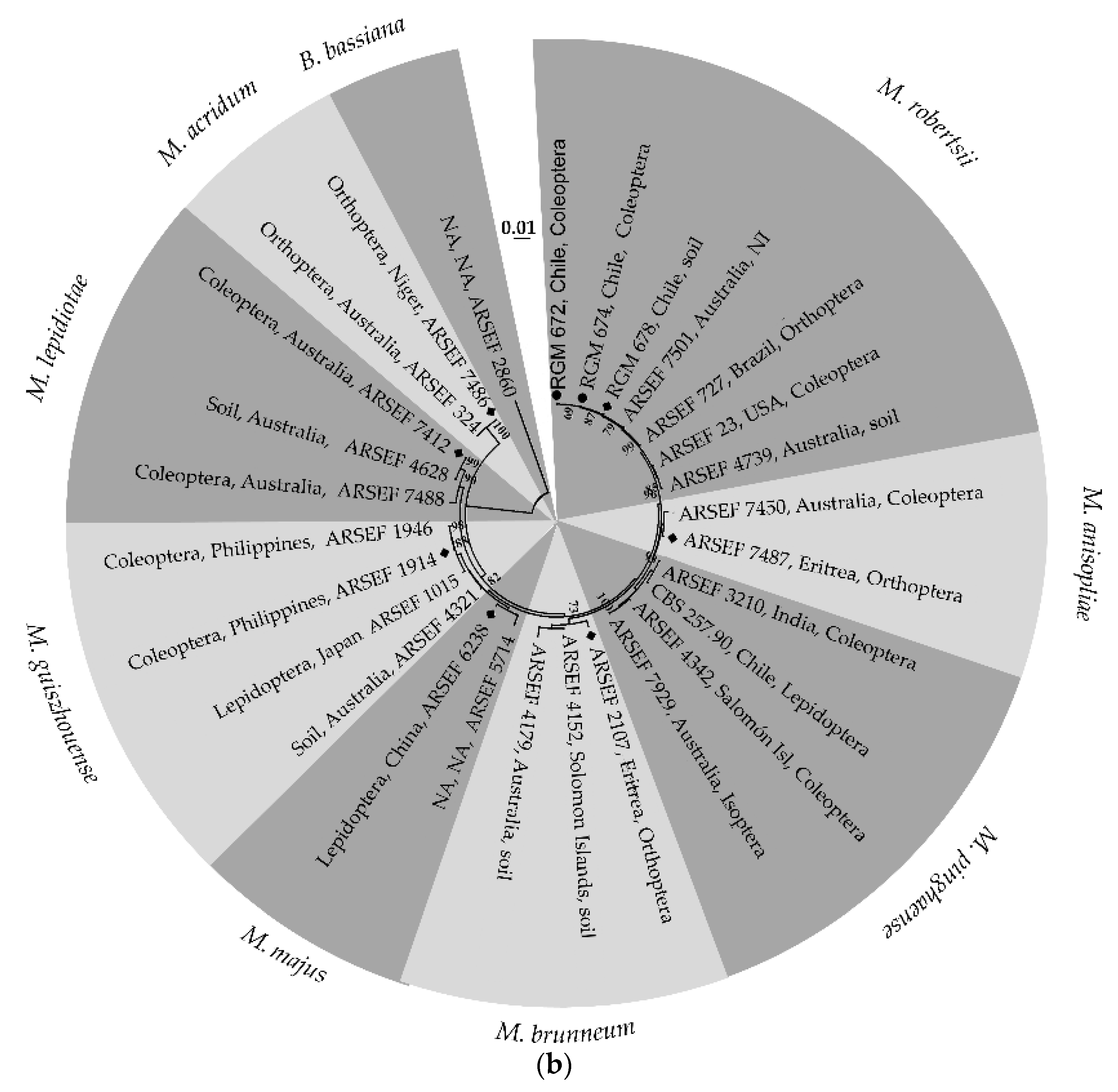
3.1. Molecular Identification of the EPF Strains
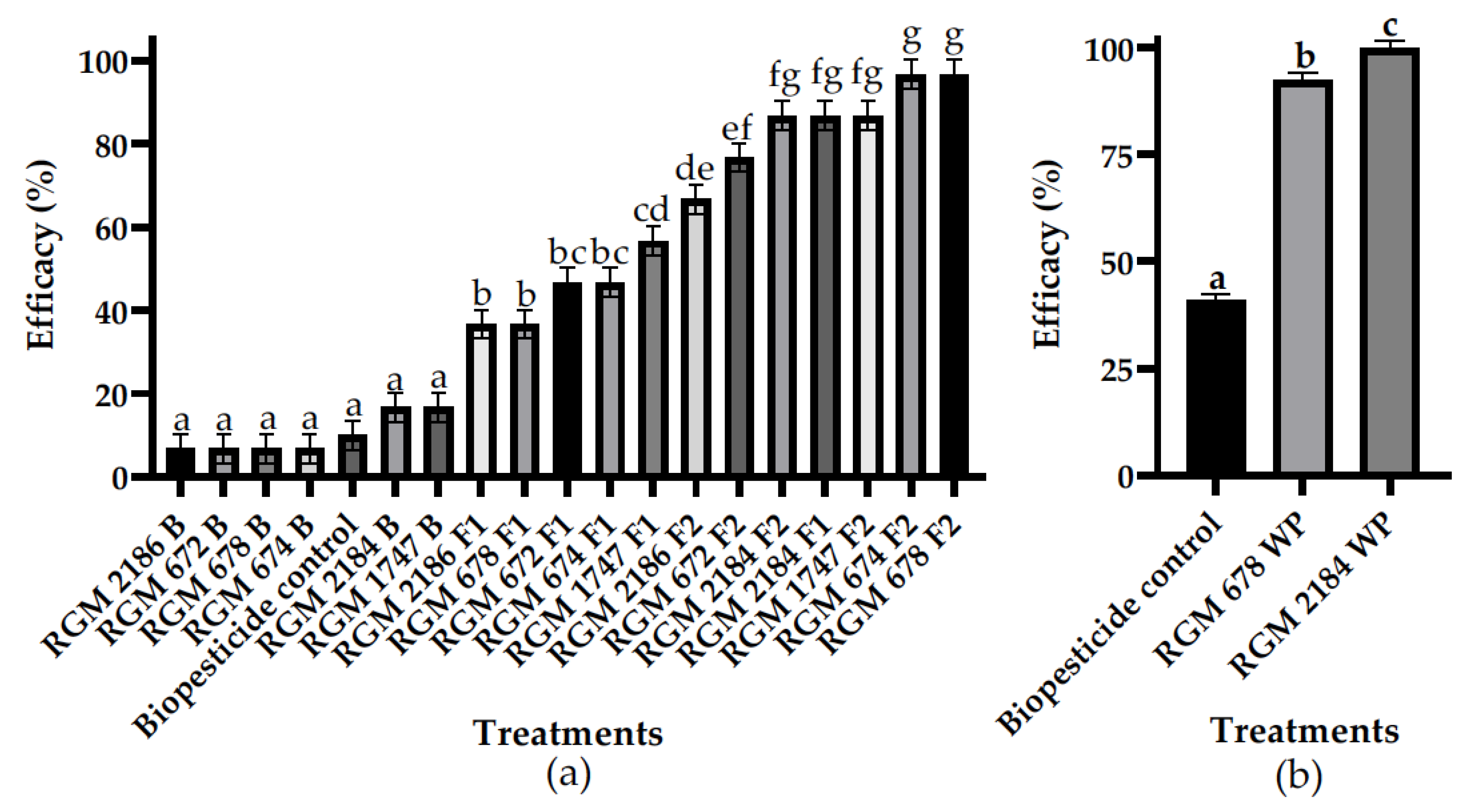
3.2. In Vitro Efficacy Assay Selection of Formulated EPF
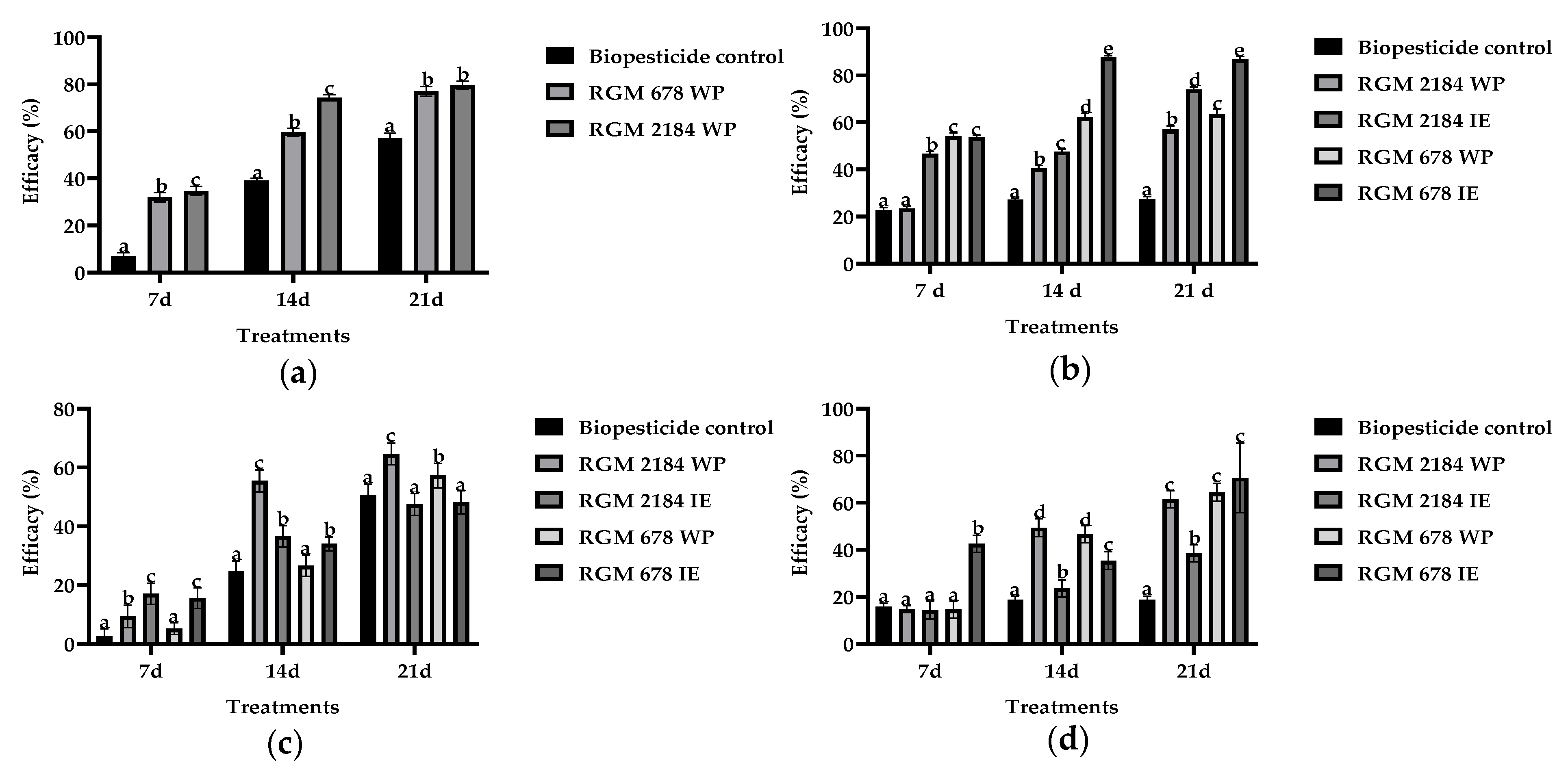
3.3. Field Efficacy Assay of the Formulated EPF
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilligan, T.M.; Epstein, M.E.; Passoa, S.C.; Powell, J.A.; Sage, O.C.; Brown, J.W. Discovery of Lobesia botrana ([Denis & Schiffermüller]) in California: An Invasive Species New to North America (Lepidoptera: Tortricidae). Proc. Entomol. Soc. Wash. 2011, 113, 14–30. (accessed on 1 June 2021). [Google Scholar] [CrossRef]
- Lobesia botrana (European Grapevine Moth). Available online: https://www.cabi.org/isc/datasheet/42794 (accessed on 1 June 2021).
- Declara Control Obligatorio de la Plaga Polilla del Racimo de la vid (Lobesia botrana). Available online: https://www.sag.gob.cl/sites/default/files/RESOLUCION_2109_2008.pdf (accessed on 1 June 2021).
- Martín-Vertedor, D.; Ferrero-García, J.J.; Torres-Vila, L.M. Global warming affects phenology and voltinism of Lobesia botrana in Spain. Agric. For. Entomol. 2010, 12, 169–176. [Google Scholar] [CrossRef]
- Castex, V.; de Cortázar-Atauri, I.G.; Calanca, P.; Beniston, M.; Moreau, J. Assembling and testing a generic phenological model to predict Lobesia botrana voltinism for impact studies. Ecol. Mode 2020, 420, 1–13. [Google Scholar] [CrossRef]
- Fermaud, M. Association of Botrytis cinerea with Grape Berry Moth Larvae. Phytopathology 1989, 79, 651–656. [Google Scholar] [CrossRef]
- Cozzi, G.; Somma, S.; Haidukowski, M.; Logrieco, A.F. Ochratoxin A Management in Vineyards by Lobesia botrana Biocontrol. Toxins 2013, 5, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Simmons, G.; Sepulveda, M.S.; Barrios, E.F.; Villegas, M.I.; Jimenez, R.M.; Jerez, A.G.; Henderson, R.; Riffo, H.D. Development of Sterile Insect Technique for Control of the European Grapevine Moth, Lobesia botrana, in Urban Areas of Chile. Insects 2021, 12, 378. [Google Scholar] [CrossRef]
- Lucchi, A.; Scaramozzino, P.L.; Loni, A. A review of insect parasitoids associated with Lobesia botrana (Denis & Schiffermüller, 1775) in Italy. 1. Diptera Tachinidae and Hymenoptera Braconidae (Lepidoptera, Tortricidae). ZooKeys 2017, 647, 67–100. [Google Scholar] [CrossRef]
- Plantey, R.L.; Papura, D.; Couture, C.; Thiéry, D.; Pizzuolo, P.H.; Bertoldi, M.V.; Lucero, G.S. Characterization of entomopathogenic fungi from vineyards in Argentina with potential as biological control agents against the European grapevine moth Lobesia botrana. BioControl 2019, 64, 501–511. [Google Scholar] [CrossRef]
- Beris, E.; Korkas, E. Additive and synergistic interactions of entomopathogenic fungi with Bacillus thuringiensis for the control of the European grapevine moth Lobesia botrana (Denis and Schiffermüller) (Lepidoptera: Tortricidae). Egypt. J. Biol. Pest Control. 2021, 31, 1–7. [Google Scholar] [CrossRef]
- Biswas, T.; Joop, G.; Rafaluk-Mohr, C. Cross-Resistance: A consequence of bi-partite host-parasite coevolution. Insects 2018, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Qasim, M.; Islam, S.U.; Islam, W.; Noman, A.; Khan, K.A.; Hafeez, M.; Hussain, D.; Dash, C.K.; Bamisile, B.S.; Akutse, K.S.; et al. Characterization of mycotoxins from entomopathogenic fungi (Cordyceps fumosorosea) and their toxic effects to the development of asian citrus psyllid reared on healthy and diseased citrus plants. Toxicon 2020, 188, 39–47. [Google Scholar] [CrossRef]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? Insights into alternative modes of action of entomopathogenic fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-athogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [CrossRef]
- Altimira, F.; De La Barra, N.; Rebufel, P.; Soto, S.; Soto, R.; Estay, P.; Vitta, N.; Tapia, E. Potential biological control of the pupal stage of the European grapevine moth Lobesia botrana by the entomopathogenic fungus Beauveria pseudobassiana in the winter season in Chile. BMC Res. Notes 2019, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Minnis, A.M.; Sung, G.-H.; Luangsa-Ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia 2009, 101, 512–530. [Google Scholar] [CrossRef] [Green Version]
- Kepler, R.M.; Rehner, S.A. Genome-assisted development of nuclear intergenic sequence markers for entomopathogenic fungi of the Metarhizium anisopliae species complex. Mol. Ecol. Resour. 2013, 13, 210–217. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tapia, E.; Altimira, F.; De La Barra, N.; Vitta, N.; Estay, P. Composición biopesticida en base a hongos ento-mopatógenos nativos para el biocontrol y/o manejo integrado de Lobesia botrana en vides, ciruelos y arándanos, y método de aplicación de dicha composición, en diapausa invernal, en pupas ubicadas en zonas posteriores/no expuestas del ritidoma de la planta. INAPI 201802396, 4 May 2021. Available online: https://patents.google.com/patent/CL2018002396A1/es (accessed on 1 June 2021).
- Abbott, W.S. A Method of computing the effectiveness of an insecticide. 1925. J. Econ. Entomol. 1987, 3, 302–303. [Google Scholar]
- Henderson, C.F.; Tilton, E.W. Tests with acaricides against the brow wheat mite. J. Econ. Entomol. 1985, 48, 157–161. [Google Scholar] [CrossRef]
- Aguilera, J.A.; Deymié, M.C.; Herrera, M.; Vazquez, F.; Cuthbertson, G.S.; Lopez-Lastra, C.; Lechner, B. The entomopathogenic fungus, Metarhizium anisopliae for the European grapevine moth, Lobesia botrana Den. & Schiff. (Lepidoptera: Tortricidae) and its effect to the phytopathogenic fungus, Botrytis cinerea. Egypt. J. Biol. Pest Control. 2018, 28, 1–8. [Google Scholar] [CrossRef]
- Danks, H.V. The roles of insect cocoons in cold conditions. Eur. J. Entomol. 2004, 101, 433–437. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altimira, F.; De La Barra, N.; Godoy, P.; Roa, J.; Godoy, S.; Vitta, N.; Tapia, E. Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile. Insects 2022, 13, 8. https://doi.org/10.3390/insects13010008
Altimira F, De La Barra N, Godoy P, Roa J, Godoy S, Vitta N, Tapia E. Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile. Insects. 2022; 13(1):8. https://doi.org/10.3390/insects13010008
Chicago/Turabian StyleAltimira, Fabiola, Nathalia De La Barra, Paulo Godoy, Juan Roa, Sebastián Godoy, Nancy Vitta, and Eduardo Tapia. 2022. "Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile" Insects 13, no. 1: 8. https://doi.org/10.3390/insects13010008
APA StyleAltimira, F., De La Barra, N., Godoy, P., Roa, J., Godoy, S., Vitta, N., & Tapia, E. (2022). Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile. Insects, 13(1), 8. https://doi.org/10.3390/insects13010008