
Insights into the Antennal Characteristics and Olfactory Strategy of the Endangered Rhino Stomach Bot Fly Gyrostigma rhinocerontis (Diptera: Oestridae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition of Samples
2.2. Morphological Analyses under Light and Scanning Electron Microscopy
3. Results
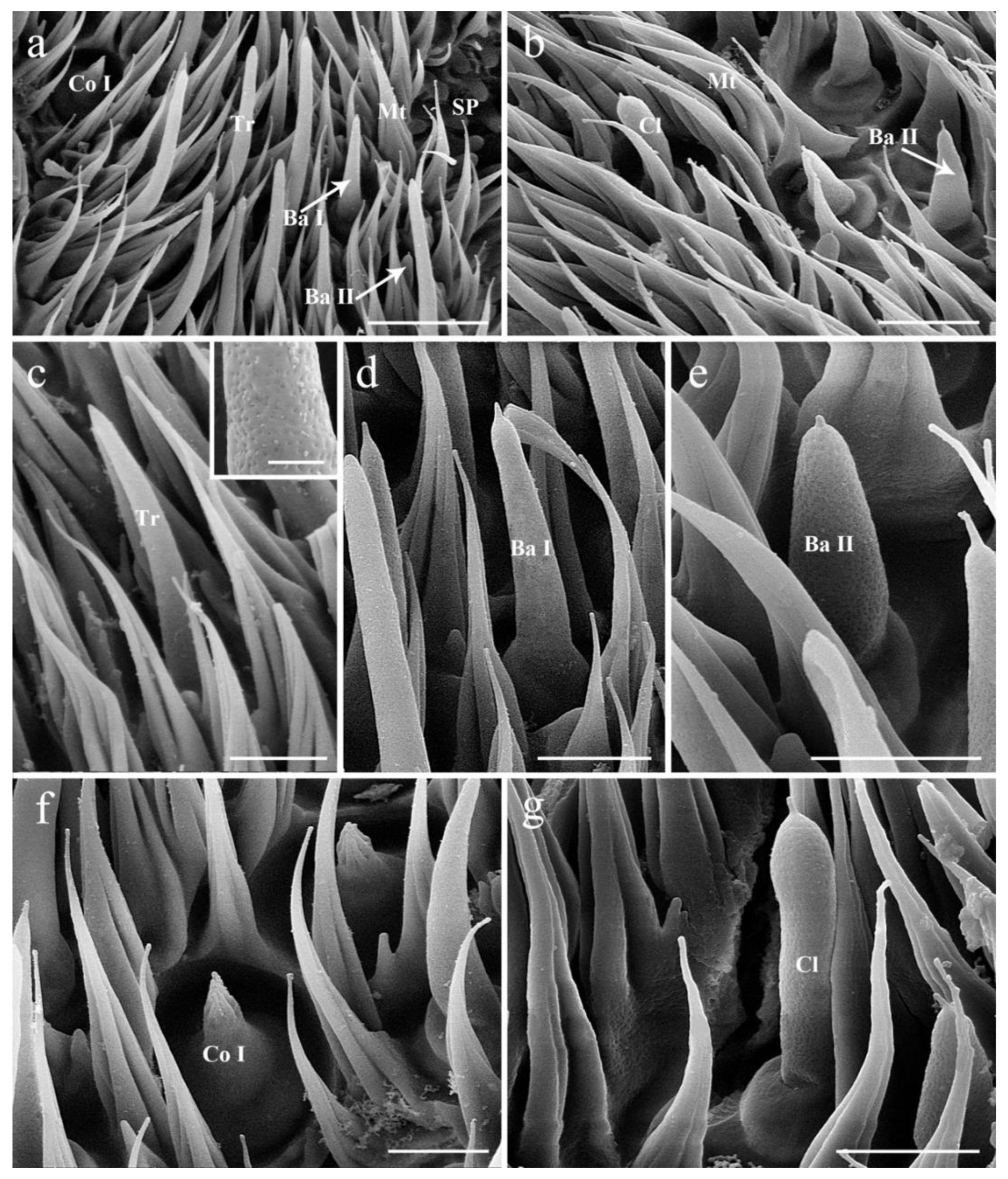
3.1. General Description of the Antenna in G. rhinocerontis
3.2. Antennal Scape and Pedicel
3.3. Antennal Postpedicel
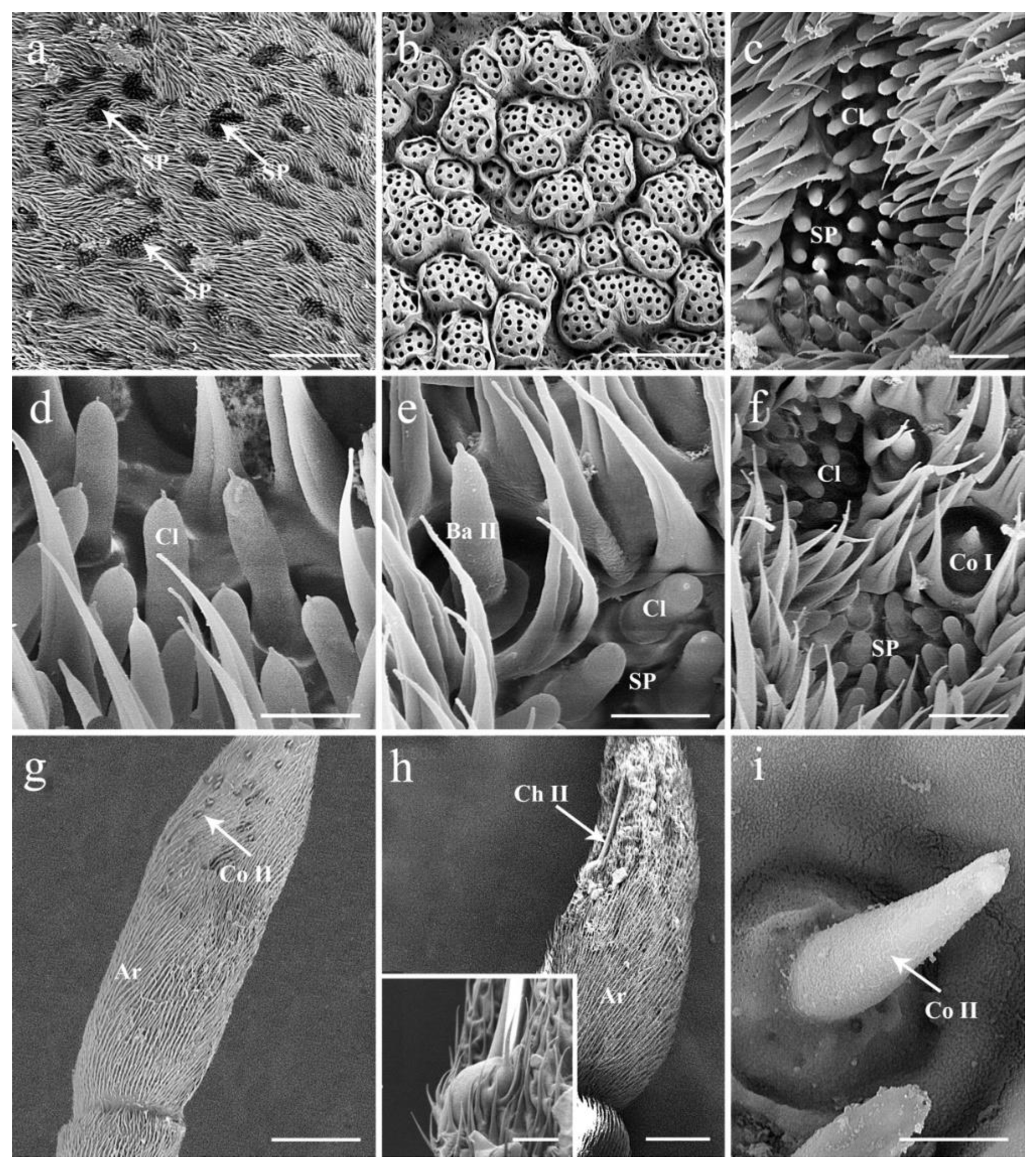
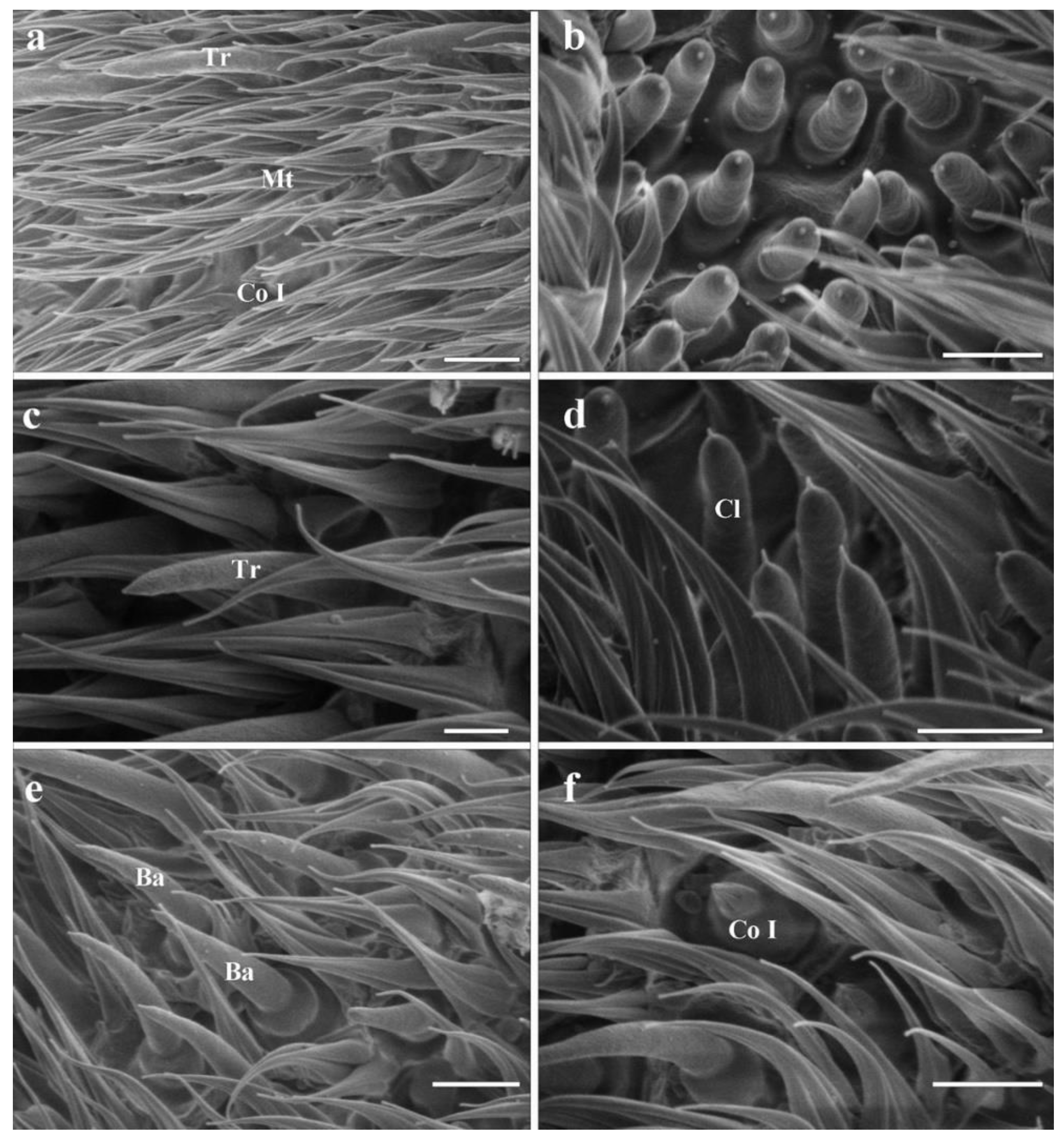
3.4. General Description of the Sensilla on Postpedicel
3.4.1. Trichoid Sensilla
3.4.2. Basiconic Sensilla
3.4.3. Coeloconic Sensilla
3.4.4. Clavate Sensilla
3.4.5. Sensory Pits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, M.J.; Wall, R. Myiasis of Humans and Domestic Animals. Adv. Parasitol. 1995, 35, 257–334. [Google Scholar] [CrossRef]
- Robbins, K.; Khachemoune, A. Cutaneous myiasis: A review of the common types of myiasis. Int. J. Dermatol. 2010, 49, 1092–1098. [Google Scholar] [CrossRef]
- James, M.T. The flies that cause myiasis in man. Misc. Publ. US Depart. Agric. 1947, 631, 1–175. [Google Scholar]
- Zumpt, F. Myiasis in Man and Animals of the Old World; Butterworths: London, UK, 1965; pp. 1–264. [Google Scholar]
- Grunin, K.J. 64b; Hypodermatidae. In Die Fliegen der PalaÈarktischen Region, 8; Lindner, E., Ed.; Schweizerbart’sche: Stuttgart, Deutschland, 1965; pp. 1–154. [Google Scholar]
- Grunin, K.J. 64a’; Oestridae. In Die Fliegen der PalaÈarktischen Region, 8; Lindner, E., Ed.; Schweizerbart’sche: Stuttgart, Deutschland, 1966; pp. 1–96. [Google Scholar]
- Grunin, K.J. 64a; Gasterophilidae. In Die Fliegen der PalaÈarktischen Region, 8; Lindner, E., Ed.; Schweizerbart’sche: Stuttgart, Deutschland, 1969; pp. 1–66. [Google Scholar]
- Papavero, N. The World Oestridae (Diptera) Mammals and Continental Drift. In Series Entomologica; Springer: Berlin/Heidelberg, Germany, 1977; Volume 14, pp. 1–240. [Google Scholar] [CrossRef]
- Colwell, D.D.; Hall, M.J.R.; Scholl, P.J. The Oestrid Flies: Biology, Host-Parasite Relationships, Impact and Management; CABI: Wallingford/Cambridge, UK, 2006; pp. 1–357. [Google Scholar] [CrossRef]
- Schneider, D.; Steinbrecht, R.A. Checklist of insect olfactory sensilla. Sym. Zool. Soc. Lond. 1968, 23, 279–297. [Google Scholar]
- Chapman, R.F. Chemoreception: The significance of receptor numbers. Adv. Insect. Phys. 1982, 16, 247–356. [Google Scholar] [CrossRef]
- Chapman, R.F.; Simpson, S.J.; Douglas, A.E. The Head, Ingestion, Utilization and Distribution of Food. In The Insects: Structure and Function, 5th ed.; Chapman, R.F., Ed.; Cambridge University Press: New York, NY, USA, 2013; pp. 3–13. [Google Scholar]
- Colwell, D.D.; Otranto, D.; Stevens, J.R. Oestrid flies: Eradication and extinction versus biodiversity. Trends. Parasitol. 2009, 25, 500–504. [Google Scholar] [CrossRef]
- Penzhorn, B.L.; Krecek, R.C.; Horak, I.G.; Verster, A.J.M.; Walker, J.B.; Boomker, J.D.F.; Knapp, S.E.; Quandt, S.K.F. Parasites of African Rhinos: A Documentation. In Proceedings of the a Symposium on Rhinos as Game Ranch Animals, Onderstepoort, South Africa, 9–10 September 1994; pp. 9–10. [Google Scholar]
- Miller, M.A.; Buss, P.E. Rhinoceridae (Rhinoceroses). In Zoo and Wild Animal Medicine, 8th ed.; Miller, R.E., Fowler, M.E., Eds.; Elsevier Health Sciences, Saunders: St. Louis, MI, USA, 2014; Volume 8, pp. 538–547. [Google Scholar]
- Pachaly, J.R.; Monteiro-filho, L.P.C.; Gonçalves, D.D.; Voltarelli-pachaly, E.M. Gyrostigma rhinocerontis (Diptera: Oestridae, Gasterophilinae) in white rhinoceroses (Ceratotherium simum) imported from South Africa: Occurrence in Itatiba, São Paulo, Brazil. Pesqui. Vet. Bras. 2016, 36, 749–752. [Google Scholar] [CrossRef]
- Kretzschmar, P.; Auld, H.; Boag, P.; Gansloßer, U.; Scott, C.; Van Coeverden de Groot, P.J.; Courtiol, A. Mate choice, reproductive success and inbreeding in white rhinoceros: New insights for conservation management. Evol. Appl. 2020, 13, 699–714. [Google Scholar] [CrossRef]
- Harper, C.; Ludwig, A.; Clarke, A.; Makgopela, K.; Yurchenko, A.; Guthrie, A.; Dobrynin, P.; Tamazian, G.; Emslie, R.; Heerden, M.; et al. Robust forensic matching of confiscated horns to individual poached African rhinoceros. Curr. Biol. 2018, 28, R13–R14. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, B.; Chu, H.; Zhang, D.; Li, K. Gasterophilus (Diptera, Gasterophilidae) infestation of equids in the Kalamaili Nature Reserve, China. Parasite 2016, 23, 36. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Li, K.; Hu, D. The incidence and species composition of Gasterophilus (Diptera, Gasterophilidae) causing equine myiasis in northern Xinjiang, China. Vet. Parasitol. 2016, 217, 36–38. [Google Scholar] [CrossRef]
- Stevens, J.R.; Wallman, J.F.; Otranto, D.; Wall, R.; Pape, T. The evolution of myiasis in humans and other animals in the old and new worlds (part ii): Biological and life-history studies. Trends. Parasitol. 2006, 22, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Birch, M.C.; Haynes, K.F. Insect Pheromones. In Studies in Biology; Edward Arnold: London, UK, 1982; pp. 1–58. [Google Scholar]
- Steinbrecht, R.A. Structure and Function of Insect Olfactory Sensilla. In Olfaction in Mosquito-Host Interactions; Bock, G.R., Cardew, G., Eds.; Wiley: London, UK, 1996; pp. 158–177. [Google Scholar]
- Hunter, F.F.; Adserballe, C.F. Cuticular structures on the antennae of Hypoderma bovis De Geer (Diptera: Oestridae) females. Int. J. Insect Morphol. Embryol. 1996, 25, 173–181. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Q.K.; Hu, D.F.; Li, K. Sensilla on the antennal funiculus of the stomach bot fly, Gasterophilus nigricornis (Diptera: Oestridae). Med. Vet. Entomol. 26, 314–322. [CrossRef]
- Zhang, D.; Li, X.Y.; Liu, X.H.; Wang, Q.K.; Pape, T. The antenna of horse stomach bot flies: Morphology and phylogenetic implications (Oestridae, Gasterophilinae: Gasterophilus Leach). Sci. Rep. 2016, 6, 34409. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Liu, X.H.; Ge, Y.Q.; Zhang, D. Scanning electron microscopy of antennal sensory organs of the cattle grub, Hypoderma lineatum (Diptera: Oestridae). Parasitol. Res. 2015, 114, 3865. [Google Scholar] [CrossRef]
- Liu, X.H.; Li, X.Y.; Li, K.; Zhang, D. Ultrastructure of antennal sensory organs of horse nasal-myiasis fly, Rhinoestrus purpureus (Diptera: Oestridae). Parasitol. Res. 2015, 114, 2527. [Google Scholar] [CrossRef]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Boonchu, N.; Chaiwong, T.; Ngern-Klun, R.; Sripakdee, D.; Vogtsberger, R.C.; Olson, J.K. Antennal sensilla of some forensically important flies in families Calliphoridae, Sarcophagidae and Muscidae. Micron 2004, 35, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Smallegange, R.C.; Kelling, R.J.; Den Otter, C.J. Types and numbers of sensilla on antennae and maxillary palps of small and large houseflies, Musca domestica (Diptera, Muscidae). Microsc. Res. Tech. 2008, 71, 880–886. [Google Scholar] [CrossRef]
- Setzu, M.D.; Poddighe, S.; Angioy, A.M. Sensilla on the antennal funiculus of the blow fly, Protophormia terraenovae (Diptera: Calliphoridae). Micron 2011, 42, 471–477. [Google Scholar] [CrossRef]
- Wang, Q.K.; Liu, X.H.; Lu, P.F.; Zhang, D. Ultrastructure of antennal sensilla in Hydrotaea armipes (Fallén) (Diptera: Muscidae): New evidence for taxonomy of the genus Hydrotaea. Zootaxa. 2014, 3790, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.K.; Yang, Y.Z.; Liu, M.Q.; Zhang, D. Fine structure of Delia platura (Meigen) (Diptera: Anthomyiidae) revealed by scanning electron microscopy. Microsc. Res. Tech. 2014, 77, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, Q.K.; Hu, D.F.; Li, K. Cuticular structures on antennae of the bot fly, Portschinskia magnifica (Diptera: Oestridae). Parasitol. Res. 2012, 111, 1651–1659. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Q.K.; Liu, X.H.; Chen, Y.O.; Li, K. Sensilla on antenna and maxillary palp of predaceous fly, Lispe neimongola Tian et Ma (Diptera: Muscidae). Micron 2013, 49, 33–39. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Q.K.; Yang, Y.Z.; Chen, Y.O.; Li, K. Sensory organ of the antenna of two Fannia species (Diptera: Fanniidae). Parasitol. Res. 2013, 112, 2177–2185. [Google Scholar] [CrossRef]
- Pezzi, M.; Whitmore, D.; Chicca, M.; Semeraro, B.; Brighi, F.; Leis, M. Ultrastructural morphology of the antenna and maxillary palp of Sarcophaga tibialis (Diptera: Sarcophagidae). J. Med. Entomol. 2016, 53, 807–814. [Google Scholar] [CrossRef]
- Pape, T. Phylogeny of Oestridae (Insecta: Diptera). Syst Entomol. 2001, 26, 133–171. [Google Scholar] [CrossRef]
- Cumming, J.M.; Wood, D.M. Adult Morphology and Terminology. In Manual of Afrotropical Diptera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2017; Volume 1, pp. 89–133. [Google Scholar]
- Liu, G.T.; Wang, Q.K.; Liu, X.H.; Li, X.Y.; Pang, X.N.; Zhang, D. Antennal and palpal sensilla of three predatory Lispe species (Diptera: Muscidae): An ultrastructural investigation. Sci. Rep. 2021, 11, 18357. [Google Scholar] [CrossRef]
- Roh, G.H.; Lee, Y.J.; Park, C.G. Morphology and distribution of antennal sensilla in a parasitoid fly, Gymnosoma rotundatum (Diptera: Tachinidae). Microsc. Res. Tech. 2020, 83, 589–596. [Google Scholar] [CrossRef]
- Abouzied, E.M. Ultrastructure of the sensory organs of male Exorista sp. (Diptera: Tachinidae) collected from Al-baha, KSA. Bulltein Entomol. Soc. Egypt 2008, 85, 13–28. [Google Scholar]
- Dwyer, R.A.; Witte, C.; Buss, P.; Goosen, W.J.; Miller, M. Epidemiology of tuberculosis in multi-host wildlife systems: Implications for Black (Diceros bicornis) and White (Ceratotherium simum) Rhinoceros. Front. Vet. Sci. 2020, 7, 580476. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M. Sensory Ecology, Behaviour, and Evolution; Oxford University Press: Oxford, UK, 2015; pp. 3–17. [Google Scholar]
- Andreani, A.; Sacchetti, P.; Belcari, A. Comparative morphology of the deer ked Lipoptena fortisetosa first recorded from Italy. Med. Vet. Entomol. 2019, 33, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Andreani, A.; Belcari, A.; Sacchetti, P.; Romani, R. Antennal morphology and fine structure of flagellar sensilla in Hippoboscid Flies with special reference to Lipoptena fortisetosa (Diptera: Hippoboscidae). Insects 2022, 13, 236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, X.H.; Li, X.Y.; Cao, J.; Chu, H.J.; Li, K. Ultrastructural investigation of antennae in three cutaneous myiasis flies: Melophagus ovinus, Hippobosca equina, and Hippobosca longipennis (Diptera: Hippoboscidae). Parasitol. Res. 2015, 114, 1887–1896. [Google Scholar] [CrossRef]
- Wang, Q.; Shang, Y.; Hilton, D.S.; Inthavong, K.; Zhang, D.; Elgar, M.A. Antennal scales improve signal detection efficiency in moths. Proc. R. Soc. B 2018, 285, 20172832. [Google Scholar] [CrossRef]
- Poddighe, S.; Dekker, T.; Scala, A.; Angioy, A.M. Olfaction in the female sheep botfly. Naturwissenschaften 2010, 97, 827–835. [Google Scholar] [CrossRef]
- Fernandes, F.D.F.; Linardi, P.M.; Chiarini-Garcia, H. Morphology of the antenna of Dermatobia hominis (Diptera: Cuterebridae) based on scanning electron microscopy. J. Med. Entomol. 2002, 39, 36–43. [Google Scholar] [CrossRef] [PubMed]
- McAlpine, D.K. Observation on antennal morphology in Diptera with particular reference to the articular surfaces between segments 2 and 3 in the Cyclorrhapha. Rec. Aust. Mus. 2011, 63, 113–166. [Google Scholar] [CrossRef] [Green Version]
- Elgar, M.A.; Zhang, D.; Wang, Q.; Wittwer, B.; Pham, H. Insect antennal morphology: Evolution of diverse solutions to a similar problem. Yale J. Med. Biol. 2018, 91, 457–469. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensilla Type | Sex | Length (μm) | Basal Diameter (μm) | Tip Diameter (μm) |
|---|---|---|---|---|
| Ch I | Female | 133.10 ± 4.28 | − | − |
| Male | − | − | − | |
| Ch II | Female | 74.67 | − | − |
| Male | 76.38 | − | − | |
| Tr | Female | 18.15 ± 1.82 | 2.14 ± 0.32 | − |
| Male | 18.05 ± 0.59 | 2.26 ± 0.15 | ||
| Ba I | Female | 11.33 ± 1.99 | 2.40 ± 0.42 | − |
| Male | 10.05 ± 0.47 | 2.46 ± 0.30 | ||
| Ba II | Female | 7.85 ± 0.61 | 2.94 ± 0.63 | − |
| Male | − | − | − | |
| Co I | Female | 2.88 ± 0.40 | 1.97 ± 0.25 | − |
| Male | 2.59 ± 0.13 | 1.92 ± 0.13 | − | |
| Co II | Female | 6.87 ± 0.58 | 2.35 ± 0.12 | − |
| Male | 6.54 ± 0.32 | 2.23 ± 0.15 | − | |
| Cl | Female | 8.29 ± 0.94 | 2.01 ± 0.14 | 2.20 ± 0.15 |
| Male | 7.92 ± 0.52 | 2.10 ± 0.30 | 2.21 ± 0.14 |
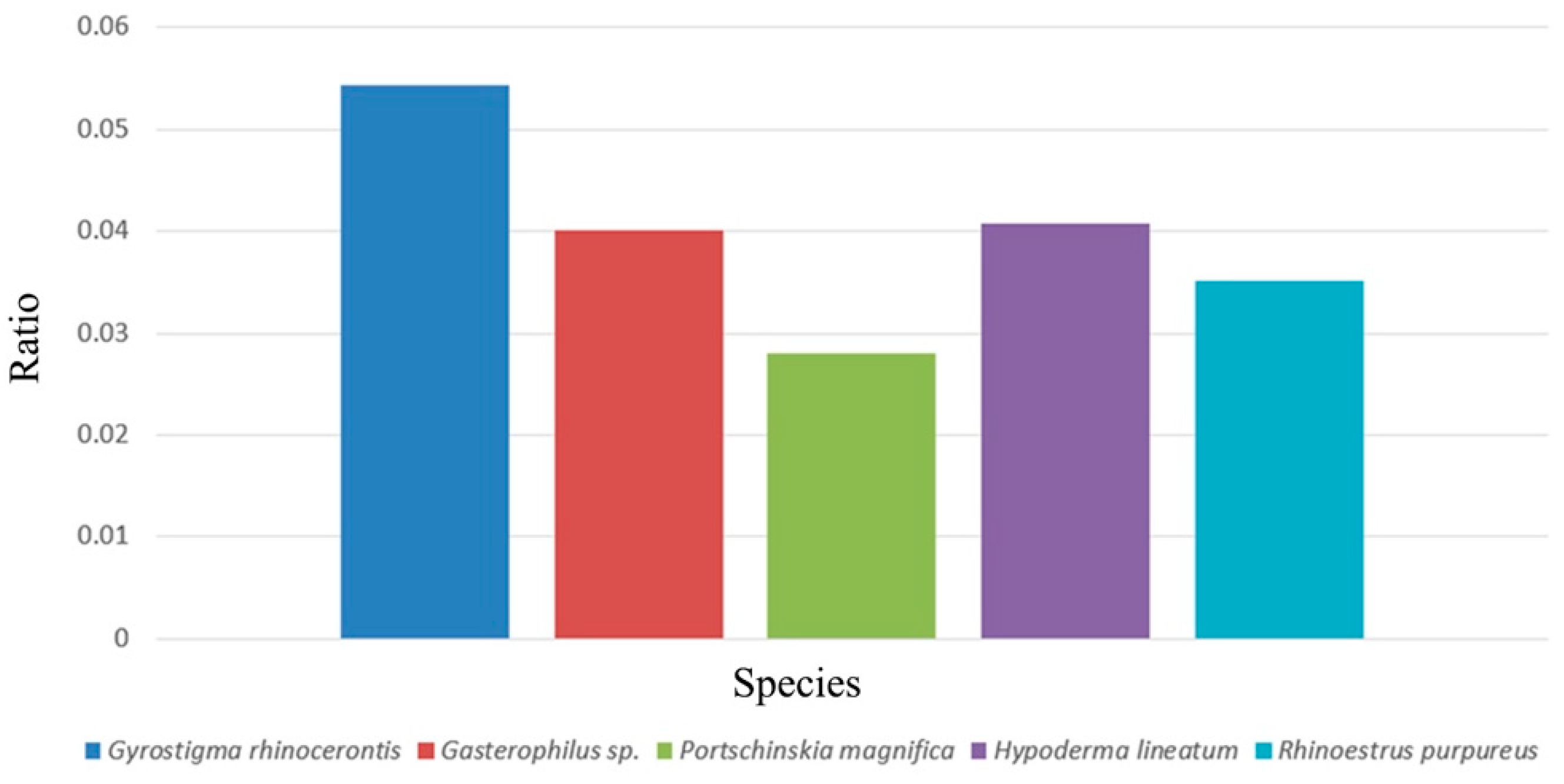
| Subfamily | Species | Pedicel Shape | Number of Sensory Pits | Sensilla Number in Each Sensory Pit | Host | Oviposition Location |
|---|---|---|---|---|---|---|
| Gasterophilinae | Gasterophilus haemorrhoidalis (Linnaeus) [9,26,38] | partly enveloping postpedicel | 60 | 2–5 (Auriculate sensilla) | Equine | Singly on hair of lips (on host) |
| Gasterophilus intestinalis (De Geer) [9,26,38] | partly enveloping postpedicel | 83 | 2–5 (Auriculate sensilla) | Equine | Hairs on forelegs and chest (on host) | |
| Gasterophilus nasalis (Linnaeus) [9,26,38] | partly enveloping postpedicel | 47 | 1 (Clavate sensilla) | Equine | Hairs under chin (on host) | |
| Gasterophilus pecorum (Fabricius) [9,26,38] | partly enveloping postpedicel | 0 | 0 | Equine | Grass or vegetation | |
| Gasterophilus nigricornis (Loew) [9,25,38] | partly enveloping postpedicel | 68 | 5–7 (Auriculate sensilla) | Equine | Hairs of cheek or neck (on host) | |
| Gyrostigma rhinocerontis (Hope) [9,26,38] | greatly enlarged and has a reduced ventral part | 500–700 | ~50 | Rhinoceros | Skin surface (on host) | |
| Cobboldia loxodontis Brauer [9,38] | partly enveloping the postpedicel | unknown | unknown | Elephants | Base of tusk (on host) | |
| Oestrinae | Oestrus ovis Linnaeus [9,38,49] | partly enveloping the postpedicel | 50 | 3 (Basiconica sensilla) | Ovine | All Oestrinae are larviparous; forcefully inject their progeny on to host |
| Rhinoestrus purpureus (Brauer) [9,28,38] | partly enveloping the postpedicel | 37 | 2–8 (Clavate sensilla; basiconic sensilla) | Equine | (Same as above) | |
| Hypodermatinae | Hypoderma bovis (Linnaeus) [9,24,38] | surrounding and enclosing base of postpedicel | >300 | <6 (Basiconica sensilla) | Bovine | Hairs of host |
| Hypoderma lineatum (Villers) [9,27,38] | enlarged and encircles nearly entire postpedicel | 30 | 2–6 (Basiconic sensilla; Trichoid sensilla) | Bovine | Hairs of host | |
| Oestromyia leporina (Pallas) [9,38], unpublished data | partly enveloping postpedicel | 25 | 2–5 | Pikas | Hairs of host | |
| Portschinskia magnifica Pleske [9,34,38] | hollowed pedicel entirely enclosing postpedicel | 0 | - | Mice | unknown | |
| Cuterebrinae | Dermatobia hominis (Linnaeus Jr.) [8,38,50] | Normal shape as in other calyptrate flies | ~3–5 | 1 (Coeloconic sensilla) | Non-specific; larger mammals and birds | Egg attached to different species of blood-feeding insects captured by female bot fly. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Li, X.; Wang, Q.; Zhang, C.; Yang, M.; Zhou, T.; Li, K.; Zhang, D. Insights into the Antennal Characteristics and Olfactory Strategy of the Endangered Rhino Stomach Bot Fly Gyrostigma rhinocerontis (Diptera: Oestridae). Insects 2022, 13, 889. https://doi.org/10.3390/insects13100889
Xu W, Li X, Wang Q, Zhang C, Yang M, Zhou T, Li K, Zhang D. Insights into the Antennal Characteristics and Olfactory Strategy of the Endangered Rhino Stomach Bot Fly Gyrostigma rhinocerontis (Diptera: Oestridae). Insects. 2022; 13(10):889. https://doi.org/10.3390/insects13100889
Chicago/Turabian StyleXu, Wentian, Xinyu Li, Qike Wang, Chenglin Zhang, Minghai Yang, Tongshan Zhou, Kai Li, and Dong Zhang. 2022. "Insights into the Antennal Characteristics and Olfactory Strategy of the Endangered Rhino Stomach Bot Fly Gyrostigma rhinocerontis (Diptera: Oestridae)" Insects 13, no. 10: 889. https://doi.org/10.3390/insects13100889
APA StyleXu, W., Li, X., Wang, Q., Zhang, C., Yang, M., Zhou, T., Li, K., & Zhang, D. (2022). Insights into the Antennal Characteristics and Olfactory Strategy of the Endangered Rhino Stomach Bot Fly Gyrostigma rhinocerontis (Diptera: Oestridae). Insects, 13(10), 889. https://doi.org/10.3390/insects13100889