Population Genetics of the Blueberry Gall Midge, Dasineura oxycoccana (Diptera: Cecidomyiidae), on Blueberry and Cranberry and Testing Invasion Scenarios




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
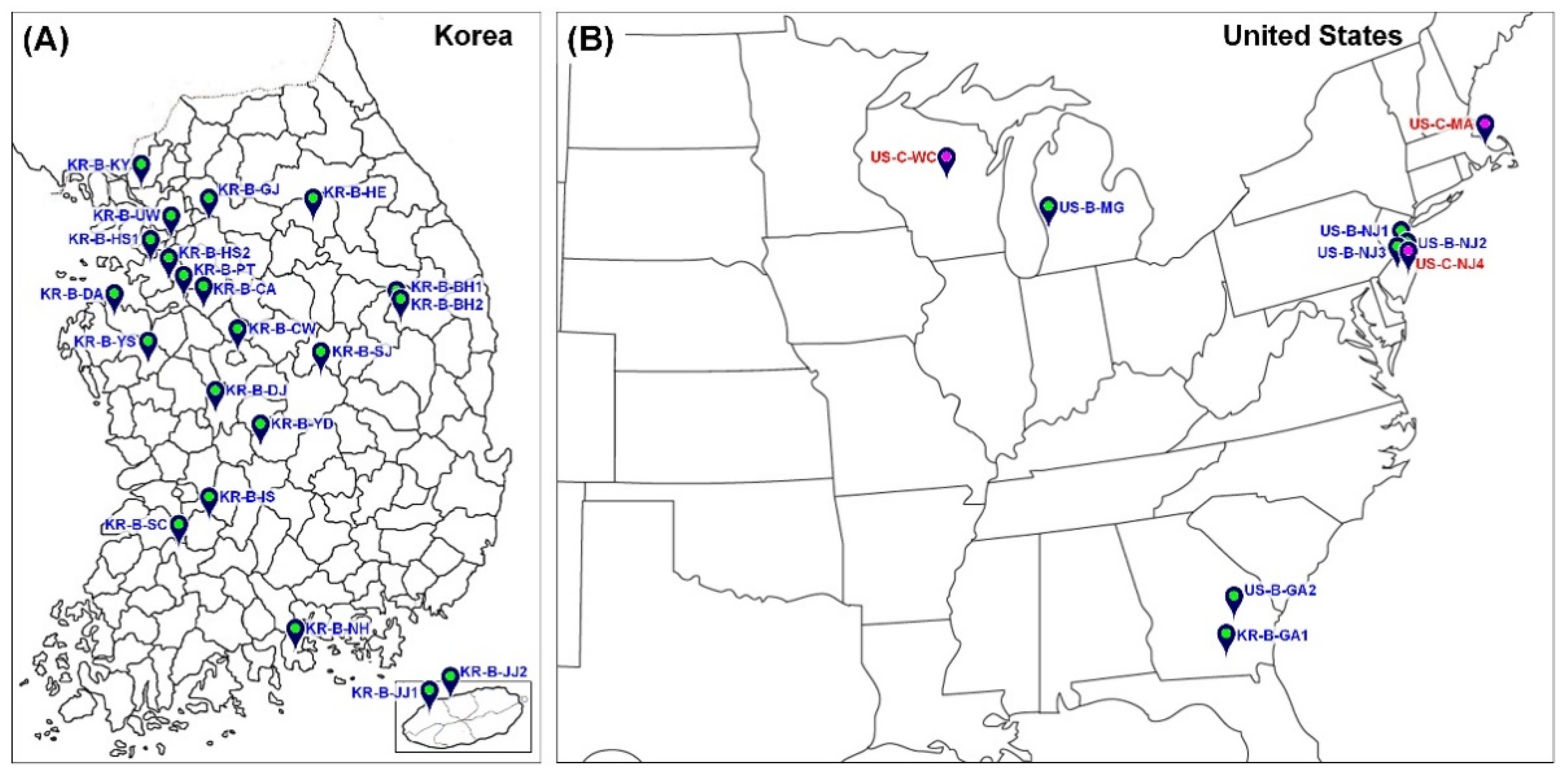
2.1. Collection Sites
2.2. Microsatellite Genotyping
2.3. Data Analyses
2.4. ABC Analysis
3. Results
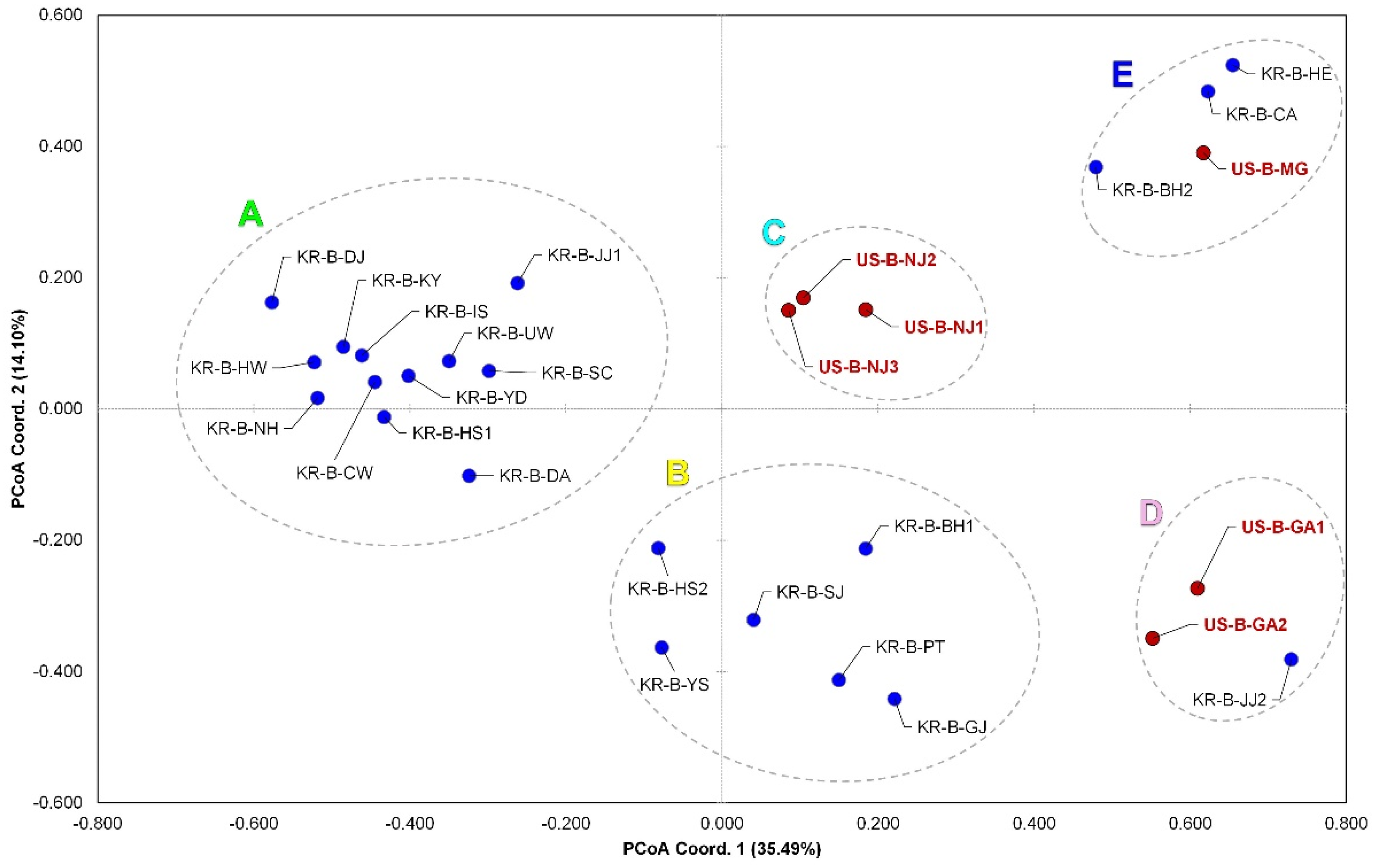
3.1. Population Genetics Analyses
3.2. Inferring Introductions from Source to Invaded Regions
4. Discussion
4.1. Ecological Speciation between the Two Host Races in Blueberry and Cranberry
4.2. Genetic Structure and Fragmentation within D. oxycoccana
4.3. Inferring Introductions from Source to Invaded Regions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lyrene, P.M.; Payne, J.A. Blueberry gall midge; a pest on rabbiteye blueberry in Florida. Proc. Fla. State Hort. Soc. 1992, 105, 297–300. [Google Scholar]
- Cook, M.A.; Fitzpatrick, S.M.; Roitberg, B.D. Phenology of Dasineura oxycoccana (Diptera: Cecidomyiidae) on cranberry and blueberry indicates potential for gene flow. J. Econ. Entomol. 2012, 105, 1205–1213. [Google Scholar] [CrossRef]
- Roubos, C.R.; Liburd, O.E. Evaluation of emergence traps for monitoring blueberry gall midge (Diptera: Cecidomyiidae) adults and within field distribution of midge infestation. J. Econ. Entomol. 2010, 103, 1258–1267. [Google Scholar] [CrossRef]
- Roubos, C.R.; Liburd, O.E. Pupation and Emergence of Blueberry Gall Midge, Dasineura oxycoccana (Diptera: Cecidomyiidae), Under Varying Temperature Conditions. Fla. Entomol. 2010, 93, 283–290. [Google Scholar] [CrossRef]
- Collins, J.A.; Drummond, F.A. The Blueberry Gall Midge (Diptera: Cecidomyiidae): A Recent Pest of Wild Blueberry (Vaccinium angustifolium; Ericales: Ericaceae) and Its Impact on Potential Yield. J. Econ. Entomol. 2019, 112, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Q. Blueberry Gall Midge A Possible New Pest in the Northwest. Or. State Univ. Ext. Serv. Fact Sheet 2005, EM8889, 1–4. [Google Scholar]
- EPPO. EPPO Global Database; EPPO: Paris, France, 2021; Available online: https://gd.eppo.int/ (accessed on 11 January 2022).
- Reekie, M.; Mackenzie, K.; Lees, B. The biology and pest potential of cranberry tipworm (Diptera, Cecidomyiidae) on lowbush blueberry. Acta Hortic. 2009, 810, 401–410. [Google Scholar] [CrossRef]
- Mathur, S.; Cook, M.A.; Sinclair, B.J.; Fitzpatrick, S.M. DNA Barcodes Suggest Cryptic Speciation in Dasineura Oxycoccana (Diptera: Cecidomyiidae) on Cranberry, Vaccinium Macrocarpon, and Blueberry, V. Corymbosum. Fla. Entomol. 2012, 95, 387–394. [Google Scholar] [CrossRef]
- Lim, J.-R.; Kim, E.-J.; Moon, H.-C.; Cho, C.-H.; Han, S.-G.; Kim, H.-J.; Song, Y.-J. Patterns of Insect Pest Occurrences and Dasineura oxycoccana Johnson in Blueberry Farms in Jeonbuk Province. Korean J. Appl. Entomol. 2016, 55, 45–51. [Google Scholar] [CrossRef]
- Sampson, B.J.; Stringer, S.J.; Spiers, J.m. Integrated Pest Management for Dasineura oxycoccana (Diptera: Cecidomyiidae) in Blueberry. Environ. Entomol. 2002, 31, 339–347. [Google Scholar] [CrossRef]
- Collins, D.W.; Harris, K.M.; Glover, R.H. Dasineura oxycoccana (Johnson, 1899) (Diptera, Cecidomyiidae), a pest of cultivated blueberres, new to Britain. Dipter. Dig. 2010, 17, 53–59. [Google Scholar]
- Hahn, N.G.; Isaacs, R. Distribution and phenology of Dasineura oxycoccana (Diptera: Cecidomyiidae) in Michigan blueberries. Environ. Entomol. 2012, 41, 455–462. [Google Scholar] [CrossRef]
- Dernisky, A.K.; Evans, R.C.; Liburd, O.E.; MacKenzie, K. Characterization of early floral damage by cranberry tipworm (Dasineura oxycoccana Johnson) as a precursor to reduced fruit set in rabbiteye blueberry (Vaccinium ashei Reade). Int. J. Pest Manag. 2005, 51, 143–148. [Google Scholar] [CrossRef]
- Sampson, B.J.; Rinehart, T.A.; Liburd, O.E.; Stringer, S.J.; Spiers, J.M. Biology of Parasitoids (Hymenoptera) Attacking Dasineura oxycoccana and Prodiplosis vaccinii (Diptera: Cecidomyiidae) in Cultivated Blueberries. Ann. Entomol. Soc. Am. 2006, 99, 113–120. [Google Scholar] [CrossRef]
- Fitzpatrick, S.M.; Gries, R.; Khaskin, G.; Peach, D.A.H.; Iwanski, J.; Gries, G. Populations of the Gall Midge Dasineura oxycoccana on Cranberry and Blueberry Produce and Respond to Different Sex Pheromones. J. Chem. Ecol. 2013, 39, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.A.; Ozeroff, S.N.; Fitzpatrick, S.M.; Roitberg, B.D. Host-associated differentiation in reproductive behaviour of cecidomyiid midges on cranberry and blueberry. Entomol. Exp. Appl. 2011, 141, 8–14. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.; Vincent, C.; Isaacs, R. Blueberry IPM: Past Successes and Future Challenges. Annu. Rev. Entomol. 2019, 64, 95–114. [Google Scholar] [CrossRef]
- Toledo Hernández, R.A.; Mikery, O.; Ibañez, S.; Aguilar, I.; Sánchez, D.; Rodríguez, D. First Record of Invasive Pest Blueberry Gall Midge, Dasineura oxycoccana (Johnson) in Mexico: Molecular and Morphological Confirmation. Southwest. Entomol. 2021, 46, 963–970. [Google Scholar] [CrossRef]
- Bosio, G.; Bogetti, C.; Brussino, G.; Gremo, F.; Scarpelli, F. Dasineura oxycoccana, a new pest of highbush blueberry in Italy. Inf. Fitopatol. 1998, 11, 36–41. [Google Scholar]
- Prodorutti, D.; Pertot, I.; Giongo, L.; Gessler, C. Highbush Blueberry: Cultivation, Protection, Breeding and Biotechnoligy. Eur. J. Plant Sci. Biotechnol. 2007, 1, 44–56. [Google Scholar]
- CABI. Invasive Species Compendium; CAB International: Wallingford, UK, 2022; Available online: https://www.cabi.org/isc (accessed on 11 January 2022).
- Yoshida, K.; Elsayed, A.K.; Matsuura, H.; Horigome, M.; Aizawa, T.; Tokuda, M. Occurrence of Dasineura oxycoccana (Diptera: Cecidomyiidae) infesting cultivated blueberry in Japan. Jpn. J. Appl. Entomol. Zool. 2017, 61, 25–28. [Google Scholar] [CrossRef]
- Kang, T.-J.; Cho, M.R.; Ahn, S.-J.; Kim, H.-H.; Kim, S.J.; Lee, S.-C.; Lee, H.-S. Monitoring and Environment-friendly Management of Blueberry Gall Midge, Dasineura oxycoccana (Diptera: Cecidomyiidae), on Blueberry. Korean J. Org. Agric. 2012, 20, 607–618. [Google Scholar] [CrossRef]
- Kim, H.; Rodriguez-Saona, C.; Kwon, D.H.; Park, S.; Kang, T.-J.; Kim, S.-J.; Hong, K.-J.; Lee, H.-S. Development and characterization of 12 microsatellite loci from the blueberry gall midge Dasineura oxycoccana (Diptera: Cecidomyiidae). Appl. Entomol. Zool. 2015, 50, 415–418. [Google Scholar] [CrossRef]
- MAFRA. Ministry of Agriculture, Food and Rural Affairs; Agricultural Business Registration Information Service: Sejong, Korea, 2022; Available online: https://edu.agrix.go.kr/uni_docs7/biOlap/dashBoard.do (accessed on 11 January 2022).
- Steck, G.J.; Lyrene, P.M.; Payne, J.A. Blueberry Gall Midge, Dasineura oxycoccana (Johnson) (Insecta: Diptera: Cecidomyiidae): EENY136/IN293, 6/2000. EDIS 2000, 2002, 1–3. [Google Scholar] [CrossRef]
- Torchin, M.E.; Lafferty, K.D.; Dobson, A.P.; McKenzie, V.J.; Kuris, A.M. Introduced species and their missing parasites. Nature 2003, 421, 628–630. [Google Scholar] [CrossRef]
- Lombaert, E.; Guillemaud, T.; Cornuet, J.-M.; Malausa, T.; Facon, B.; Estoup, A. Bridgehead effect in the worldwide invasion of the biocontrol harlequin ladybird. PLoS ONE 2010, 5, e9743. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.; Kim, S.; Lee, Y.; Lee, H.-S.; Lee, S.-J.; Choi, D.-S.; Jeon, J.; Lee, J.-H. Population Genetics for Inferring Introduction Sources of the Oriental Fruit Fly, Bactrocera dorsalis: A Test for Quarantine Use in Korea. Insects 2021, 12, 851. [Google Scholar] [CrossRef] [PubMed]
- APQA. Animal and Plant Quarantine Agency, Republic of Korea, Plant Quarantine Online Complaint System. 2022. Available online: https://okminwon.pqis.go.kr/minwon/information/statistics.html (accessed on 14 January 2022).
- Miller, N.; Estoup, A.; Toepfer, S.; Bourguet, D.; Lapchin, L.; Derridj, S.; Kim, K.S.; Reynaud, P.; Furlan, L.; Guillemaud, T. Multiple transatlantic introductions of the western corn rootworm. Science 2005, 310, 992. [Google Scholar] [CrossRef] [PubMed]
- Behura, S.K. Molecular marker systems in insects: Current trends and future avenues. Mol. Ecol. 2006, 15, 3087–3113. [Google Scholar] [CrossRef]
- Ascunce, M.S.; Yang, C.-C.; Oakey, J.; Calcaterra, L.; Wu, W.-J.; Shih, C.-J.; Goudet, J.; Ross, K.G.; Shoemaker, D. Global Invasion History of the Fire Ant Solenopsis invicta. Science 2011, 331, 1066–1068. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Arnaud-Haond, S.; Belkhir, K. GENCLONE: A computer program to analyse genotypic data, test for clonality and describe spatial clonal organization. Mol. Ecol. Notes 2007, 7, 15–17. [Google Scholar] [CrossRef]
- Dorken, M.E.; Eckert, C.G. Severely reduced sexual reproduction in northern populations of a clonal plant, Decodonverticillatus (Lythraceae). J. Ecol. 2001, 89, 339–350. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (Version-1.2)—Population-Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rice, W.R. Analyzing Tables of Statistical Tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Brookfield, J.F.Y. A simple new method for estimating null allele frequency from heterozygote deficiency. Mol. Ecol. 1996, 5, 453–455. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT, A Program to Estimate and Test Gene Diversities and Fixation Indices (version 2.9.3.2); Institute of Ecology, University of Lausanne: Lausanne, Switzerland, 2002; Available online: https://www.unil.ch/izea/softwares/fstat.html (accessed on 11 January 2022).
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.-M. Computer note. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Luikart, G. Description and Power Analysis of Two Tests for Detecting Recent Population Bottlenecks From Allele Frequency Data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E.; Huff, D.R. Evolutionary implications of allozyme and RAPD variation in diploid populations of dioecious buffalograss Buchloë dactyloides. Mol. Ecol. 1995, 4, 135–148. [Google Scholar] [CrossRef]
- Smouse, P.E.; Peakall, R. Spatial autocorrelation analysis of individual multiallele and multilocus genetic structure. Heredity 1999, 82, 561–573. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Piry, S.; Alapetite, A.; Cornuet, J.-M.; Paetkau, D.; Baudouin, L.; Estoup, A. GENECLASS2: A software for genetic assignment and first-generation migrant detection. J. Hered. 2004, 95, 536–539. [Google Scholar] [CrossRef]
- Rannala, B.; Mountain, J.L. Detecting immigration by using multilocus genotypes. Proc. Natl. Acad. Sci. USA 1997, 94, 9197–9201. [Google Scholar] [CrossRef]
- Paetkau, D.; Slade, R.; Burden, M.; Estoup, A. Genetic assignment methods for the direct, real-time estimation of migration rate: A simulation-based exploration of accuracy and power. Mol. Ecol. 2004, 13, 55–65. [Google Scholar] [CrossRef]
- Cornuet, J.-M.; Pudlo, P.; Veyssier, J.; Dehne-Garcia, A.; Gautier, M.; Leblois, R.; Marin, J.-M.; Estoup, A. DIYABCv2.0: A software to make Approximate Bayesian Computation inferences about population history using Single Nucleotide Polymorphism, DNA sequence and microsatellite data. Bioinformatics 2014, 30, 1187–1189. [Google Scholar] [CrossRef] [PubMed]
- Estoup, A.; Guillemaud, T. Reconstructing routes of invasion using genetic data: Why, how and so what? Mol. Ecol. 2010, 19, 4113–4130. [Google Scholar] [CrossRef] [PubMed]
- Cornuet, J.-M.; Ravigné, V.; Estoup, A. Inference on population history and model checking using DNA sequence and microsatellite data with the software DIYABC (v1.0). BMC Bioinform. 2010, 11, 401. [Google Scholar] [CrossRef] [PubMed]
- Cornuet, J.-M.; Santos, F.; Beaumont, M.A.; Robert, C.P.; Marin, J.-M.; Balding, D.J.; Guillemaud, T.; Estoup, A. Inferring population history with DIY ABC: A user-friendly approach to approximate Bayesian computation. Bioinformatics 2008, 24, 2713–2719. [Google Scholar] [CrossRef] [Green Version]
- Delmotte, F.; Sabater-Muñoz, B.; Prunier-Leterme, N.; Latorre, A.; Sunnucks, P.; Rispe, C.; Simon, J.-C. Phylogenetic evidence for hybrid origins of asexual lineages in an aphid species. Evolution 2003, 57, 1291–1303. [Google Scholar] [CrossRef]
- Reichel, K.; Masson, J.-P.; Malrieu, F.; Arnaud-Haond, S.; Stoeckel, S. Rare sex or out of reach equilibrium? The dynamics of FIS in partially clonal organisms. BMC Genet. 2016, 17, 76. [Google Scholar] [CrossRef]
- Guillemaud, T.; Beaumont, M.A.; Ciosi, M.; Cornuet, J.-M.; Estoup, A. Inferring introduction routes of invasive species using approximate Bayesian computation on microsatellite data. Heredity 2010, 104, 88–99. [Google Scholar] [CrossRef]
- Schlautman, B.; Covarrubias-Pazaran, G.; Fajardo, D.; Steffan, S.; Zalapa, J. Discriminating power of microsatellites in cranberry organelles for taxonomic studies in Vaccinium and Ericaceae. Genet. Resour. Crop Evol. 2017, 64, 451–466. [Google Scholar] [CrossRef]
- Walsh, B.D. On phytophagic varieties and phytophagic species. Proc. Entomol. Soc. Phila. 1864, 3, 403–430. [Google Scholar]
- Matsubayashi, K.W.; Ohshima, I.; Nosil, P. Ecological speciation in phytophagous insects. Entomol. Exp. Appl. 2010, 134, 1–27. [Google Scholar] [CrossRef]
- Dieckmann, U.; Doebeli, M. Adaptive dynamics of speciation: Sexual populations. In Adaptive Speciation; Dieckmann, U., Doebeli, M., Metz, J.A.J., Tautz, D., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 76–111. [Google Scholar]
- Via, S. Reproductive isolation between sympatric races of pea aphids. I. Gene flow restriction and habitat choice. Evolution 1999, 53, 1446–1457. [Google Scholar] [CrossRef] [PubMed]
- Via, S.; Bouck, A.C.; Skillman, S. Reproductive isolation between divergent races of pea aphids on two hosts. II. Selection against migrants and hybrids in the parental environments. Evolution 2000, 54, 1626–1637. [Google Scholar] [CrossRef]
- Hawthorne, D.J.; Via, S. Genetic linkage of ecological specialization and reproductive isolation in pea aphids. Nature 2001, 412, 904–907. [Google Scholar] [CrossRef]
- Via, S.; Hawthorne, D.J. The Genetic Architecture of Ecological Specialization: Correlated Gene Effects on Host Use and Habitat Choice in Pea Aphids. Am. Nat. 2002, 159, S76–S88. [Google Scholar] [CrossRef]
- Medina, R.F.; Dickey, A.M.; Harrison, K.; Miller, G.L. Host-associated differentiation in a pecan and water hickory Aphidomorpha community. Entomol. Exp. Appl. 2017, 162, 366–378. [Google Scholar] [CrossRef]
- Medina, R.F.; Szendrei, Z.; Harrison, K.; Isaacs, R.; Averill, A.; Malo, E.A.; Rodriguez-Saona, C. Exploring host-associated differentiation in the North American native cranberry fruitworm, Acrobasis vaccinii, from blueberries and cranberries. Entomol. Exp. Appl. 2014, 150, 136–148. [Google Scholar] [CrossRef]
- Fitzpatrick, S.M. Insect life histories in fruit, shoot and root environments of cranberry and blueberry. Acta Hortic. 2009, 810, 231–250. [Google Scholar] [CrossRef]
- Coville, F.V. Improving the wild blueberry. In Yearbook of the United States Department of Agriculture; United States Department of Agriculture: Washington, DC, USA, 1937; pp. 559–574. [Google Scholar]
- Schlautman, B.; Diaz-Garcia, L.; Covarrubias-Pazaran, G.; Schlautman, N.; Vorsa, N.; Polashock, J.; Ogden, E.L.; Brown, A.; Lin, Y.-C.; Bassil, N.; et al. Comparative genetic mapping reveals synteny and collinearity between the American cranberry and diploid blueberry genomes. Mol. Breed. 2018, 38, 9. [Google Scholar] [CrossRef]
- Roubos, C.R.; Isaacs, R. Blueberry Gall Midge Dasineura oxycoccana (Johnson) (Diptera: Cecidomyiidae). Mich. State Univ. Ext. Bull. 2013, E3191, 1–4. [Google Scholar]
- Hulme, P.E. Unwelcome exchange: International trade as a direct and indirect driver of biological invasions worldwide. One Earth 2021, 4, 666–679. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Kim, H.; Hoelmer, K.A.; Lee, S. Population genetics of the soybean aphid in North America and East Asia: Test for introduction between native and introduced populations. Biol. Invasions 2017, 19, 597–614. [Google Scholar] [CrossRef]
- Carter, M.; Smith, M.; Harrison, R. Genetic analyses of the Asian longhorned beetle (Coleoptera, Cerambycidae, Anoplophora glabripennis), in North America, Europe and Asia. Biol. Invasions 2010, 12, 1165–1182. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.; Lee, Y.; Lee, H.-S.; Lee, S.-J.; Lee, J.-H. Tracing the Origin of Korean Invasive Populations of the Spotted Lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae). Insects 2021, 12, 539. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pop. ID | No. | Ho (SD) | He (SD) | HWE | HS | NA | RS | FIS |
|---|---|---|---|---|---|---|---|---|
| KR-B-UW | 20 | 0.788 (0.051) | 0.740 (0.024) | Ns | 0.74 | 6.83 | 1.74 | −0.07 |
| KR-B-GJ | 17 | 0.692 (0.068) | 0.727 (0.024) | Ns | 0.73 | 6.17 | 1.73 | 0.06 |
| KR-B-HS1 | 23 | 0.803 (0.052) | 0.698 (0.018) | Ns | 0.70 | 6.75 | 1.70 | −0.15 |
| KR-B-HS2 | 12 | 0.788 (0.051) | 0.740 (0.024) | Ns | 0.70 | 6.69 | 1.72 | −0.13 |
| KR-B-KY | 20 | 0.792 (0.059) | 0.686 (0.026) | Ns | 0.68 | 5.25 | 1.69 | −0.16 |
| KR-B-PT | 2 | 0.708 (0.114) | 0.653 (0.075) | Ns | 0.68 | 2.42 | 1.65 | −0.13 |
| KR-B-HE | 18 | 0.880 (0.032) | 0.697 (0.024) | * excess | 0.69 | 5.75 | 1.70 | −0.27 |
| KR-B-CW | 20 | 0.817 (0.059) | 0.706 (0.022) | Ns | 0.70 | 6.00 | 1.71 | −0.16 |
| KR-B-YD | 39 | 0.738 (0.052) | 0.705 (0.017) | Ns | 0.70 | 8.08 | 1.70 | −0.05 |
| KR-B-DA | 15 | 0.769 (0.061) | 0.699 (0.022) | Ns | 0.70 | 5.25 | 1.70 | −0.10 |
| KR-B-CA | 20 | 0.821 (0.042) | 0.732 (0.023) | Ns | 0.73 | 6.17 | 1.73 | −0.13 |
| KR-B-DJ | 9 | 0.898 (0.040) | 0.619 (0.027) | * excess | 0.60 | 3.67 | 1.62 | −0.49 |
| KR-B-YS | 8 | 0.891 (0.035) | 0.719 (0.040) | Ns | 0.68 | 4.50 | 1.72 | −0.30 |
| KR-B-IS | 20 | 0.721 (0.064) | 0.701 (0.026) | Ns | 0.70 | 6.17 | 1.70 | −0.03 |
| KR-B-SC | 20 | 0.809 (0.049) | 0.721 (0.025) | Ns | 0.72 | 6.67 | 1.72 | −0.13 |
| KR-B-HW | 20 | 0.713 (0.072) | 0.649 (0.033) | Ns | 0.65 | 3.92 | 1.65 | −0.10 |
| KR-B-BH1 | 13 | 0.788 (0.062) | 0.699 (0.051) | Ns | 0.69 | 6.08 | 1.70 | −0.14 |
| KR-B-BH2 | 7 | 0.833 (0.060) | 0.782 (0.026) | Ns | 0.78 | 5.33 | 1.78 | −0.07 |
| KR-B-SJ | 31 | 0.805 (0.049) | 0.734 (0.029) | Ns | 0.73 | 7.83 | 1.73 | −0.10 |
| KR-B-NH | 20 | 0.825 (0.064) | 0.647 (0.036) | * excess | 0.64 | 4.83 | 1.65 | −0.29 |
| KR-B-JJ1 | 19 | 0.626 (0.081) | 0.720 (0.061) | * deficit | 0.72 | 7.08 | 1.72 | 0.13 |
| KR-B-JJ2 | 11 | 0.659 (0.077) | 0.714 (0.028) | Ns | 0.72 | 5.58 | 1.71 | 0.08 |
| US-B-GA1 | 8 | 0.762 (0.072) | 0.729 (0.028) | Ns | 0.73 | 4.58 | 1.73 | −0.05 |
| US-B-GA2 | 9 | 0.731 (0.059) | 0.689 (0.038) | Ns | 0.69 | 5.08 | 1.69 | −0.07 |
| US-B-NJ1 | 4 | 0.681 (0.069) | 0.788 (0.054) | Ns | 0.81 | 4.42 | 1.79 | 0.16 |
| US-B-NJ2 | 32 | 0.784 (0.048) | 0.779 (0.030) | Ns | 0.78 | 11.17 | 1.78 | −0.01 |
| US-B-NJ3 | 35 | 0.848 (0.046) | 0.789 (0.019) | Ns | 0.79 | 9.92 | 1.79 | −0.08 |
| US-B-MG | 40 | 0.759 (0.077) | 0.776 (0.033) | * deficit | 0.78 | 11.08 | 1.78 | 0.02 |
| US-C-NJ4 | 40 | 0.697 (0.094) | 0.681 (0.062) | Ns | 0.68 | 10.83 | 1.68 | −0.02 |
| US-C-MA | 40 | 0.655 (0.119) | 0.596 (0.090) | Ns | 0.60 | 8.17 | 1.60 | −0.10 |
| US-C-WC | 40 | 0.617 (0.119) | 0.538 (0.084) | Ns | 0.54 | 7.42 | 1.54 | −0.15 |
| Case | Among Groups | Among Populations within Groups | Within Populations | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Va | PV | p | Vb | PV | p | Vc | PV | p | |
| 1 | 0.82 | 14.58 | <0.0001 | 0.59 | 10.62 | <0.0001 | 4.18 | 74.80 | <0.0001 |
| 2 * | 0.38 | 7.49 | <0.0001 | 0.42 | 8.34 | <0.0001 | 4.31 | 84.17 | <0.0001 |
| 3 * | 0.54 | 10.65 | <0.0001 | 0.22 | 4.32 | <0.0001 | 4.31 | 85.03 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Rodriguez-Saona, C.; Lee, H.-S. Population Genetics of the Blueberry Gall Midge, Dasineura oxycoccana (Diptera: Cecidomyiidae), on Blueberry and Cranberry and Testing Invasion Scenarios. Insects 2022, 13, 880. https://doi.org/10.3390/insects13100880
Kim H, Rodriguez-Saona C, Lee H-S. Population Genetics of the Blueberry Gall Midge, Dasineura oxycoccana (Diptera: Cecidomyiidae), on Blueberry and Cranberry and Testing Invasion Scenarios. Insects. 2022; 13(10):880. https://doi.org/10.3390/insects13100880
Chicago/Turabian StyleKim, Hyojoong, Cesar Rodriguez-Saona, and Heung-Sik Lee. 2022. "Population Genetics of the Blueberry Gall Midge, Dasineura oxycoccana (Diptera: Cecidomyiidae), on Blueberry and Cranberry and Testing Invasion Scenarios" Insects 13, no. 10: 880. https://doi.org/10.3390/insects13100880
APA StyleKim, H., Rodriguez-Saona, C., & Lee, H. -S. (2022). Population Genetics of the Blueberry Gall Midge, Dasineura oxycoccana (Diptera: Cecidomyiidae), on Blueberry and Cranberry and Testing Invasion Scenarios. Insects, 13(10), 880. https://doi.org/10.3390/insects13100880