
Optimising Vine Weevil, Otiorhynchus sulcatus F. (Coleoptera: Curculionidae), Monitoring Tool Design

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Cultures
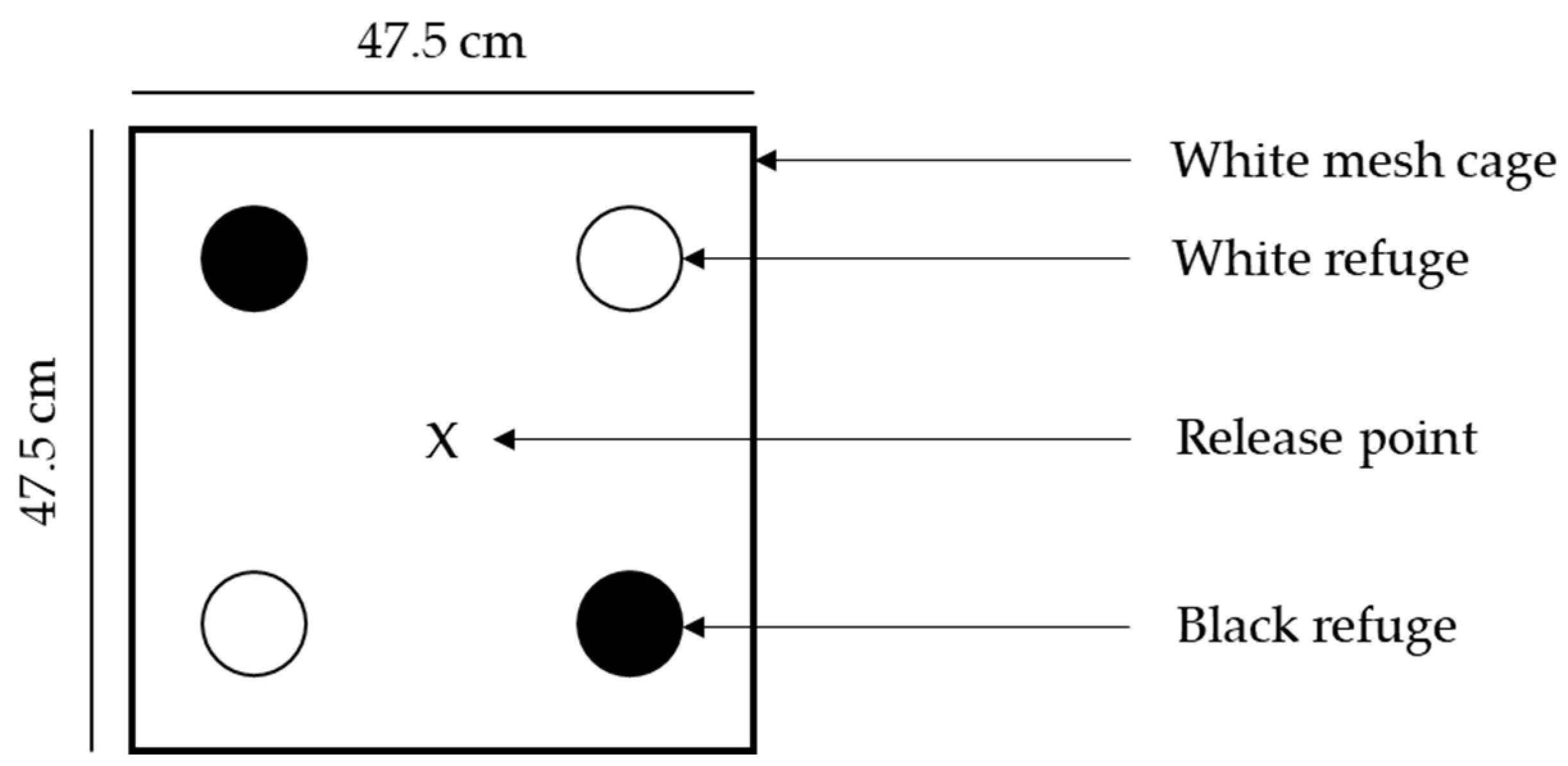
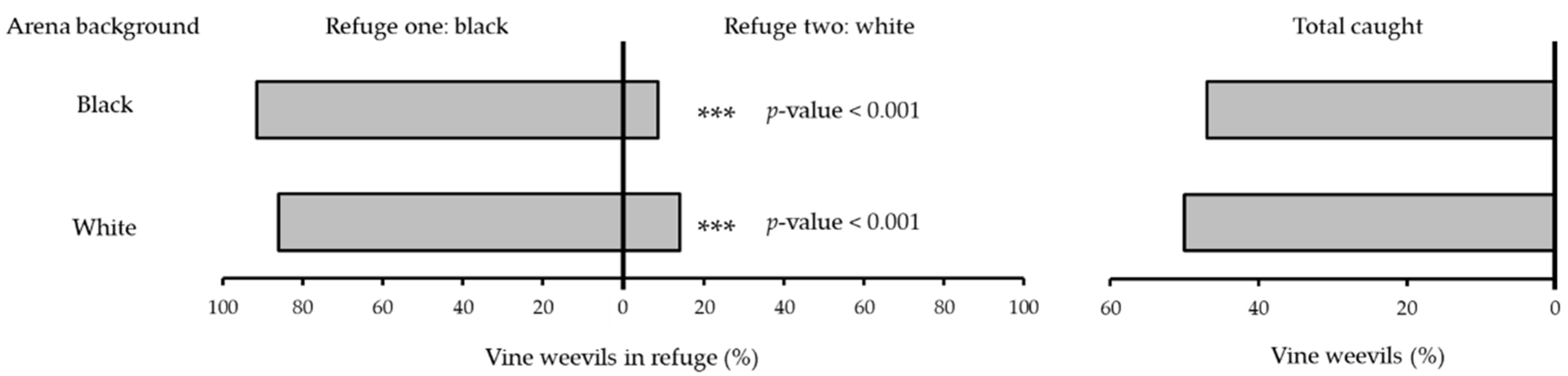
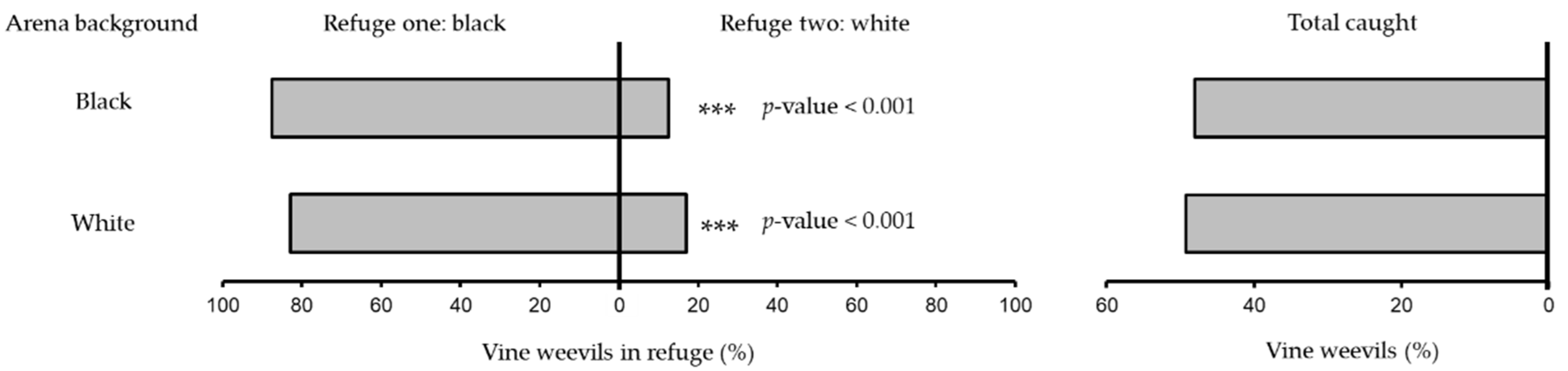
2.2. Refuge Colour (Dark vs. Light)
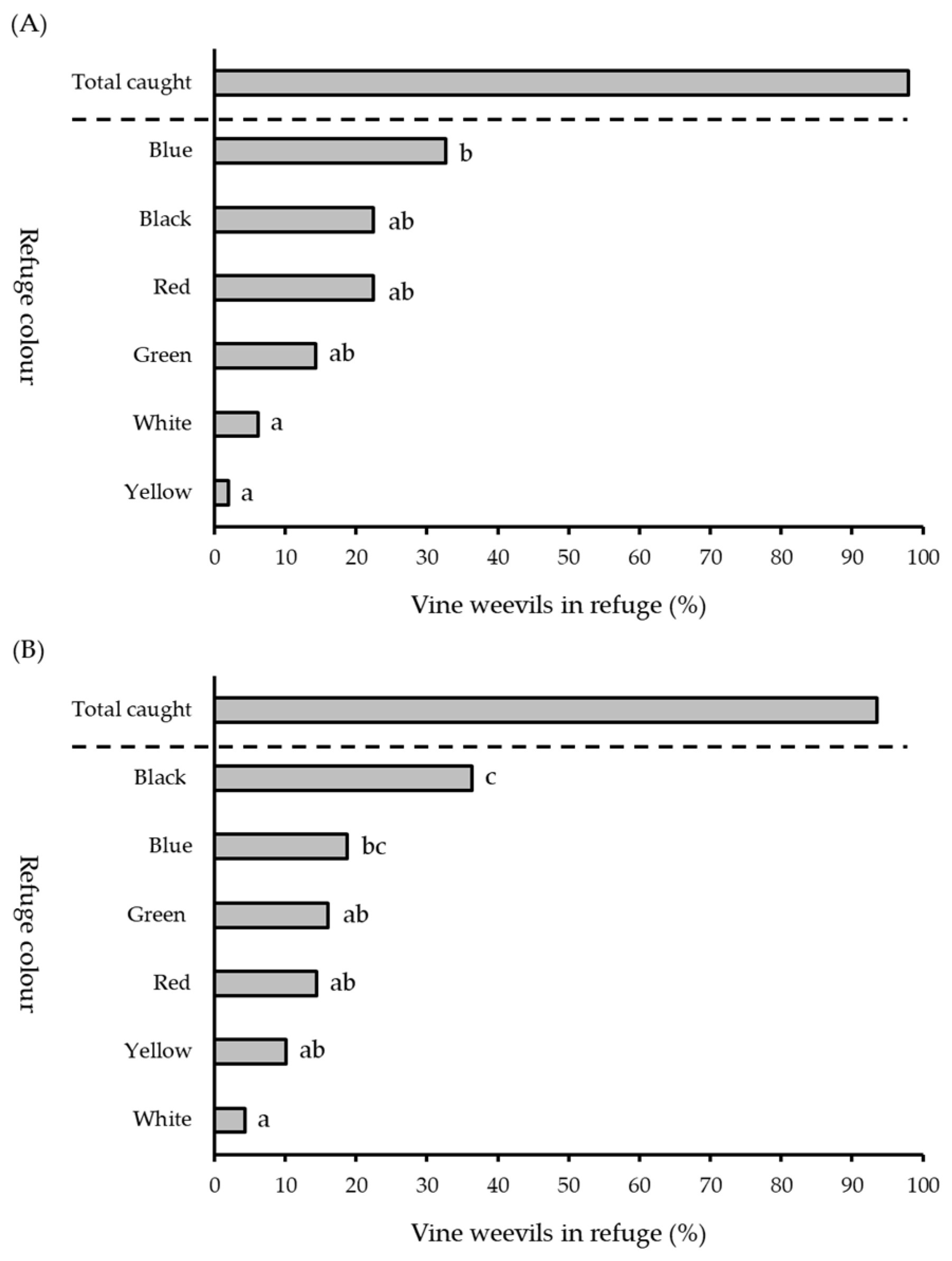
2.3. Refuge Colour (Multiple)
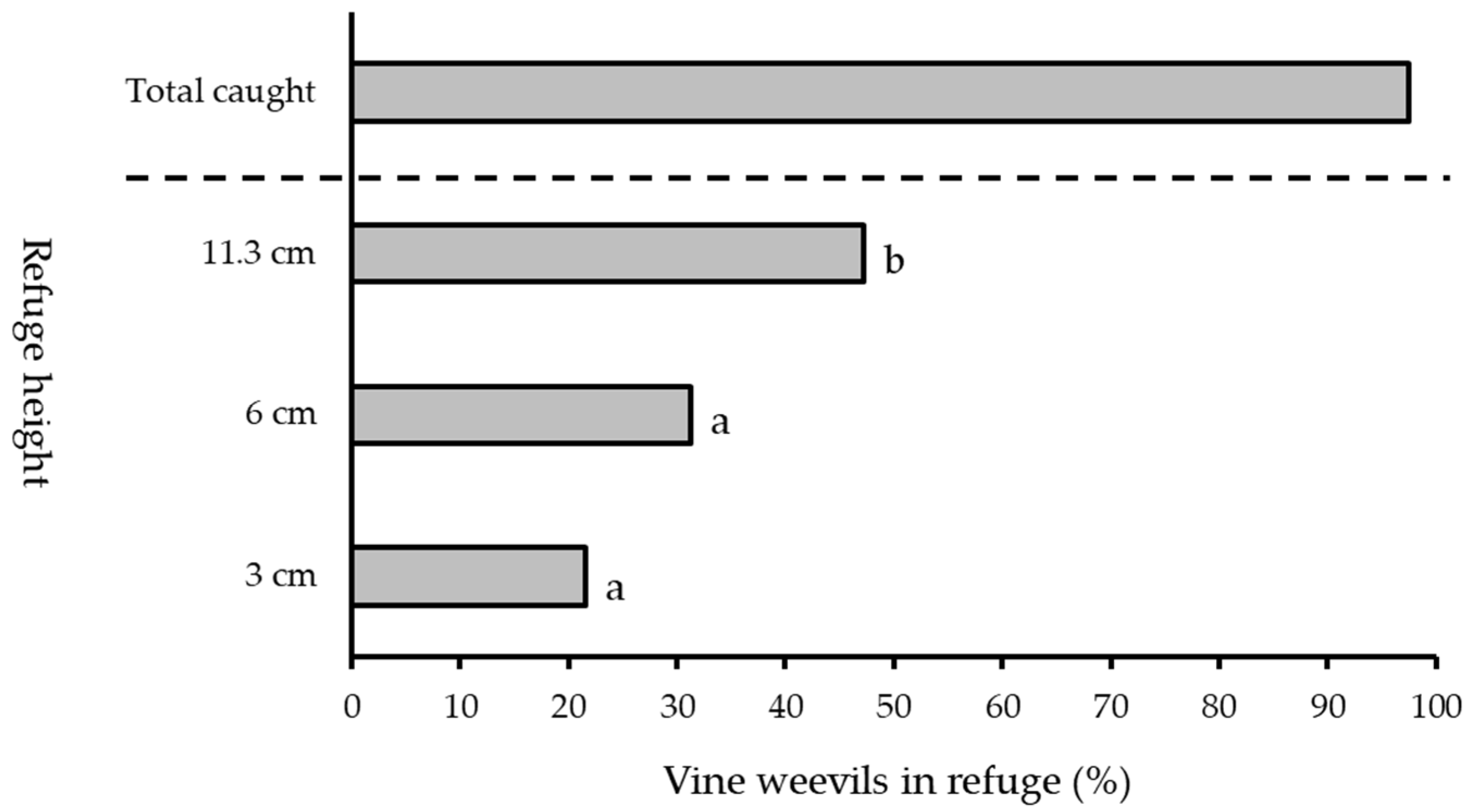
2.4. Refuge Height
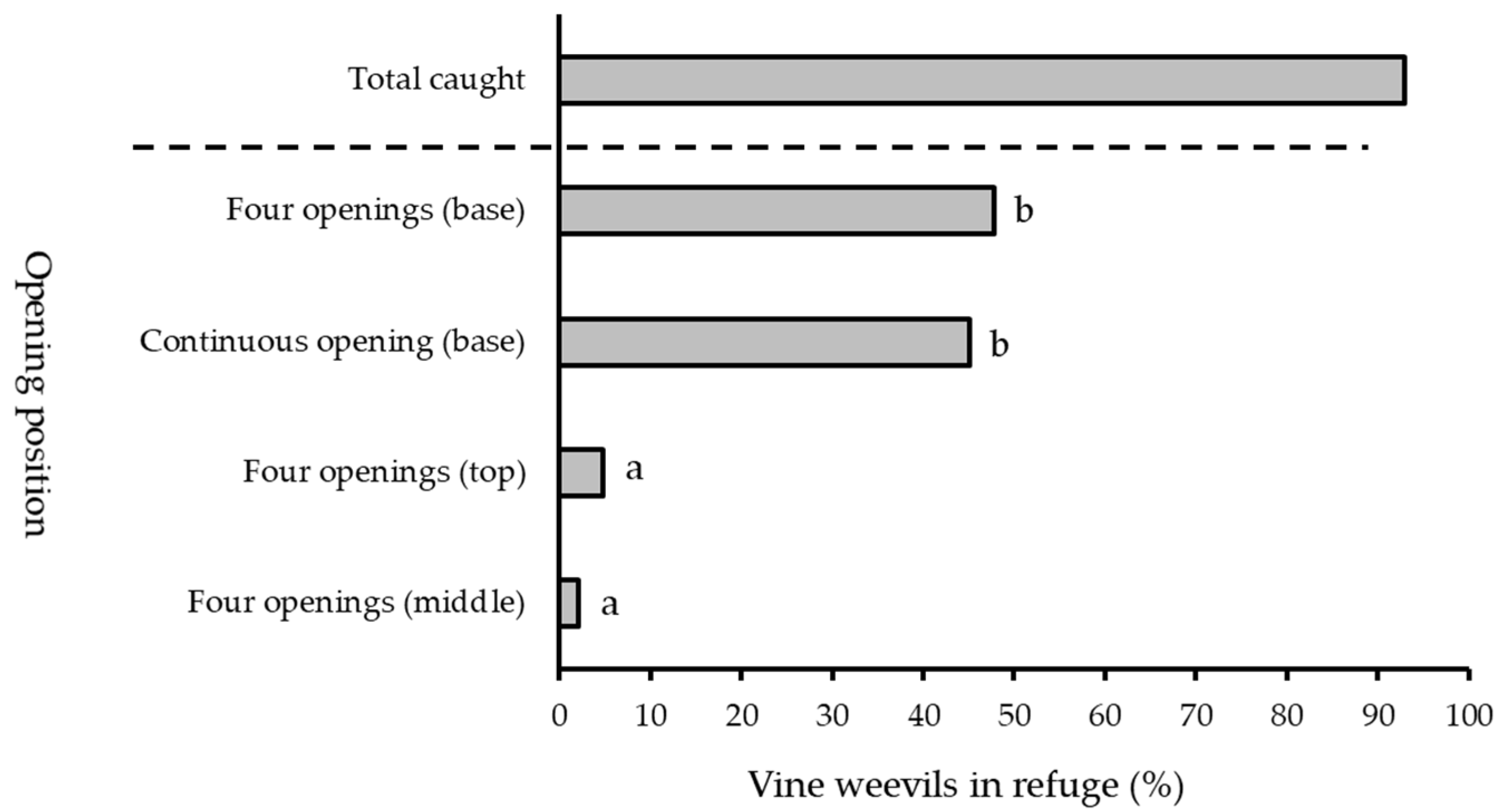
2.5. Refuge Entrance Position
2.6. Statistical Analysis
3. Results
3.1. Refuge Colour (Dark vs. Light)
3.2. Refuge Colour (Multiple)
3.3. Refuge Height
3.4. Refuge Entrance Positions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moorhouse, E.R.; Charnley, A.K.; Gillespie, A.T. A Review of the biology and control of the vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae). Ann. Appl. Biol. 1992, 121, 431–454. [Google Scholar] [CrossRef]
- Van Tol, R.W.H.M.; Bruck, D.J.; Griepink, F.C.; De Kogel, W.J. Field attraction of the vine weevil Otiorhynchus sulcatus to kairomones. J. Econ. Entomol. 2012, 105, 169–175. [Google Scholar] [CrossRef]
- CABI Datasheet: Otiorhynchus Sulcatus (Vine Weevil). 2021. Available online: https://www.cabi.org/isc/datasheet/38071 (accessed on 21 June 2021).
- Evenhuis, H.H. Bionomics and control of the black vine weevil Otiorhynchus sulcatus. Meded. Fac. Landbouwwet. Rijksuniv. Gent 1978, 43, 607–611. [Google Scholar]
- Mason, E.C. Observations on the life history and control of the vine weevil on cyclamen and foliage plants. Plant. Pathol. 1960, 9, 29–33. [Google Scholar] [CrossRef]
- Cone, W.W. The black vine weevil, Brachyrhinus sulcatus, as a pest of grapes in South Central Washington1. J. Econ. Entomol. 1963, 56, 677–680. [Google Scholar] [CrossRef]
- Penman, D.R.; Scott, R.R. Adult emergence and egg production of the black vine weevil in Canterbury. N. Z. J. Exp. Agric. 1976, 4, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Watt, K.; Graham, J.; Gordon, S.C.; Woodhead, M.; Mcnicol, R.J. Current and future transgenic control strategies to vine weevil and other insect resistance in strawberry. J. Hortic. Sci. Biotechnol. 1999, 74, 409–421. [Google Scholar] [CrossRef]
- Bruck, D.J. Effect of potting media components on the infectivity of Metarhizium anisopliae against the black vine weevil (Coleoptera: Curculionidae). J. Environ. Hortic. 2006, 24, 91–94. [Google Scholar] [CrossRef]
- Cowles, R.S. Protecting container-grown crops from black vine weevil larvae with bifenthrin. J. Environ. Hortic. 2001, 19, 184–189. [Google Scholar] [CrossRef]
- Smith, F.F. Biology and Control of the Black Vine Weevi; US Department of Agriculture: Washington, DC, USA, 1932; Volume 325.
- Blackshaw, R.P. Studies on the chemical control of vine weevil larvae in hardy-ornamental nursery stock. J. Hortic. Sci. 1987, 62, 63–66. [Google Scholar] [CrossRef]
- Blackshaw, R.P.; O’Neill, S. Chlorpyrifos compost treatment for vine weevil control: Growth of ornamental plants compared with an aldrin standard. J. Hortic. Sci. 1987, 62, 67–69. [Google Scholar] [CrossRef]
- Cross, J.V.; Buxton, J.H.; Jacobson, R.; Richardson, D.M. Chemical control of vine weevil larvae on container-grown hardy ornamental nursery stock 1986–1989. Ann. Appl. Biol. 1995, 127, 533–542. [Google Scholar] [CrossRef]
- Reding, M.E.; Persad, A.B. Systemic insecticides for control of black vine weevil (Coleoptera: Curculionidae) in container- and field-grown nursery crops. J. Econ. Entomol. 2009, 102, 927–933. [Google Scholar] [CrossRef]
- Reding, M.E.; Ranger, C.M. Systemic insecticides reduce feeding, survival, and fecundity of adult black vine weevils (Coleoptera: Curculionidae) on a variety of ornamental nursery crops. J. Econ. Entomol. 2011, 104, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Hillocks, R.J. Farming with fewer pesticides: EU pesticide review and resulting challenges for UK agriculture. Crop. Prot. 2012, 31, 85–93. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Q.; Lu, W.; Liu, F. Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennis. J. Pest. Sci. 2015, 88, 383–392. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messéan, A. Toward a reduced reliance on conventional pesticides in European agriculture. Plant Dis. 2016, 100, 10–24. [Google Scholar] [CrossRef] [Green Version]
- Georgis, R.; Poinar, G.J.; Wilson, A.P. Susceptibility of strawberry root weevil Otiorhynchus sulcatus to Neoaplectanid and Heterorhabditid nematodes. IRCS Med. Sci. Microbiol. Parasitol. Infect. Dis. 1982, 10, 442. [Google Scholar]
- Moorhouse, E.R.; Easterbrook, M.A.; Gillespie, A.T.; Charnley, A.K. Control of Otiorhynchus sulcatus (Fabricius) (Coleoptera: Curculionidae) larvae on a range of hardy ornamental nursery stock species using the entomopathogenic fungus Metarhizium anisopliae. Biocontrol. Sci. Technol. 1993, 3, 63–72. [Google Scholar] [CrossRef]
- Lola-Luz, T.; Downes, M.; Dunne, R. Control of black vine weevil larvae Otiorhynchus sulcatus (Fabricius) (Coleoptera: Curculionidae) in grow bags outdoors with nematodes. Agric. For. Entomol. 2005, 7, 121–126. [Google Scholar] [CrossRef]
- Shah, F.A.; Ansari, M.A.; Prasad, M.; Butt, T.M. Evaluation of black vine weevil (Otiorhynchus sulcatus) control strategies using Metarhizium anisopliae with sublethal doses of insecticides in disparate horticultural growing media. Biol. Control. 2007, 40, 246–252. [Google Scholar] [CrossRef]
- Ansari, M.A.; Shah, F.A.; Butt, T.M. Combined use of entomopathogenic nematodes and Metarhizium anisopliae as a new approach for black vine weevil, Otiorhynchus sulcatus, control. Entomol. Exp. Appl. 2008, 129, 340–347. [Google Scholar] [CrossRef]
- Pope, T.W.; Hough, G.; Arbona, C.; Roberts, H.; Bennison, J.; Buxton, J.; Prince, G.; Chandler, D. Investigating the potential of an autodissemination system for managing populations of vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae) with entomopathogenic fungi. J. Invertebr. Pathol. 2018, 154, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higley, L.G.; Pedigo, L.P. Economic injury level concepts and their use in sustaining environmental quality. Agric. Ecosyst. Environ. 1993, 46, 233–243. [Google Scholar] [CrossRef]
- Kogan, M. Integrated Pest Management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef]
- Ramsden, M.W.; Kendall, S.L.; Ellis, S.A.; Berry, P.M. A review of economic thresholds for invertebrate pests in UK arable crops. Crop. Prot. 2017, 96, 30–43. [Google Scholar] [CrossRef]
- Li, S.Y.; Fitzpatrick, S.M.; Henderson, D.E. Grooved board traps for monitoring the black vine weevil (Coleoptera: Curculionidae) in raspberry fields. J. Entomol. Soc. Br. Columbia 1995, 92, 97–100. [Google Scholar]
- Bennison, J.; Allen, J.; Atwood, J.; Pope, T.W. Vine Weevil in Soft Fruit Crops; AHDB (Agriculture and Horticulture Development Board). 2018. Available online: https://ahdb.org.uk/knowledge-library/vine-weevil-control-in-soft-fruit-crops (accessed on 19 June 2021).
- Gordon, S.C.; Woodford, J.A.T.; Grassi, A.; Zini, M.; Tuovinen, T.; Lindqvist, I.; McNicol, J.W. Monitoring and importance of wingless weevils (Otiorhynchus spp.) in European red raspberry production. IOBC-WPRS Bull. 1995, 26, 55–60. [Google Scholar]
- Hanula, J.L. Monitoring Adult emergence, ovary maturation, and control of the black vine weevil (Coleoptera: Curculionidae). J. Entomol. Sci. 1990, 25, 134–142. [Google Scholar] [CrossRef]
- Phillips, P.A. Simple monitoring of black vine weevil in vineyards. Calif. Agric. 1989, 43, 12–13. [Google Scholar]
- Van Tol, R.W.H.M.; Elberse, I.A.M.; Bruck, D.J. Development of a refuge-kairomone device for monitoring and control of the vine weevil, Otiorhynchus sulcatus, by lure-and-kill and lure-and-infect. Crop. Prot. 2020, 129, 105045. [Google Scholar] [CrossRef]
- Roberts, J.M.; Jahir, A.; Graham, J.; Pope, T.W. Catch me if you can: The influence of refuge/trap design, previous feeding experience, and semiochemical lures on vine weevil (Coleoptera: Curculionidae) monitoring success. Pest. Manag. Sci. 2020, 76, 553–560. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Owens, E.D. Visual detection of plants by herbivorous insects. Annu. Rev. Entomol. 1983, 28, 337–364. [Google Scholar] [CrossRef]
- Mizell, R.F.; Tedders, W.L. Evaluation of trap type and color for monitoring Hylobius pales and Pachylobius picivorus in Florida. Fla. Entomol. 1999, 82, 615–624. [Google Scholar] [CrossRef]
- Leskey, T.C. Visual cues and capture mechanisms associated with traps for plum curculio (Coleoptera: Curculiondae). J. Entomol. Sci. 2006, 41, 97–106. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Giannone, F.; Moino, S.; Rassati, D. Exploiting trap color to improve surveys of longhorn beetles. J. Pest. Sci. 2021, 94, 871–883. [Google Scholar] [CrossRef]
- Riley, D.G.; Schuster, D.J. Pepper weevil adult response to coloured sticky traps in pepper fields. Southwest Entomol. 1994, 19, 93–107. [Google Scholar]
- Silva, D.; Salamanca, J.; Kyryczenko-Roth, V.; Alborn, H.T.; Rodriguez-Saona, C. Comparison of trap types, placement, and colors for monitoring Anthonomus musculus (Coleoptera: Curculionidae) adults in highbush blueberries. J. Insect Sci. 2018, 18. [Google Scholar] [CrossRef]
- Abuagla, A.M.; Al-Deeb, M.A. Effect of bait quantity and trap color on the trapping efficacy of the pheromone trap for the red palm weevil, Rhynchophorus ferrugineus. J. Insect Sci. 2012, 12, 120. [Google Scholar] [CrossRef] [Green Version]
- Al-Saoud, A.H. Effect of ethyl acetate and trap colour on weevil captures in red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae) pheromone traps. Int. J. Trop. Insect Sci. 2013, 33, 202–206. [Google Scholar] [CrossRef]
- Hoover, S.E.R.; Lindgren, B.S.; Keeling, C.I.; Slessor, K.N. Enantiomer preference of Trypodendron lineatum and effect of pheromone dose and trap length on response to lineatin-baited traps in interior British Columbia. J. Chem. Ecol. 2000, 26, 667–677. [Google Scholar] [CrossRef]
- Reddy, G.; Balakrishnan, S.; Remolona, J.E.; Kikuchi, R.; Bamba, J.P. Influence of trap type, size, color, and trapping location on capture of Rhabdoscelus obscurus (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 2011, 104, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Epsky, N.D.; Shuman, D. Hole density and capture of stored-product insect pests in grain probe traps. J. Econ. Entomol. 2002, 95, 1326–1332. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, C.; Samietz, J.; Dorn, S. Visual orientation of overwintered Anthonomus pomorum (Coleoptera: Curculionidae). Environ. Entomol. 2004, 33, 1410–1415. [Google Scholar] [CrossRef] [Green Version]
- Reddy, G.V.P.; Cruz, Z.T.; Guerrero, A. Development of an efficient pheromone-based trapping method for the banana root borer Cosmopolites sordidus. J. Chem. Ecol. 2009, 35, 111–117. [Google Scholar] [CrossRef]
- Tansey, J.A.; Dosdall, L.M.; Keddie, B.A.; Noble, S.D. Contributions of visual cues to cabbage seedpod weevil, Ceutorhynchus obstrictus (Marsham) (Coleoptera: Curculionidae), resistance in novel host genotypes. Crop. Prot. 2010, 29, 476–481. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. 2019. Available online: https://www.r-project.org/ (accessed on 21 February 2021).
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1. 3-1. 2019. Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 27 February 2021).
- Rojas, J.C.; Wyatt, T.D. Role of visual cues and interaction with host odour during the host-finding behaviour of the cabbage moth. Entomol. Exp. Appl. 1999, 91, 59–65. [Google Scholar] [CrossRef]
- Bruce, T.J.A. Interplay between insects and plants: Dynamic and complex interactions that have coevolved over millions of years but act in milliseconds. J. Exp. Bot. 2015, 66, 455–465. [Google Scholar] [CrossRef]
- Piñero, J.C.; Jácome, I.; Vargas, R.; Prokopy, R.J. Response of female melon fly, Bactrocera cucurbitae, to host-associated visual and olfactory stimuli. Entomol. Exp. Appl. 2006, 121, 261–269. [Google Scholar] [CrossRef]
- Lebesa, L.N.; Khan, Z.R.; Hassanali, A.; Pickett, J.A.; Bruce, T.J.A.; Skellern, M.; Krüger, K. Responses of the blister beetle Hycleus apicicornis to visual stimuli. Physiol. Entomol. 2011, 36, 220–229. [Google Scholar] [CrossRef]
- Cook, S.M.; Skellern, M.P.; Döring, T.F.; Pickett, J.A. Red oilseed rape? The potential for manipulation of petal colour in control strategies for the pollen beetle (Meligethes aeneus). Arthropod-Plant Interact. 2013, 7, 249–258. [Google Scholar] [CrossRef]
- Hesler, L.S.; Sutter, G.R. Effect of trap color, volatile attractants, and type of toxic bait dispenser on captures of adult corn rootworm beetles (Coleoptera: Chrysomelidae). Environ. Entomol. 1993, 22, 743–750. [Google Scholar] [CrossRef]
- Roubos, C.R.; Liburd, O.E. Effect of trap color on captures of grape root borer (Lepidoptera: Sesiidae) males and non-target insects. J. Agric. Urban. Entomol. 2008, 25, 99–109. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, T.; Su, Y.; Wang, Z.; Zhang, X.; Wang, S.; Liu, Y. Influence of trap color, type, and placement on capture efficacy for Protaetia brevitarsis (Coleoptera: Scarabaeidae). J. Econ. Entomol. 2021, 114, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Hallett, R.H.; Oehlschlager, A.C.; Borden, J.H. Pheromone trapping protocols for the asian palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Int. J. Pest. Manag. 1999, 45, 231–237. [Google Scholar] [CrossRef]
- Kern, R.; Egelhaaf, M.; Srinivasan, M.V. Edge detection by landing honeybees: Behavioural analysis and model simulations of the underlying mechanism. Vision Res. 1997, 37, 2103–2117. [Google Scholar] [CrossRef] [Green Version]
- Lehrer, M.; Campan, R. Shape discrimination by wasps (Paravespula germanica) at the food source: Generalization among various types of contrast. J. Comp. Physiol. A 2004, 190, 651–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Entwistle, P.F. Some evidence for a colour sensitive phase in the light period of Scolytidae and Platypodidae. Entomol. Exp. Appl. 1963, 6, 143–148. [Google Scholar] [CrossRef]
- Timmons, G.M.; Potter, D.A. Influence of pheromone trap color on capture of lilac borer males. Environ. Entomol. 1981, 10, 756–759. [Google Scholar] [CrossRef]
- Wang, X.; Ji, Y.-C.; Wen, C.; Zhang, G.-Y.; Wen, J.-B. Effects of trap color and shape on the capture of Eucryptorrhynchus scrobiculatus (Coleoptera: Curculionidae). J. Econ. Entomol. 2019, 112, 2744–2750. [Google Scholar] [CrossRef]
- Tedders, W.L.; Mizell, R.F.; Wood, B.W. Effects of color and trunk-wrap on pecan weevil catch in pyramidal traps. J. Entomol. Sci. 1996, 31, 414–419. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Q.-H.; Wang, Y.; Liu, G.-T.; Zhou, X.; Niu, J.; Schlyter, F. Catching Ips duplicatus (Sahlberg) (Coleoptera: Scolytidae) with pheromone-baited traps: Optimal trap type, colour, height and distance to infestation. Pest. Manag. Sci. 2010, 66, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V.P.; Fettköther, R.; Noldt, U.; Dettner, K. Capture of female Hylotrupes bajulus as influenced by trap type and pheromone blend. J. Chem. Ecol. 2005, 31, 2169–2177. [Google Scholar] [CrossRef] [PubMed]
- Sajeewani, P.A.H.; Dissanayaka, D.M.S.K.; Wijayaratne, L.K.W.; Burks, C.S. Changes in shape, texture and airflow improve efficiency of monitoring traps for Tribolium castaneum (Coleoptera: Tenebrionidae). Insects 2020, 11, 778. [Google Scholar] [CrossRef]
- Vandersar, T.J.D.; Borden, J.H. Visual orientation of Pissodes strobi Peck (Coleoptera: Curculionidae) in relation to host selection behaviour. Can. J. Zool. 1977, 55, 2042–2049. [Google Scholar] [CrossRef]
- Toews, M.D.; Phillips, T.W. Factors affecting capture of Cryptolestes ferrugineus (Coleoptera: Laemophloeidae) in traps placed in stored wheat. J. Econ. Entomol. 2002, 95, 200–207. [Google Scholar] [CrossRef]
- Lee, J.C.; Burrack, H.J.; Barrantes, L.D.; Beers, E.H.; Dreves, A.J.; Hamby, K.A.; Haviland, D.R.; Isaacs, R.; Richardson, T.A.; Shearer, P.W.; et al. Evaluation of monitoring traps for Drosophila suzukii (Diptera: Drosophilidae) in North America. J. Econ. Entomol. 2012, 105, 1350–1357. [Google Scholar] [CrossRef] [Green Version]
- Renkema, J.M.; Buitenhuis, R.; Hallett, R.H. Optimizing trap design and trapping protocols for Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2014, 107, 2107–2118. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Reddy, K.V.S.; Sikora, R.A. Reduction of banana weevil populations with pseudostem traps. Int. J. Pest. Manag. 1994, 40, 300–304. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Raman, A. Visual cues are relevant in behavioral control measures for Cosmopolites sordidus (Coleoptera: Curculionidae). J. Econ. Entomol. 2011, 104, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Kelber, A. Ovipositing butterflies use a red receptor to see green. J. Exp. Biol. 1999, 202, 2619–2630. [Google Scholar] [CrossRef] [PubMed]
- Dyer, A.G.; Chittka, L. Biological significance of distinguishing between similar colours in spectrally variable illumination: Bumblebees (Bombus terrestris) as a case study. J. Comp. Physiol. 2004, 190, 105–114. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fezza, E.; Roberts, J.M.; Bruce, T.J.A.; Walsh, L.E.; Gaffney, M.T.; Pope, T.W. Optimising Vine Weevil, Otiorhynchus sulcatus F. (Coleoptera: Curculionidae), Monitoring Tool Design. Insects 2022, 13, 80. https://doi.org/10.3390/insects13010080
Fezza E, Roberts JM, Bruce TJA, Walsh LE, Gaffney MT, Pope TW. Optimising Vine Weevil, Otiorhynchus sulcatus F. (Coleoptera: Curculionidae), Monitoring Tool Design. Insects. 2022; 13(1):80. https://doi.org/10.3390/insects13010080
Chicago/Turabian StyleFezza, Eugenia, Joe M. Roberts, Toby J. A. Bruce, Lael E. Walsh, Michael T. Gaffney, and Tom W. Pope. 2022. "Optimising Vine Weevil, Otiorhynchus sulcatus F. (Coleoptera: Curculionidae), Monitoring Tool Design" Insects 13, no. 1: 80. https://doi.org/10.3390/insects13010080
APA StyleFezza, E., Roberts, J. M., Bruce, T. J. A., Walsh, L. E., Gaffney, M. T., & Pope, T. W. (2022). Optimising Vine Weevil, Otiorhynchus sulcatus F. (Coleoptera: Curculionidae), Monitoring Tool Design. Insects, 13(1), 80. https://doi.org/10.3390/insects13010080