1. Introduction
Most aspects of body physiology, metabolism, and behavior are controlled by the clock and lead to daily or seasonal strategies. The relationship between the timing of life cycle events and seasonal climatic changes (i.e., phenology) is a fundamental biological process in natural systems. Phenology is the main factor determining population dynamics, species interaction, animal movement, and the evolution of life history [
1,
2]. The timing of phenological events is gradually changing as a result of climate change [
3,
4,
5]. Along with other adaptive mechanisms, plasticity in phenology is essential for maintaining many aspects of biodiversity in a changing environment, such as species demography, species interaction, and species distribution [
6,
7,
8]. In response to seasonal natural changes, the species composition of populations and the number of species in them undergoes significant fluctuations [
9,
10,
11].
The rhythms of the vital activity of insects as poikilothermic animals are also adapted to seasonal environmental changes. Insects are particularly sensitive to an increase or decrease in temperatures above or below their optimum, to frost and drought, as well as to a decrease in the availability of resources, particularly food [
4,
12]. Therefore, seasonal rhythms of insect activity depend on a variety of environmental factors, most often on temperature, photoperiod, and humidity [
10,
13,
14]. In this regard, the observed climate changes lead to clear shifts in the phenology of species, changes in their life cycles of development and reproduction [
15,
16,
17,
18,
19,
20]. Thus, the seasonal aspects of insect biology are key processes that can link climate change to population conservation and possibly to community composition [
21].
Especially clear seasonal rhythms were found in a wide variety of insect groups living in temperate latitudes. In particular, the seasonal activity of species of Carabidae [
22,
23], Staphylinidae [
24], Mordellidae [
25], Scarabaeidae [
26], Cerambycidae [
27], Elateridae [
28], and many others. The phenological features of Lepidoptera and Hymenoptera of temperate climate have been well studied [
29,
30,
31,
32]. No less interesting are the seasonal dynamics of individual families and species of Diptera. In the forest zone of Russia in the second half of September, there was a gradual increase in the number of Diptera with a peak in mid-October. The autumn increase in the number of Diptera in different biotopes exceeded the summer peak several times [
33]. Based on the analysis of the activity of 194 Syrphidae species, ten phenological groups were identified, which differed in peaks of activity during the season [
34]. Many parasitic Diptera species depend on the seasonal activity of their hosts, which serve for the development of larvae [
35]. Anisopodidae activity occurs at the end of August and autumn [
36]. The seasonal activity of
Stomoxys calcitrans shows one large peak at the end of summer and a second smaller peak just before the end of the flight season [
37]. The phenological phases of
Ceratitis capitata development depended on the abundance of food items—various fruits [
38]. Generally known as fruit flies, family Drosophilidae consists of approximately 4000 species worldwide [
39,
40]. The majority of adult drosophilids feed on the bacteria and yeasts arising from the fermentation of various plant substrates (fruit, tree sap, rotting leaves, etc.). Their larvae also prefer the bacteria and yeasts arising from the fermentation of carbohydrates [
41] but some species feed on living mushrooms, living plant tissues as miners, etc., [
42]. The aim of the research was to study the species diversity and seasonal dynamics of drosophilids in various forest biotopes of the center of the European part of Russia. The objectives of the research were: (1) study of the species diversity of drosophilids in various biotopes using beer traps; (2) study of the seasonal dynamics of mass species of drosophilids.
3. Results
3.1. Faunistic Composition
Until recently, the fauna of the Drosophilidae of the Republic of Mordovia was totally unknown. The first paper with a short regional drosophilid faunistic list considered ecological questions of insect post-fire forest recovery [
49]. This preliminary faunistic list includes 15 species in 6 genera of Drosophildae. Here we give an addition with a new list of Drosophilidae of the Republic of Mordovia consisting of 30 species in 9 genera.
Among the flies collected in beer traps we found 4 genera and 9 species of subfamily Steganinae and 5 genera and 21 species of subfamily Drosophilinae:
Steganinae
Amiota (Amiota) albilabris (Roth in Zetterstedt, 1860)
Amiota (Amiota) alboguttata (Wahlberg, 1839)
Amiota (Amiota) rufescens (Oldenberg, 1914)
*Amiota (Amiota) subtusradiata Duda, 1934
Amiota (Phortica) semivirgo Maca, 1977
Gitona distigma Meigen, 1830
Leucophenga maculata (Dufour, 1839)
Leucophenga quinquemaculata Strobl, 1893
*Stegana (Steganina) coleoptrata (Scopoli, 1763)
Drosophilinae
- 10.
*Chymomyza amoena (Loew, 1862)
- 11.
*Chymomyza caudatula Oldenberg, 1914
- 12.
Chymomyza costata (Zetterstedt, 1838)
- 13.
*Chymomyza fuscimana (Zetterstedt, 1838)
- 14.
*Drosophila (Dorsilopha) busckii Coquillett, 1901
- 15.
*Drosophila (Drosophila) funebris (Fabricius, 1787)
- 16.
Drosophila (Drosophila) histrio Meigen, 1830
- 17.
*Drosophila (Drosophila) hydei Sturtevant, 1921
- 18.
*Drosophila (Drosophila) immigrans Sturtevant, 1921
- 19.
*Drosophila (Drosophila) kuntzei Duda, 1924
- 20.
Drosophila (Drosophila) phalerata Meigen, 1830
- 21.
Drosophila (Drosophila) testacea von Roser, 1840
- 22.
Drosophila (Drosophila) transversa Fallen, 1823
- 23.
Drosophila (Sophophora) bifasciata Pomini, 1940
- 24.
*Drosophila (Sophophora) melanogaster Meigen, 1830
- 25.
Drosophila (Sophophora) obscura Fallen, 1823
- 26.
*Drosophila (Sophophora) tristis Fallen, 1823
- 27.
*Hirtodrosophila confusa (Staeger, 1844)
- 28.
*Hirtodrosophila trivittata (Strobl, 1893)
- 29.
Scaptodrosophila rufifrons (Loew, 1873)
- 30.
*Scaptomyza (Hemiscaptomyza) unipunctum (Zetterstedt, 1847)
3.2. Seasonal Dynamics of Drosophilidae
As a result of the study, 4725 individuals from 9 genera and 30 species were detected in 2019 (
Table 1).
As we can conclude from our results, nine drosophild species (D. obscura, D. histrio, D. kuntzei, D. testacea, D. phalerata, S. rufifrons, D. bifasciata, A. semivirgo and L. quinquemaculata) were the most abundant in 2019, e.g., each of them with total number of flies caught in traps more than 100 exemplars. The amount of flies belonging to these 9 species is 4496 exemplars, which is 95.15% of total drosophilid number in our collection. We consider the other 21 species collected in amounts less than 100 flies as relatively rare or weakly attracted to this type of traps.
Interestingly, the most abundant species of Drosophilidae demonstrate different patterns of seasonal dynamics. Six species, e.g., D. obscura, D. histrio, D. kuntzei, D. testacea, D. phalerata, and D. bifasciata, show very strong increases in collected drosophilid numbers in October. However, among this group, D. obscura and D. bifasciata show additional moderate summer increases in July, and D. histrio in May, August, and September. On the contrary, two species, S. rufifrons and A. semivirgo, show low numbers in May–June increasing in July up to maximum values in August followed by decreases in September–October. One species, L. quinquemaculata, demonstrates similar maximal numbers in May and October, decreases in June, noticeable increases in July, and minimal equal numbers in August–September.
3.3. Species diversity of Drosophildae
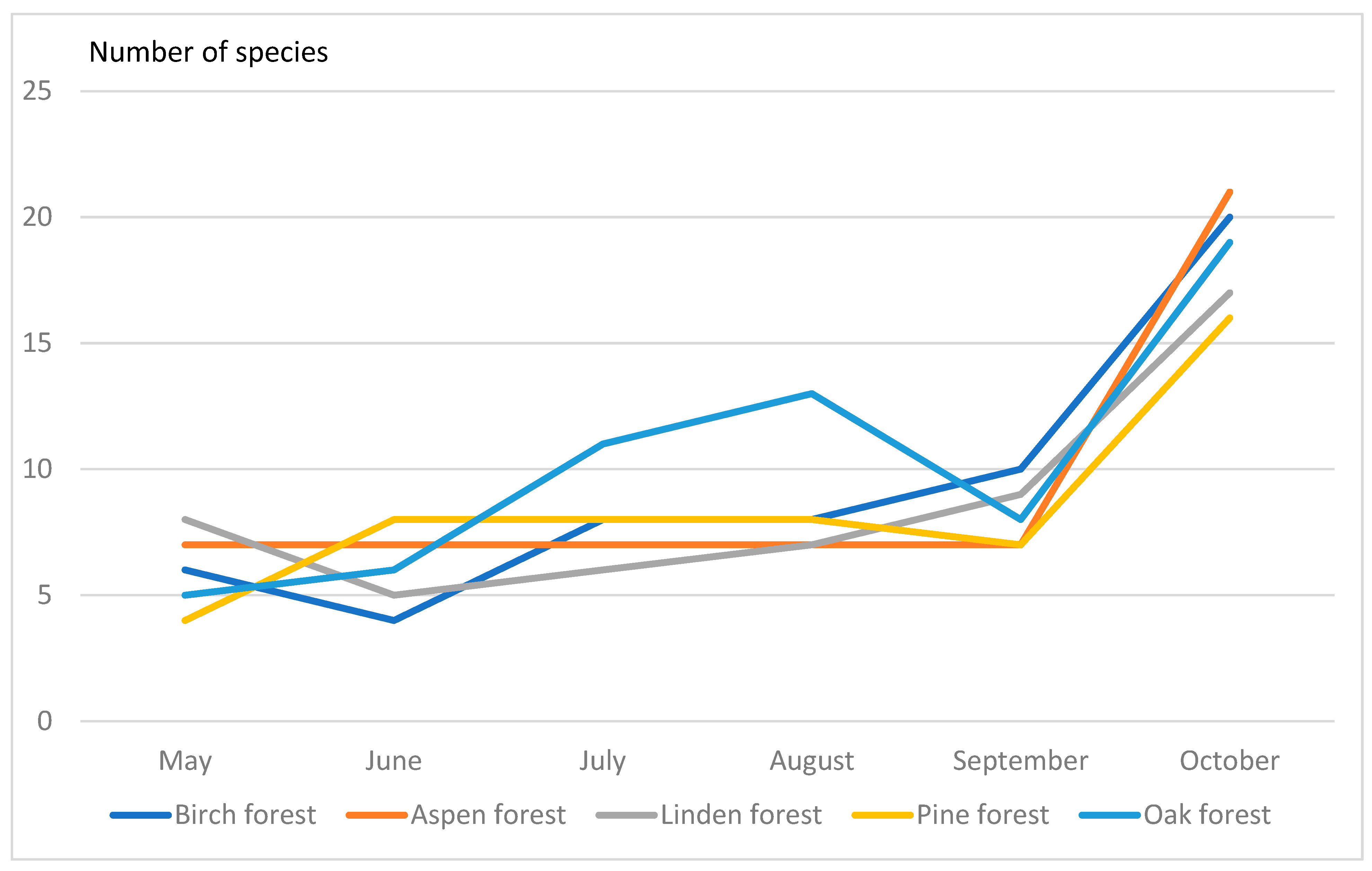
The drosophilid species diversity, e.g., number of collected species, varied between different types of forest since May to October (
Figure 1). We found that species diversity have maximal values in October in all types of forest examined.
In birch and linden forests, the number of drosophilid species was 20, in pine—19 species, in aspen—21 species. The greatest species diversity was observed in oak forest (23 species). At the same time, the calculated indices showed interesting results. Thus, according to the Shannon–Weaver index, the most diverse communities were in the linden forest (index 2.11), and the least diverse in the oak forest (index 1.87). In other communities, this index was intermediate and very similar (1.95–1.99). The Simpson index showed that the dominance of one or two drosophilid species is maximal in the oak forest (0.31). At the same time, in the linden forest, the dominance of species is the least pronounced (0.15), i.e., here the community is more aligned (
Table A10).
3.4. Seasonal Dynamics of Drosophilidae in Five Biotopes
We studied seasonal dynamics of Drosophilidae in five types of forest. We found that the drosophilid abundance was as follows: maximum value was in birch forest (1322) and the lowest in oak forest (640). Interestingly, the number of females exceeded the number of males in traps in all types of forest.
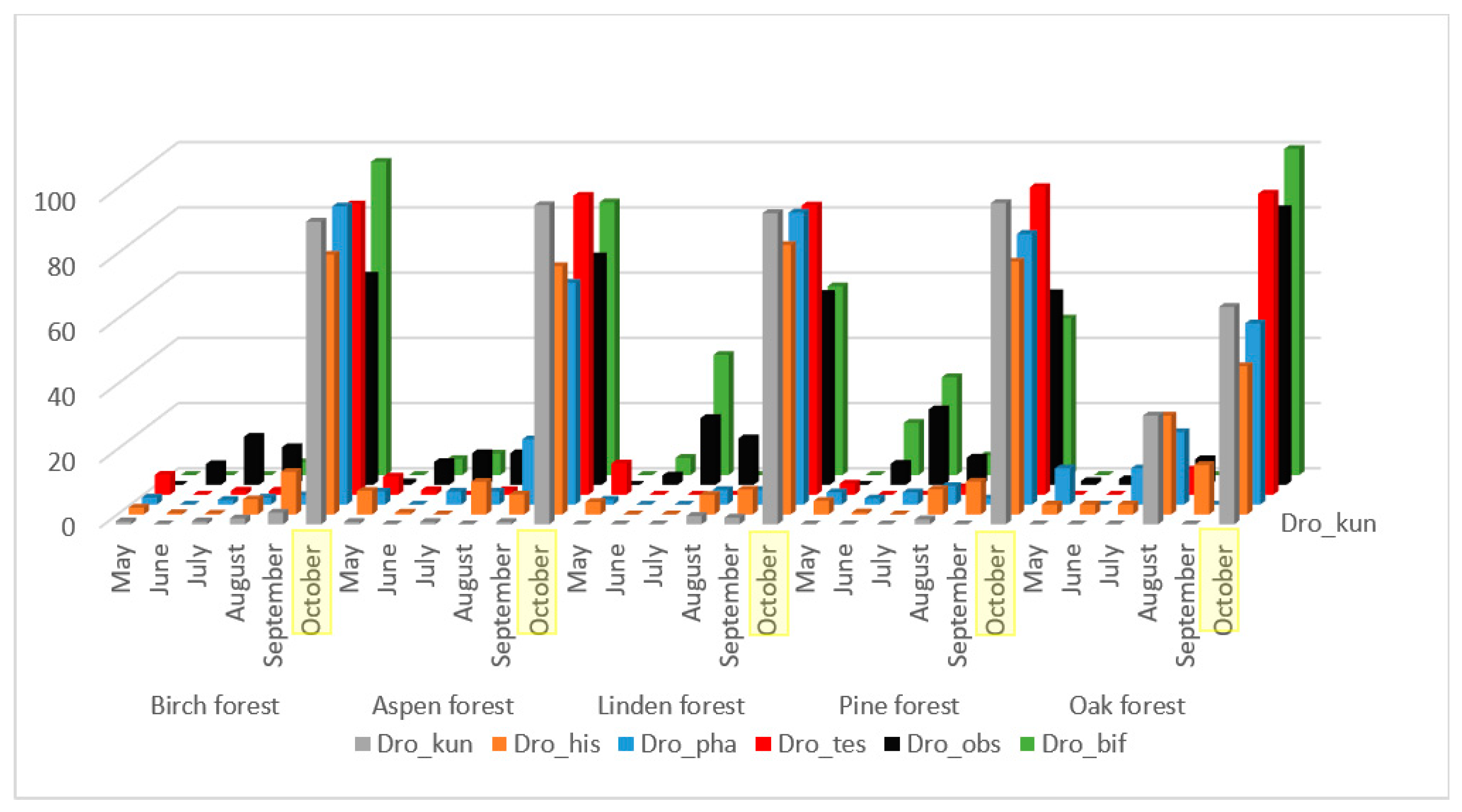
The majority of the mass species presented in
Figure 2, with the exception of
L. quinquemaculata, have a significant correlation of population fluctuations throughout the entire accounting season, from May to October (
Table 2). These species are characterized by low representation in June, an increase in numbers in July–September, and maximum representation in October. Some differences in seasonal dynamics by biotopes are caused by a small intermediate peak in the abundance of
D. kuntzei, D. histrio, and
D. phalerata species in August in oak forest collections, and in
D. obscura and
D. bifasciata species in linden and pine forests.
We found the highest significant correlation of seasonal dynamics between closely related species
D. obscura and
D. bifasciata (
Table 2). They are typical xylosaprobionts, their larvae live mainly in the tissues under the bark and in the fermenting tree sap [
42]. The second group with high significant correlation of seasonal dynamics consists of
D. histrio, D. kuntzei, D. phalerata, and
D. testacea. All these species are mycetobionts, their larvae live in various fungi.
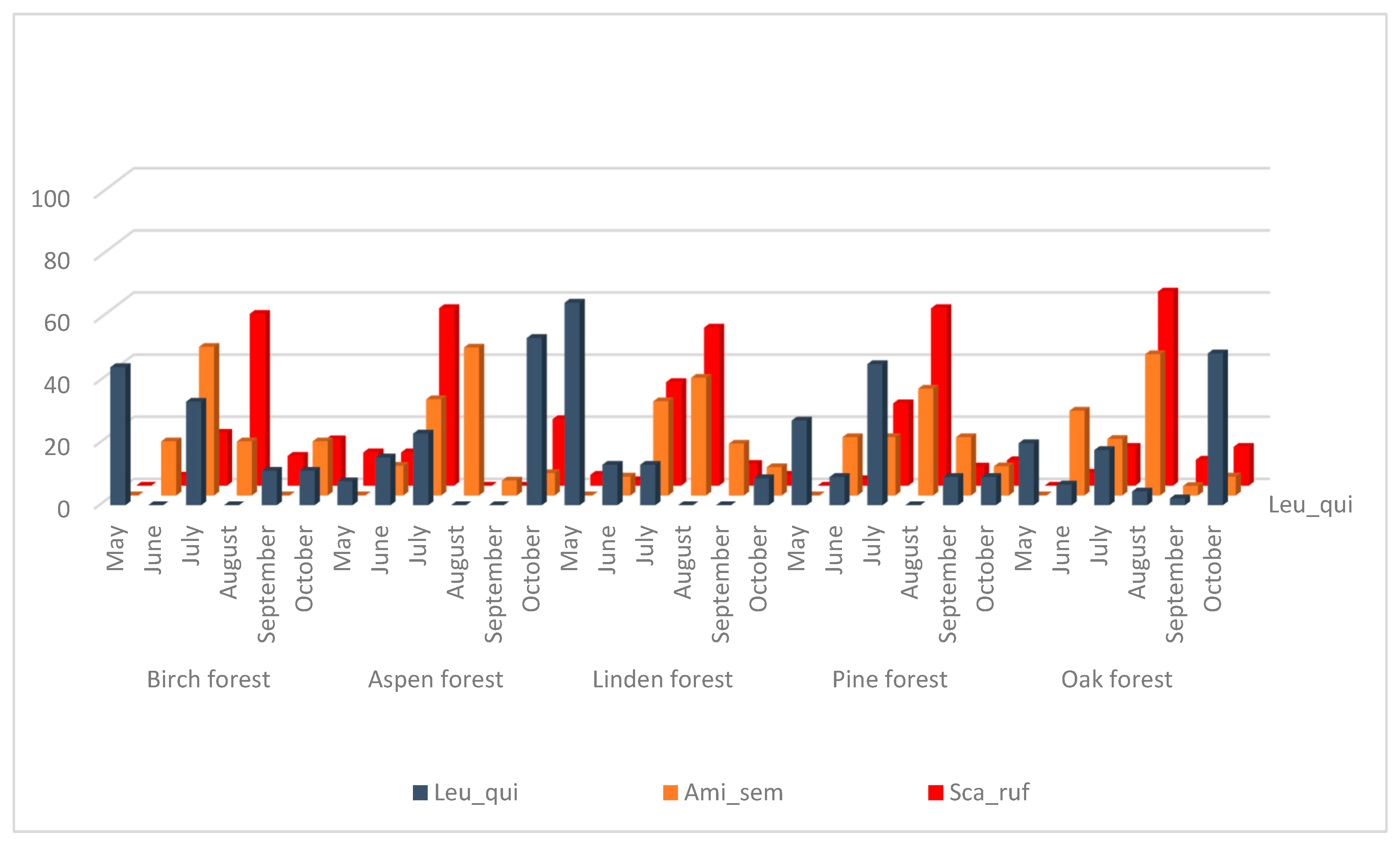
The species
Amiota semivirgo and
S. rufifrons also have significantly correlated seasonal dynamics (
Figure 3,
Table 3) but their main peak is observed in July–August in all biotopes, and in September–October, the number of collected flies decreases sharply. These species are also xylosaprobionts. Seasonal fluctuations in the number of
L. quinquemaculata species do not show a significant correlation with any of drosophilid species and show a maximum in May and July in birch and pine forests, in May only in the linden forest, and in October in aspen and oak forests. The larvae of
L. quinquemaculata could be found mainly in bracket fungi so they occupy a rather separate and specific ecological niche.
The group of mycetobionts developing mainly in various species of basidiomycetes includes mass species
D. histrio, D. kuntzei, D. phalerata, D. testacea (
Table A2,
Table A3,
Table A4 and
Table A5), and
L. quinquemaculata rearing in bracket fungi (
Table A9), which were found in an amount of more than 100 specimens (
Table 4).
The second large ecological group of drosophila includes xylosaprobionts (
D. obscura, D. bifasciata, S. rufifrons, A. semivirgo) (
Table A1,
Table A6,
Table A7 and
Table A8); their larvae live mainly in tissues under the bark and in fermenting tree sap (
Table 5).
4. Discussion
The influence of seasonal changes on the abundance of Drosophilidae has been studied mainly in tropical and temperate climatic zones. Their abundance in tropical regions is affected by precipitation, and in regions with a temperate climate, temperature fluctuations are most affected [
49,
50,
51,
52,
53]. Our studies have shown that Drosophilidae in central Russia have one peak in numbers, which begins at the end of September with a maximum in mid-October. At this time, daytime temperatures were recorded at no higher than 15 °C, and at night—no more than 10 °C. At the same time, throughout the season, the number of this family in traps was more or less constant without sharp peaks or lows. Similar dynamics were found in experiments in Uşak province, Turkey [
54]. The average temperature of October and November with the highest numbers of Drosophilidae was from 5 to 10 °C. At the same time, in September, when the temperature was more favorable for fruit flies, the amount of catch was less [
54].
Our work is the first study considering seasonal dynamics of Drosophilidae in European Russia. A total of 4725 individuals belonging to 9 genera and 30 species of drosophilid flies were identified in Mordovia State Nature Reserve. D. obscura and D. histrio were the most abundant species in beer traps. At the same time, seven more species (D. bifasciata, D. kuntzei, D. phalerata, D. testacea, L. quinquemaculata) were observed in traps with high numbers.
Among the 30 species of drosophila collected in Republic of Mordovia, 5 species of the genus
Drosophila (
D. busckii, D. funebris, D. hydei, D. immigrans, D. melanogaster) are synanthropic, i.e., closely related to humans and their activities. They live and breed in places where they can find fermenting and rotting fruits and vegetables, wine, beer and juices [
42,
55,
56,
57,
58]. These species occur in small numbers in wild biotopes, apparently, due to migration attempts or wind transport. Most of the other drosophilid species (24 in our collections) are typical forest dwellers, which rarely occur far from the forest or groups of trees. The larvae of these drosophilids develop in moist tissues under the bark of deciduous trees, in fermenting tree sap, and in various fungi, including ascomycetes and tinders [
59,
60,
61]. The larvae of the last species in our faunistic list,
Gitona distigma Mg., according to the literature, are phytophages living in inflorescences of family Asteraceae plants, e.g.,
Sonchus and
Crepis species [
42]. Therefore,
G. distigma may occur in different biotopes, not only in forests, sometimes even in people’s houses.
Here we compare the drosophilid fauna of the Republic of Mordovia with other regions of European Russia, we used data for the Moscow region—35 species [
62], Voronezh region—18 species [
62,
63], Samara region—13 species [
62,
64], and North Karelia—19 species [
65] (
Table 6).
As can be seen from
Table 6, the largest number of drosophilid species was observed in the Moscow region and Republic of Mordovia; this is a consequence of the special studies of this family conducted in these regions. Nevertheless, by now the degree of similarity is about 2/3 of the total number of species, we have found 21 common species for the fauna of the Republic of Mordovia and the Moscow region.
We studied seasonal dynamics of Drosophilidae in five types of forest (birch, aspen, linden, pine, and oak). Interestingly, the highest abundance of drosophilids was found in October in all types of the forests examined. We found that the drosophilid abundance demonstrated maximum value in birch forest and the lowest value in oak forest. In our collection we found representatives of two main ecological groups—mycetobionts and xylosaprobionts.
The total number of mass mycetobionts (2493) is 55.45% of the total number of drosophilid mass species (4496) and 52.76% of the total number of collected flies. At the same time, the larvae of
D. histrio, D. kuntzei, D. phalerata and
D. testacea develop mainly in the fruit bodies of basidiomycetes, and the larvae of
L. quinquemaculata develop in the bracket fungi. As can be seen from
Table 4, the number of imagos of
D. histrio, D. kuntzei, D. phalerata and
D.testacea collected in the oak forest is minimal, and several times less than in other biotopes. On the contrary,
L. quinquemaculata imagos were collected in maximum quantity in the oak forest. We suggest that this is due to noticeable differences in the composition of the mycoflora of oak forests and other types of forests. Apparently, the number of basidiomycetes growing in the oak forest was minimal or their species composition was less attractive for these drosophilid mycetobionts (
D. histrio, D. kuntzei, D. phalerata, and
D. testacea). On the contrary, bracket fungi, apparently, occur most often in the oak forest, which explains the largest number of
L.quinquemaculata collected here. The question of the relationship of various drosophilid species with fungi in Republic of Mordovia has not been studied yet but perhaps deserves a separate investigation.
The total number of mass xylosaprobionts (2003) is 44.55% of the total number of drosophilid mass species (4496) and 42.39% of the total number of flies collected. As can be seen from
Table 5, xylosaprobionts demonstrate the maximum abundance in oak and birch forests. Apparently, this is due to the greatest number of wounds on tree trunks in these biotopes, which attract drosophilids of these species (
D. obscura, D. bifasciata, S. rufifrons, A. semivirgo).
We found the highest significant correlation of seasonal dynamics between closely related xylosaprobiont species D. obscura and D. bifasciata. The second group with high significant correlation of seasonal dynamics consists of mycetobiont species D. histrio, D. kuntzei, D. phalerata, and D. testacea. The third group includes xylosaprobiont species A. semivirgo and S. rufifrons. Apparently, the similarity observed in the seasonal dynamics of some drosophilid species is influenced at a high degree by their food preferences and rearing sites.
We also analyzed species communities in five biotopes by calculating the Shannon–Weaver index and the Simpson index. It turned out that the greatest differences were found between oak and linden forests: the most diverse species community lives in the linden forest and the least diverse in the oak forest. On the contrary, the dominance of drosophilid species in the linden forest is the least pronounced, and in the oak forest it is the largest among all biotopes (
Table A10).
In addition, according to our data, the mass species of drosophilids of the Republic of Mordovia show a different picture of seasonal population peaks. They can be divided into different types: species with summer–autumn peaks of abundance (
D. obscura and
D. bifasciata), with spring–autumn peaks (
D. histrio, D. testacea), only with summer peaks (
A. semivirgo and
S. rufifrons), only with autumn peaks (
D. kuntzei, D. phalerata), and with three peaks of abundance (
L. quinquemaculata) (
Table 7). Therefore, we can conclude that the presence of two or three peaks in numbers of abundance suggests the presence of two or three generations in these drosophilids. However, the presence of one peak number in our collections does not negate the possibility of having two generations, for example, in
D. kuntzei and
D. phalerata. Perhaps, for these mycetobiont species, beer traps become less attractive in the summer during the mushroom abundance season. Interestingly, for six mass species of drosophilids, the autumn peak of abundance is the maximum.
5. Conclusions
In our study, Drosophilidae species and their seasonal changes in Mordovia State Reserve were explored. It is the first investigation of drosophilid seasonal population changes considering their biotope association, abundance and species diversity in European Russia. We collected the flies by crown fermental traps in five types of forests (birch, aspen, linden, pine, and oak) from May to October in 2019. A total of 4725 individuals belonging to 9 genera and 30 species of drosophilid flies were identified, among them 15 species in 3 genera are new to Republic of Mordovia. Drosophila obscura Fll. and D. histrio Mg. were the most abundant species in traps, the other mass species are D. kuntzei, D. testacea, D. phalerata, S. rufifrons, D. bifasciata, A. semivirgo, and L. quinquemaculata. Interestingly, the highest abundance of drosophilids and their species diversity was found in October in all types of the forests examined. We found the highest significant correlation of seasonal dynamics between closely related species D. obscura and D. bifasciata, the second group with high significant correlation of seasonal dynamics consists of D. histrio, D. kuntzei, D. phalerata, and D. testacea, and finally the third group consists of A. semivirgo and S. rufifrons. Apparently, the similarity observed in the seasonal dynamics of these drosophilid species is influenced at high degree by their food preferences and rearing sites.





{kind=link}
{kind=link}
{kind=link}


