3.2. Taxonomy
Order Hemiptera Linnaeus, 1758
Suborder Heteroptera Latreille, 1810
Infraorder Cimicomorpha Leston, Pendergrast and Southwood, 1954
Family Reduviidae Latreille, 1807
Subfamily Peiratinae Amyot & Serville, 1843
GenusSigicorisMiller, 1958 stat. nov. (
Figure 2,
Figure 3,
Figure 4,
Figure 5,
Figure 6,
Figure 7,
Figure 8,
Figure 9,
Figure 10,
Figure 11,
Figure 12,
Figure 13,
Figure 14,
Figure 15,
Figure 16 and
Figure 17)
Sigicoris Miller, 1958: 74. As subgenus of Ectomocoris; Maldonado-Capriles, 1990: 350. As junior synonym of Ectomocoris.
Type species: Ectomocoris (Sigicoris) gracilis Miller, 1958, by original designation.
Diagnosis: Members of the genus can be recognized among Peiratinae by the following combination of characteristics: head with anteclypeus raised, postocular part ellipsoidal; width of eye shorter than width of interocular space; 1 + 1 tubercles of neck prominent, surface granulose; anterior lobe of pronotum with an elliptical depression on posterior portion, stripes nearly invisible; disc of scutellum broad and granulose, scutellar process slender, apex slightly directed obliquely backward in lateral view; anterior half of metapleural sulcus slightly curved, posterior half nearly straight; ventral surfaces of fore and mid femora with rows of tiny denticles, and each denticle bearing an erect, long seta apically; fore tibia slightly recurved in apical half, with fossula spongiosa occupying at least half of tibial length ventrally; mid tibia with fossula spongiosa occupying at least 1/3 of ventral surface; hemelytron with Cu on corium reduced, Cu and Pcu on membrane short and nearly straight, inner cell relatively short and somewhat quadrilateral.
Description: Coloration. Mostly brown to blackish brown, sometimes with yellowish white spots on hemelytron and femora and yellowish spots on connexivum.
Structure. Macropterous form. Most of body surface covered with scattered setae of varying lengths; costal margin of corium densely covered with short pubescence; ventral surface of abdomen covered with procumbent pubescence (
Figure 2,
Figure 3,
Figure 6,
Figure 7,
Figure 10,
Figure 11,
Figure 13,
Figure 14 and
Figure 15). Head moderately elongated, drop-shaped in dorsal view, anteocular part distinctly longer than postocular; anteclypeus raised; interocular space with a median short sulcus connecting to frontoclypeal sulcus; postocular part ellipsoidal (
Figure 4A,
Figure 8A,
Figure 12A and
Figure 16A). Eye oval in dorsal view, width of eye shorter than width of interocular space (
Figure 4A,
Figure 8A,
Figure 13A and
Figure 16A); eye reniform in lateral view, reaching dorsal margin but not reaching ventral margin of head (
Figure 4B,
Figure 8B,
Figure 12B and
Figure 16B). Ocelli located on small tubercles, width of interocellar space subequal to slightly longer than width of ocellus (
Figure 4A,
Figure 8A,
Figure 12A and
Figure 16A). Antenna long and gracile, with first antennal segment thickest and shortest, feebly curved, and second segment straight and the longest. First and second visible rostral segments thick; third segment tapered; second segment longest (
Figure 4C,
Figure 8C,
Figure 12C and
Figure 16C). Neck with 1 + 1 tubercles prominent, surface of tubercle granulose (
Figure 4A,C,
Figure 8A,C,
Figure 12A,C and
Figure 16A,C).
Pronotum with collar process developed, subconical; length of anterior lobe of pronotum less than twice the length of posterior lobe, anterior lobe with anterior margin almost straight or slightly concave, lateral margins slightly rounded, sculpture nearly invisible; depression on basal of anterior lobe elliptical with a median longitudinal sulcus; pronotal transverse sulcus above with some longitudinal short wrinkles; lateral pronotal angle rounded or narrowly rounded, posterior margin of pronotum convex (
Figure 4A,
Figure 8A,
Figure 12A and
Figure 16A). Scutellum triangular, “Y” shaped ridges and scutellar process slender (
Figure 4A,
Figure 8A,
Figure 12A and
Figure 16A), apex of process slightly directed obliquely backward in lateral view (
Figure 4B,
Figure 8B,
Figure 12B and
Figure 16B), disc of scutellum broad and finely granulose with a median, longitudinal, shallow sulcus (
Figure 4A,
Figure 8A,
Figure 12A and
Figure 16A). Stridulitrum long with total-striate type of sculpture (
Figure 4C,
Figure 8C,
Figure 12C and
Figure 16C). Pleura and sterna finely granulose (
Figure 4B,C,
Figure 8B,C,
Figure 12B,C and
Figure 16B,C); anterior half of metapleural sulcus slightly curved, posterior half nearly straight (
Figure 4B,
Figure 8B,
Figure 12B and
Figure 16B); mesosternum with a median longitudinal ridge (
Figure 4C,
Figure 8C,
Figure 12C and
Figure 16C). Fore coxa elongate; fore femur not obviously thickened but thicker than mid and hind femora; fore and mid femora with rows of tiny denticles ventrally, and each denticle bearing an erect long seta apically (
Figure 4D,E,
Figure 8D,E,
Figure 12E and
Figure 16D,E); fore tibia slightly recurved in apical half, ventral surface with fossula spongiosa occupying at least half of tibial length (
Figure 4D,
Figure 8D,
Figure 12E and
Figure 16D); mid tibia with fossula spongiosa occupying at least 1/3 of tibial length (
Figure 4E,
Figure 8E,
Figure 12F and
Figure 16E); hind coxae separated from each other less than width of one coxa (
Figure 4C,
Figure 8C,
Figure 12C and
Figure 16C). Hemelytron extending beyond tip of abdomen in macropterous male (
Figure 2B,
Figure 7B and
Figure 14B) and nearly reaching tip of abdomen in macropterous female (
Figure 3A,
Figure 6A,
Figure 11A and
Figure 15A); Cu on corium reduced, getting fainter from base to apex, Pcu and Cu on membrane short and nearly straight, inner cell relatively short and somewhat quadrilateral (
Figure 4F,
Figure 8F,
Figure 12D and
Figure 16F).
Abdomen mainly oval, posterior margin nearly straight with middlemost feebly concave onwards in male (
Figure 4G,
Figure 8G,
Figure 10B,
Figure 12H and
Figure 14B), tip of abdomen pointed in female (
Figure 3B,
Figure 6B,
Figure 11B,
Figure 12G,
Figure 15B and
Figure 16G); connexivum slightly dilated laterally (
Figure 2A,
Figure 3A,
Figure 6A,
Figure 7A,
Figure 10A,
Figure 11A,
Figure 13A,
Figure 14A and
Figure 15A); venter of abdomen smooth in female (
Figure 3B,
Figure 6B,
Figure 11B,
Figure 12G,
Figure 15B and
Figure 16G) but slightly carinate longitudinally in middle in male (
Figure 2B,
Figure 4G,
Figure 7B,
Figure 8G,
Figure 13C and
Figure 14B).
Male genitalia asymmetric (
Figure 5 and
Figure 9). Pygophore oval in ventral view (
Figure 5A and
Figure 9A); median pygophore process long and slender (
Figure 5A–C and
Figure 9A–C). Parameres subtriangular (
Figure 5D,E) or sickle-shaped (
Figure 9D,E), left paramere slightly longer and slenderer than right paramere. Phallus with basal plate bridge feebly curved and slightly longer than basal plate (
Figure 5F and
Figure 9F). Dorsal phallothecal sclerite broad and flat (
Figure 5F and
Figure 9F); lateral phallothecal sclerite moderately sclerotized, lower half of inner margin with two processes (
Figure 5I and
Figure 9I).
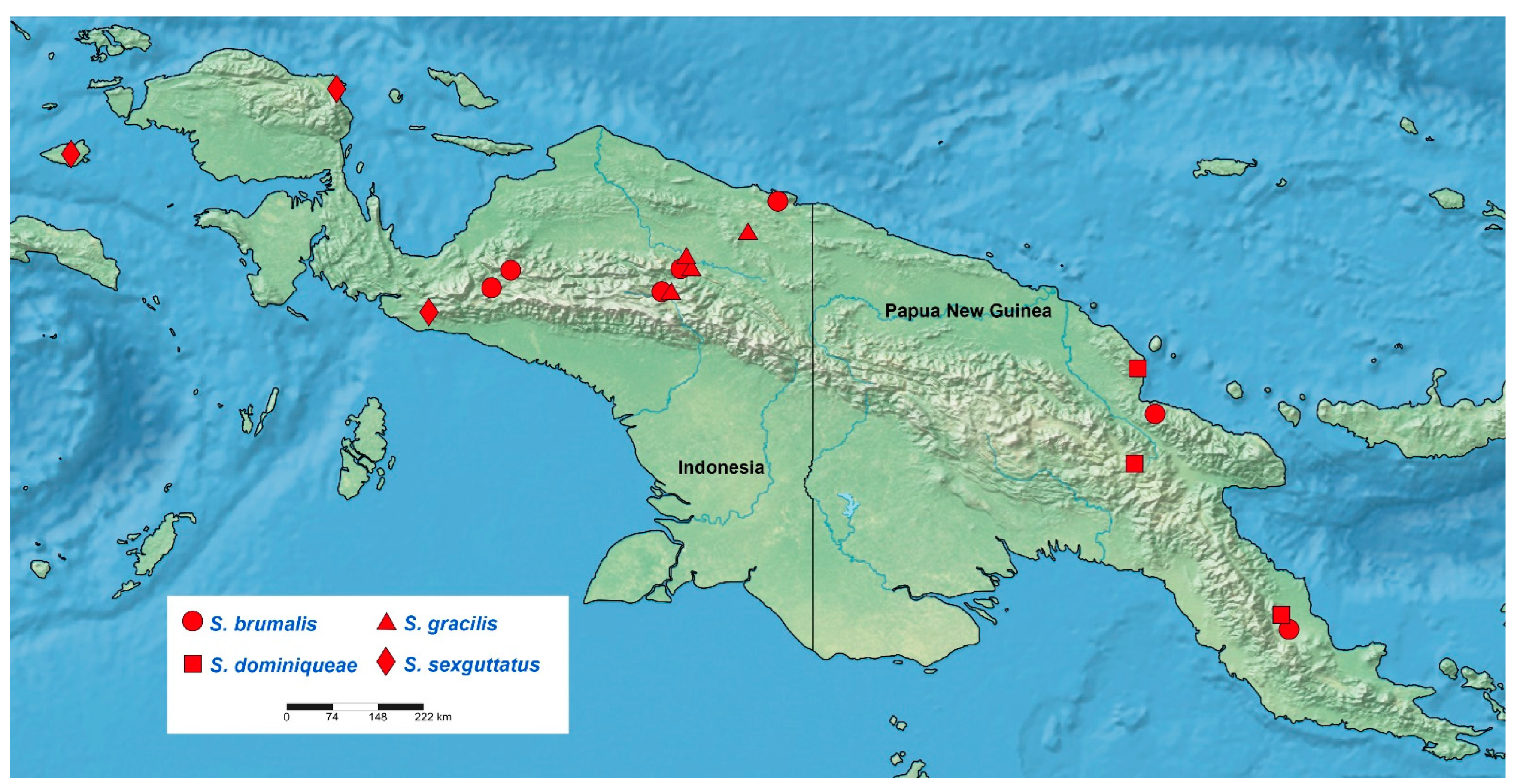
Distribution: Australian Region (New Guinea) (
Figure 17).
Systematic relationships of Sigicoris stat. nov.
We make the first attempt of the molecular phylogenetic analysis of the whole subfamily, with the taxon sampling containing over 70% of the peiratine genera (25/35) based on the COI, 16S and 18S genes. The tree topology helps us further confirm the monophyly and genus status of Sigicoris stat. nov., which indicates the application of molecular data in taxonomic revision. Sigicoris gracilis comb. nov. is not included in the phylogenetic analysis due to the specimen limitation. Sigicoris gracilis comb. nov. is the type species of Sigicoris and can be included in this genus as it shares the morphological characters as described below and the endemic distribution (New Guinea) of the other Sigicoris stat. nov. species.
Sigicoris was first proposed as a subgenus of Ectomocoris with two species, E. (Sigicoris) brumalis and E. (Sigicoris) gracilis, while some members of Sigicoris stat. nov. were originally assigned in Brachysandalus or Peirates. Sigicoris stat. nov., indeed, shares some characters with these three genera, such as the denticles on the ventral surfaces of the fore and mid femora with Brachysandalus, the similar length ratio of the anterior lobe of the pronotum to the posterior lobe with Peirates and the prominent fossula spongiosa with Ectomocoris. However, the reduced Cu on the corium and the short and nearly straight Pcu and Cu resulting in the relatively short and somewhat quadrilateral inner cell on the membrane make Sigicoris stat. nov. quite unique even in the whole Peiratinae. Except the characters listed above, Sigicoris stat. nov. can also be distinguished from Brachysandalus by the shape of the fore tibia (slightly recurved in the apical half in Sigicoris stat. nov. vs. clavate and gradually thickened to the apex in Brachysandalus), and from Peirates by the less distinct and even nearly invisible sculpture on the pronotum. As for the genus Ectomocoris to which Sigicoris used to belong, the latter could be separated from the former by the smaller eye, the less thickened fore femur, the less elongated anterior lobe of the pronotum and the shape of the abdomen in males (the posterior margin of the abdomen nearly straight with middlemost feebly concave onwards in Sigicoris stat. nov. vs. the posterior margin of the abdomen arcuate in Ectomocoris). In addition, the three sampled Sigicoris species are grouped together in our phylogenetic tree and not nested within the Ectomocoris clade that includes the type species E. quadriguttatus. The results of the morphological comparison and molecular phylogenetic analysis both demonstrate that Sigicoris should be separated from Ectomocoris as a valid genus.
So far, the only two species in Ectomocoris distributed in New Guinea are E. mimomyrmix and E. olthofi. We examined the apterous female holotype of E. mimomyrmix deposited in NHM (label information: “Holotype” disc // red-margined “Type” disc // “PAPUA: Kokoda. 1200 ft. ix. 1933. L. E. Cheesman. B.M. 1934-321.” // “Ectomocoris mimomyrmix sp. n. N.C.E. Miller det. 1950” // “NHMUK 013586064”). Unfortunately, this holotype is badly damaged: the left antenna, the second to fourth segments of the right antenna and most of the legs (except one mid femur and tibia stuck on the card) are missing and the thorax is broken and glued together. Thus, more specimens, especially male and macropterous ones, are needed to further confirm the status of this species. We also examined one male paratype of E. olthofi deposited in NHM (label information: yellow-margined “Paratype” disc // “Neth. American New Guinea Exedit. Bernhard camp, 50 m, 2 ix. 1938, J. Olthof” // “Ectomocoris olthofi sp. n. (paratype) N.C.E.Miller det. 1956” // “NHMUK 013585956”) and confirmed its status due to the typical diagnostic characters of Ectomocoris such as the anterior lobe of the pronotum more than twice longer than the posterior lobe, the strongly thickened fore femur and the fossula spongiosa nearly occupying the whole ventral surfaces of the fore and mid tibiae. Therefore, currently there are four peiratine genera distributed in New Guinea: Ceratopirates, Ectomocoris, Sigicoris stat. nov. and Sirthenea.
Key to the species of Sigicoris stat. nov.
1. Pcu and Cu on membrane slightly curved (
Figure 16F); legs and connexivum bicolored (
Figure 13,
Figure 14 and
Figure 15)...................................................................................
S. sexguttatus comb. nov.2. First visible segment of rostrum shorter than third segment (
Figure 8C); fore tibia with fossula spongiosa occupying half of tibial length (
Figure 8D); mid tibia with fossula spongiosa occupying 1/3 of tibial length (
Figure 8E).................................
S. dominiqueae sp. nov.– First visible segment of rostrum longer than third segment (
Figure 4C and
Figure 12C); fore tibia with fossula spongiosa occupying 2/3 of tibial length (
Figure 4D and
Figure 12E); mid tibia with fossula spongiosa occupying half of tibial length (
Figure 4E and
Figure 12F)....................3
3. Coloration blackish brown (
Figure 2 and
Figure 3); width of interocellar space slightly longer than width of ocellus (
Figure 4A)
........................................................S. brumalis comb. nov.– Coloration brown (
Figure 10 and
Figure 11); width of interocellar space subequal to width of ocellus (
Figure 12A)
.................................................................................S. gracilis comb. nov.
Ectomocoris (Sigicoris) brumalis Miller, 1958: 75–76. Central West New Guinea, Lake Paniai.
Ectomocoris brumalis: Maldonado-Capriles, 1990: 351.
Type material examined: Indonesia: Paratype, 1 male, yellow-margined “Paratype” disc // “Museum Leiden Nieuw Guinen Exp K. N. A. G. 1939 Araboebivak 17. X. 1939” // “Sigicoris brumalis subgen. n. sp. n. (paratype) N. C. E. Miller det. 1956” // “NHMUK 013585911” (NHM); Paratype, 1 female, yellow-margined “Paratype” disc // “Neth. Ind.-Amer. New Guinea Exp. Lower Mist Camp 1500 m 30 S. 1939 L. J. Toxopeus leg.” // “Sigicoris brumalis subgen. n. sp. n. (paratype) N. C. E. Miller det. 1956” // “NHMUK 013585912” (NHM).
Other material examined: Indonesia: 1 female, “DUTCH NEW GUINEA: Cyclops Mts. Sabron. Camp 1: 1200 ft. 15. v. 1936. L. E. Cheesman. B.M. 1936-271” // “NHMUK 013585913” (NHM); Papua New Guinea: 1 male, 1 female, “PAPUA: Kokoda. 1200ft. ix. 1933. L. E. Cheesman. B.M. 1934-321” // “NHMUK 013585915” & “NHMUK 013585916” (NHM); 2 males, 2 females, “Stn. No. 46.” // “NEW GUINEA: Madang Dist., Finisterre Mts. Damanti 3550 ft. 2-11. x. 1964” // “M.E. Bacchus. B.M. 1965-120” // “NHMUK 013585917” to “NHMUK 013585920” (NHM).
Distribution: Indonesia (Papua), Papua New Guinea (Oro, Madang).
Figure 2.
Sigicoris brumalis comb. nov., paratype male, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Figure 2.
Sigicoris brumalis comb. nov., paratype male, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Diagnosis: Body blackish brown in large part; width of interocellar space subequal to width of ocellus; first visible segment of rostrum longer than third segment; pronotum with anterior margin slightly concave, collar process prominent, lateral pronotal angle narrowly rounded; fore tibia with fossula spongiosa occupying 2/3 of tibial length, mid tibia with fossula spongiosa occupying half of tibial length; median pygophore process long, slender and straight, apex sharp, slightly oblique to left side in caudal view, base of inner margin serrated in lateral view; parameres subtriangular with inner margin concave; lower half of inner margin of lateral phallothecal sclerite with two processes, one upward and one downward.
Figure 3.
Sigicoris brumalis comb. nov., paratype female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Figure 3.
Sigicoris brumalis comb. nov., paratype female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Redescription: Macropterous form (
Figure 2 and
Figure 3).
Coloration. Blackish brown. Antennae, third visible segment of rostrum and tarsi brown (
Figure 2 and
Figure 3); hemelytron dark brown, with most of clavus and area between Pcu and Cu on corium paler, base of costal area of membrane with a diffuse pale brown stripe (
Figure 4F).
Figure 4.
Sigicoris brumalis comb. nov., male. (A) Anterior part of body with antennae and legs removed, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Left fore leg, ventral view. (E) Right mid leg, ventral view. (F) Right hemelytron, dorsal view. (G) Abdomen, ventral view; (H) ditto, caudal view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Figure 4.
Sigicoris brumalis comb. nov., male. (A) Anterior part of body with antennae and legs removed, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Left fore leg, ventral view. (E) Right mid leg, ventral view. (F) Right hemelytron, dorsal view. (G) Abdomen, ventral view; (H) ditto, caudal view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Structure. Antennae, head, disc of scutellum, pleura, sterna, coxae, ventral surfaces and apex of femora, tibiae, lateral margin of corium, eighth abdominal sternite and genitalic part densely covered with golden, procumbent, short pubescence; first and second antennal segments, lateral margins of head and pronotum, femora and tibiae covered with brown, suberect to erect setae of varying lengths; apex of dorsal surfaces of tibiae and ventral surfaces of tarsi densely covered with yellow to yellowish brown, suberect setae; venter of abdomen sparsely covered with golden, long pubescence (
Figure 2 and
Figure 3).
Figure 5.
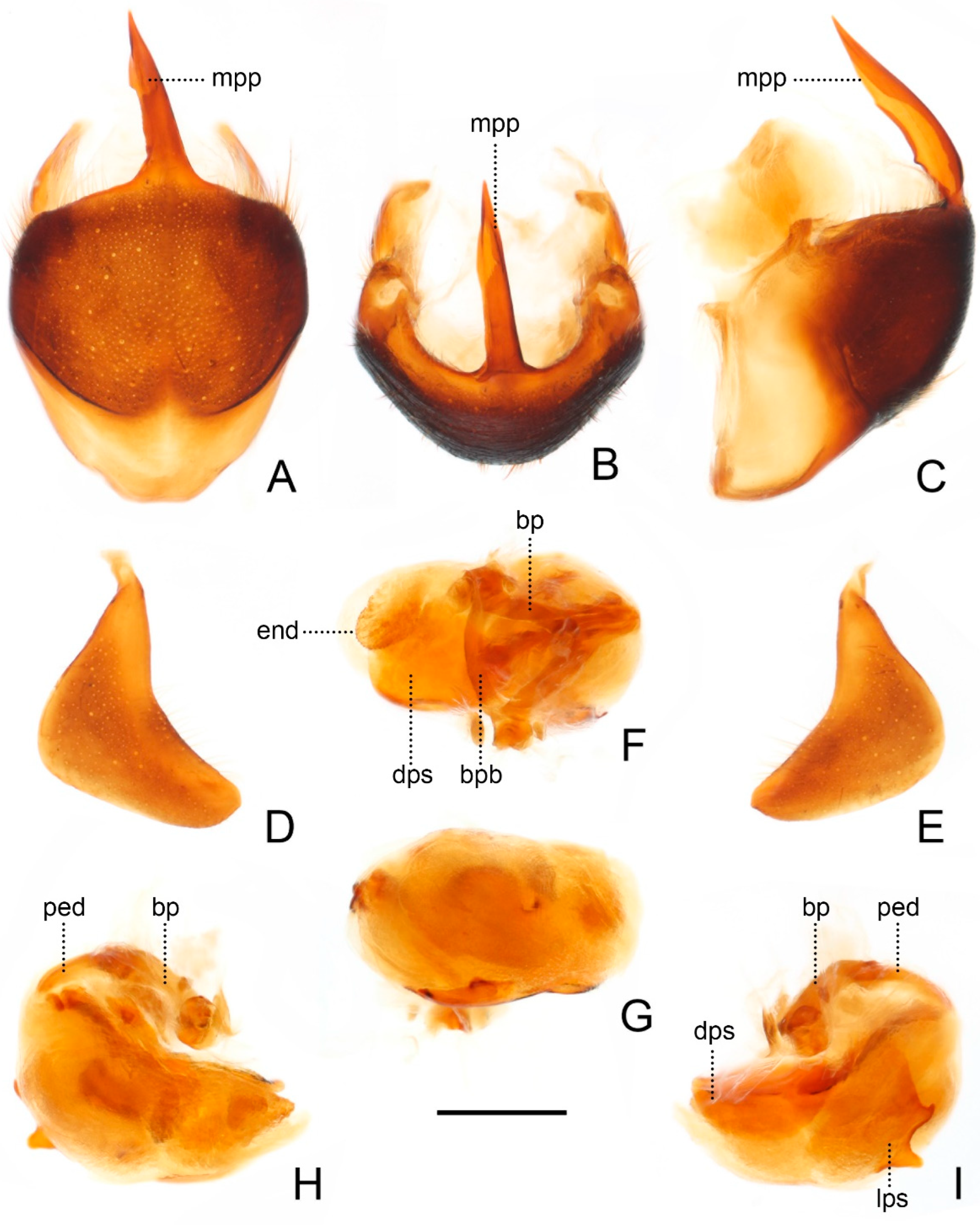
Male genitalia of Sigicoris brumalis comb. nov. (A) Pygophore, ventral view; (B) ditto, caudal view; (C) ditto, lateral view. (D) Left paramere, outer ventrolateral view. (E) Right paramere, outer ventrolateral view. (F) Phallus, dorsal view; (G) ditto, ventral view; (H) ditto, left lateral view; (I) ditto, right lateral view. Scale bar = 0.50 mm. Abbreviations: bp, basal plate; bpb, basal plate bridge; dps, dorsal phallothecal sclerite; end, endosoma; lps, lateral phallothecal sclerite; mpp, median pygophore process; ped, pedicel.
Figure 5.
Male genitalia of Sigicoris brumalis comb. nov. (A) Pygophore, ventral view; (B) ditto, caudal view; (C) ditto, lateral view. (D) Left paramere, outer ventrolateral view. (E) Right paramere, outer ventrolateral view. (F) Phallus, dorsal view; (G) ditto, ventral view; (H) ditto, left lateral view; (I) ditto, right lateral view. Scale bar = 0.50 mm. Abbreviations: bp, basal plate; bpb, basal plate bridge; dps, dorsal phallothecal sclerite; end, endosoma; lps, lateral phallothecal sclerite; mpp, median pygophore process; ped, pedicel.
Head length about 1.29 times as long as width in male and about 1.38 times as long as width in female; median short sulcus on interocular space deep (
Figure 4A); width of interocellar space slightly longer than width of ocellus (
Figure 4A); first visible segment of rostrum about 1.15 times longer than third segment (
Figure 4B). Pronotum with anterior margin slightly concave, collar process prominent (
Figure 4A); anterior lobe of pronotum about 1.59 times longer than posterior lobe in male and about 1.40 times longer than posterior lobe in female; lateral pronotal angle narrowly rounded (
Figure 4A). Fore tibia with fossula spongiosa occupying 2/3 of tibial length (
Figure 4D), mid tibia with fossula spongiosa occupying half of tibial length (
Figure 4E).
Male genitalia asymmetric (
Figure 5). Pygophore oval in ventral view (
Figure 5A); median pygophore process long, slender and straight, apex sharp (
Figure 5A–C), slightly oblique to left side in caudal view (
Figure 5B), base of inner margin serrated in lateral view (
Figure 5C). Parameres subtriangular with inner margin concave, left paramere (
Figure 5D) slightly longer and slenderer than right paramere (
Figure 5E). Phallus with phallobase strongly sclerotized, basal plate bridge feebly curved and slightly longer than basal plate (
Figure 5F); pedicel nearly straight and shorter than basal plate (
Figure 5H,I). Dorsal phallothecal sclerite distinctly sclerotized, broad and flat (
Figure 5F, H and I); lateral phallothecal sclerite moderately sclerotized, lower half of inner margin with two processes, one upward and one downward (
Figure 5I). Apical portion of endosoma with a sacciform process, surface of which is covered with rows of tiny spine-like tubercles (
Figure 5F,H).
Measurements [in mm, male (n = 4), female (n = 5)]: Body length 12.68–15.60 (male), 17.22–17.27 (female); maximum width of abdomen 3.54–4.20 (male), 4.56–5.18 (female); head length 1.71–1.74 (male), 1.91–2.08 (female); length of anteocular part 0.79–0.75 (male), 0.95–1.12 (female); length of postocular part 0.42–0.46 (male), 0.45–0.49 (female); head width 1.31–1.38 (male), 1.44–1.45 (female); eye width in dorsal view 0.38–0.47 (male), 0.45–0.41 (female); width of interocular space 0.49–0.58 (male), 0.51–0.62 (female); width of interocellar space 0.29–0.28 (male), 0.32–0.26 (female); lengths of rostral segments I:II:III = 0.73–0.91:1.23–1.51:0.68–? (male), 0.89–0.98:1.42–1.55:0.75–0.82 (female); lengths of antennal segments I:II:III:IV = 1.25–1.42:2.67–2.81:2.33–2.25:1.93–2.42 (male), 1.31–1.59:2.71–2.88:1.95–?: 1.96–? (female); length of anterior pronotal lobe 1.78–2.03 (male), 1.78–2.25 (female); length of posterior pronotal lobe 1.12–1.27 (male), 1.39–1.49 (female); width of anterior pronotal lobe 1.92–2.31 (male), 2.30–2.50 (female); width of posterior pronotal lobe 3.20–3.81 (male), 3.79–4.32 (female); scutellum length 1.33–1.52 (male), 1.78–1.71 (female); maximum width of scutellum 1.56–1.88 (male), 2.02–2.10 (female); hemelytron length 8.98–11.08 (male), 10.36–11.99 (female).
Remarks: The holotype of this species is deposited in Rijksmuseum van Natuurlijke Historie, Leiden; we only examined the paratypes deposited in the Natural History Museum, London.
Type material: Indonesia: Holotype female, red-margined “Holotype” disc // “In logs” // “PAPUA: Kokoda. 1200 ft. vS. 1933. L. E. Cheesman. B.M. 1933-577.” (NHM); Paratype 1 female, yellow-margined “Paratype” disc // “Coll. S. R. Sc. N. B. Canopy Mission Papua Neu Guinea (Madang prov): Baiteta 04. VS. 1996 Light trap AR7 Leg. Olivier Missa” orange rectangle label (IRSNB); Paratype 1 male, yellow-margined “Paratype” disc // “OKASA, E. H. D. UNDER BARK FOLLEN HOOP PINE. PINE FOREST. d 3. VIIS. 1967. F. R. WYLIE & S. AUNO” // “123” // “C.S.E.COLL. A. 5212” // “NHMUK 013588800” (NHM).
Distribution: Papua New Guinea (Madang, Oro, Eastern Highlands).
Etymology: The specific epithet is dedicated to the French entomologist, Dominique Pluot-Sigwalt (Muséum National d’Histoire Naturelle), in honor of her contributions to entomology and great support to our research.
Diagnosis: Body reddish brown to dark brown, pronotum with anterior lobe blackish brown and posterior lobe dark brown; width of interocellar space longer than width of ocellus; third visible segment of rostrum longer than first segment; pronotum with anterior margin nearly straight, collar process prominent, lateral pronotal angle rounded; fore tibia with fossula spongiosa occupying half of tibial length, mid tibia with fossula spongiosa occupying 1/3 of tibial length; median pygophore process long and extremely slender, apex sharp, nearly vertical in caudal view and curved in lateral view; parameres sickle-shaped; phallobase and dorsal phallothecal sclerite less sclerotized; lower half of inner margin of lateral phallothecal sclerite with two upward processes.
Figure 6.
Sigicoris dominiqueae sp. nov., holotype female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Figure 6.
Sigicoris dominiqueae sp. nov., holotype female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Description: Macropterous form (
Figure 6 and
Figure 7).
Coloration. Reddish brown to dark brown. Head, anterior lobe of pronotum, scutellum, pleura and sterna blackish brown; third and fourth segments of antenna brown; tarsi yellowish brown (
Figure 6 and
Figure 7); hemelytron dark brown, with apical half of clavus, area between Pcu and Cu on corium and apical portion of membrane paler, base of costal area of membrane with a diffuse pale brown stripe (
Figure 8F).
Structure. Antennae, marginal area of head, disc of scutellum, pleura, sterna, coxae, ventral surfaces and apex of femora, tibiae, lateral margin of corium and venter of abdomen densely covered with golden, procumbent, short pubescence; first and second antennal segments, lateral margins of head and pronotum, femora and tibiae covered with brown, suberect to erect setae of varying lengths; apex of dorsal surfaces of tibiae and ventral surfaces of tarsi densely covered with golden, suberect setae; venter of abdomen also sparsely covered with golden, long pubescence (
Figure 6 and
Figure 7).
Figure 7.
Sigicoris dominiqueae sp. nov., paratype male, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Figure 7.
Sigicoris dominiqueae sp. nov., paratype male, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Head length is 1.41 times as long as width in male and about 1.46 times as long as width in female; median short sulcus on interocular space deep (
Figure 8A); width of interocellar space longer than width of ocellus (
Figure 8A); first visible segment of rostrum shortest, third segment about 1.12 times longer than first segment (
Figure 8B). Pronotum with anterior margin nearly straight, collar process prominent; anterior lobe of pronotum about 1.85 times longer than posterior lobe; lateral pronotal angle rounded (
Figure 8A). Fore tibia with fossula spongiosa occupying half of tibial length (
Figure 8D), mid tibia with fossula spongiosa occupying 1/3 of tibial length (
Figure 8E).
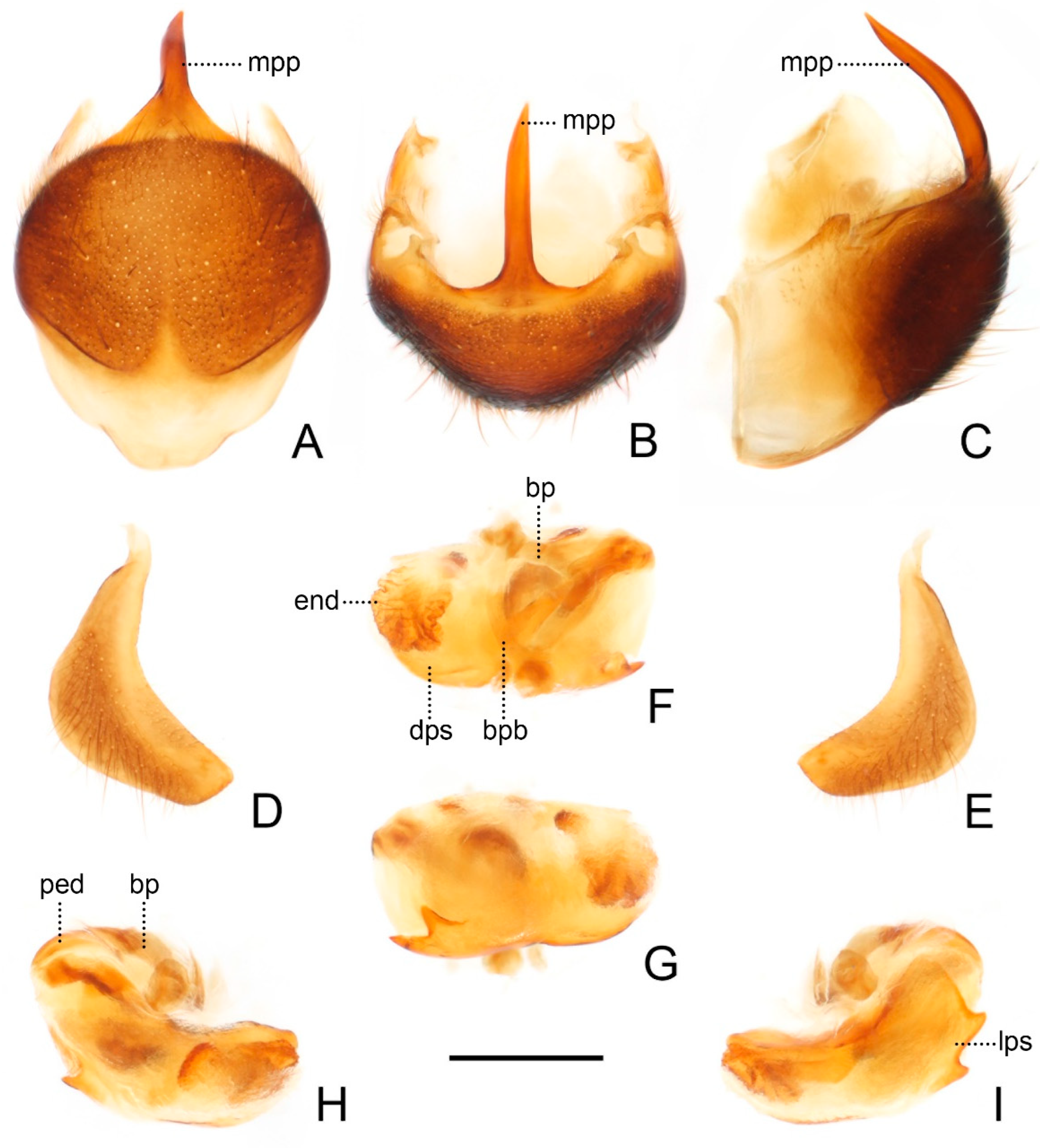
Male genitalia asymmetric (
Figure 9). Pygophore oval in ventral view (
Figure 9A); median pygophore process long and extremely slender, apex sharp (
Figure 9A–C), nearly vertical in caudal view (
Figure 9B), curved in lateral view (
Figure 9C). Parameres sickle-shaped, left paramere (
Figure 9D) slightly longer than right paramere (
Figure 9E). Phallus with phallobase feebly sclerotized, basal plate bridge feebly curved and slightly longer than basal plate (
Figure 9F); pedicel slightly curved and longer than basal plate (
Figure 9H,I). Dorsal phallothecal sclerite slightly sclerotized, broad and flat (
Figure 9F, H and I); lateral phallothecal sclerite moderately sclerotized, lower half of inner margin with two upward processes (
Figure 9I). Apical portion of endosoma with a somewhat petaloid process, surface of which is covered with tiny tubercles (
Figure 9F,G).
Figure 8.
Sigicoris dominiqueae sp. nov., paratype male. (A) Anterior part of body with antennae and legs removed, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Left fore leg, ventral view. (E) Right mid leg, ventral view. (F) Right hemelytron, dorsal view. (G) Abdomen, ventral view; (H) ditto, caudal view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Figure 8.
Sigicoris dominiqueae sp. nov., paratype male. (A) Anterior part of body with antennae and legs removed, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Left fore leg, ventral view. (E) Right mid leg, ventral view. (F) Right hemelytron, dorsal view. (G) Abdomen, ventral view; (H) ditto, caudal view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Measurements [in mm, male (n = 1), female (n = 2)]: Body length 10.56 (male), 10.65–10.70 (female); maximum width of abdomen 3.18 (male), 3.32–3.41 (female); head length 1.62 (male), 1.61–1.60 (female); length of anteocular part 0.78 (male), 0.73–0.78 (female); length of postocular part 0.31 (male), 0.31–0.24 (female); head width 1.15 (male), 1.08–1.12 (female); eye width in dorsal view 0.35 (male), 0.30–0.32 (female); width of interocular space 0.44 (male), 0.48–0.47 (female); width of interocellar space 0.16 (male), 0.12–0.19 (female); lengths of rostral segments I:II:III= 0.57: 0.97: 0.65 (male), 0.58–0.50: 1.01–0.91: 0.68–0.52 (female); lengths of antennal segments I:II:III:IV = 0.94:?:?:? (male), 0.90–0.90:2.01–1.98:1.56–1.35:1.72–1.70 (female); length of anterior pronotal lobe 1.57 (male), 1.55–1.68 (female); length of posterior pronotal lobe 0.90 (male), 0.90–0.80 (female); width of anterior pronotal lobe 1.65 (male), 1.76–1.80 (female); width of posterior pronotal lobe 2.80 (male), 2.70–2.82 (female); scutellum length 1.24 (male), 1.22–1.11 (female); maximum width of scutellum 1.31 (male), 1.25–1.31 (female); hemelytron length 7.50 (male), 6.99–7.29 (female).
Figure 9.
Male genitalia of Sigicoris dominiqueae sp. nov., paratype. (A) Pygophore, ventral view; (B) ditto, caudal view; (C) ditto, lateral view. (D) Left paramere, outer ventrolateral view. (E) Right paramere, outer ventrolateral view. (F) Phallus, dorsal view; (G) ditto, ventral view; (H) ditto, left lateral view; (I) ditto, right lateral view. Scale bar = 0.50 mm. Abbreviations: bp, basal plate; bpb, basal plate bridge; dps, dorsal phallothecal sclerite; end, endosoma; lps, lateral phallothecal sclerite; mpp, median pygophore process; ped, pedicel.
Figure 9.
Male genitalia of Sigicoris dominiqueae sp. nov., paratype. (A) Pygophore, ventral view; (B) ditto, caudal view; (C) ditto, lateral view. (D) Left paramere, outer ventrolateral view. (E) Right paramere, outer ventrolateral view. (F) Phallus, dorsal view; (G) ditto, ventral view; (H) ditto, left lateral view; (I) ditto, right lateral view. Scale bar = 0.50 mm. Abbreviations: bp, basal plate; bpb, basal plate bridge; dps, dorsal phallothecal sclerite; end, endosoma; lps, lateral phallothecal sclerite; mpp, median pygophore process; ped, pedicel.
Remarks: This new species is similar to S. brumalis comb. nov. and S. gracilis comb. nov., but it can be distinguished from those two species by the following characters: the body size is smaller (less than 10 mm in S. dominiqueae sp. nov. vs. over 12 mm in S. brumalis comb. nov. and S. gracilis comb. nov.), the pronotum is bicolored with the posterior lobe paler (vs. the pronotum unicolored in S. brumalis comb. nov. and S. gracilis comb. nov.), the first visible segment of rostrum is shorter than the third (vs. the first visible segment of rostrum longer than the third in S. brumalis comb. nov. and S. gracilis comb. nov.) and the fossula spongiosa is less prominent (the fore tibia with the fossula spongiosa occupying half of the tibial length, the mid tibia with the fossula spongiosa occupying 1/3 of the tibial length in S. dominiqueae sp. nov. vs. the fore tibia with the fossula spongiosa occupying 2/3 of the tibial length, the mid tibia with the fossula spongiosa occupying half of the tibial length in S. brumalis comb. nov. and S. gracilis comb. nov.).
Ectomocoris (Sigicoris) gracilis Miller, 1958: 74–75. Central North New Guinea, Sigi camp.
Ectomocoris gracilis: Maldonado-Capriles, 1990: 353.
Type material examined: Indonesia: Paratype, 1 female, yellow-margined “Paratype” disc // “Neth. Ind.-Amer. New Guinea Exp. Iebele Camp 1938 2250 mg xi—L. J. Toxopeus leg.” // “Sigicoris gracilis subgen. n., sp. n. (paratype) N. C. E. Miller det. 1956” // “NHMUK 013585907” (NHM); Paratype, 1 male, yellow-margined “Paratype” disc // “Neth. Ind.-American New Guinea Exped. Sigi Camp 1500 m 25 iS. 1939 L. J. Toxopeus” // “Sigicoris gracilis subgen. n., sp. n. (paratype) N. C. E. Miller det. 1956” // “NHMUK 013585908” (NHM); Paratypes, 2 females, yellow-margined “Paratype” disc // “North New Guinea, Hollandia, 140° E. Long. 3°10′ S., 300–600 m, W.S.” // “Sigicoris gracilis subgen. n., sp. n. (paratype) N. C. E. Miller det. 1956” // “B.M. 1938-461” // “NHMUK 013585909” & “NHMUK 013585910” (NHM).
Distribution: Indonesia (Papua).
Diagnosis: Body brown with head, pronotum, scutellum, pleura and sterna dark brown; width of interocellar space subequal to width of ocellus; first visible segment of rostrum longer than third segment; pronotum with anterior margin nearly straight, collar process less prominent, lateral pronotal angle narrowly rounded; fore tibia with fossula spongiosa occupying 2/3 of tibial length, mid tibia with fossula spongiosa occupying half of tibial length.
Redescription: Macropterous form (
Figure 10 and
Figure 11).
Coloration. Brown. Head, pronotum, scutellum, pleura and sterna dark brown; third and fourth segments of antenna and tarsi yellowish brown (
Figure 10 and
Figure 11); hemelytron brown, with most of clavus, area between Pcu and Cu on corium, most base and marginal area of membrane paler, base of costal area of membrane with a pale brown securiform stripe (
Figure 12D).
Structure. Antennae, head, disc of scutellum, pleura, sterna, coxae, ventral surfaces and apex of femora, tibiae, lateral margin of corium and venter of abdomen densely covered with golden, procumbent, short pubescence; first and second antennal segments, lateral margins of head and pronotum, femora and tibiae covered with yellowish brown, suberect to erect setae of varying lengths; apex of dorsal surfaces of tibiae and ventral surfaces of tarsi densely covered with golden, suberect setae; venter of abdomen also sparsely covered with golden, long pubescence (
Figure 10 and
Figure 11).
Head length is 1.24 times as long as width in male and about 1.36 times as long as width in female; median short sulcus on interocular space deep (
Figure 12A); width of interocellar space subequal to width of ocellus (
Figure 12A); first visible segment of rostrum about 1.37 times longer than third segment (
Figure 12C). Pronotum with anterior margin nearly straight, collar process less prominent (
Figure 12A); anterior lobe of pronotum 1.58 times longer than posterior lobe in male and about 1.45 times longer than posterior lobe in female; lateral pronotal angle narrowly rounded (
Figure 12A). Fore tibia with fossula spongiosa occupying 2/3 of tibial length (
Figure 12E), mid tibia with fossula spongiosa occupying half of tibial length (
Figure 12F).
Figure 10.
Sigicoris gracilis comb. nov., paratype male, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Figure 10.
Sigicoris gracilis comb. nov., paratype male, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Measurements [in mm, male (n = 1), female (n = 3)]: Body length 12.19 (male), 14.22–15.36 (female); maximum width of abdomen 3.34 (male), 3.35–4.31 (female); head length 1.59 (male), 1.92–1.93 (female); length of anteocular part 0.70 (male), 0.91–0.92 (female); length of postocular part 0.37 (male), 0.49–0.49 (female); head width 1.28 (male), 1.35–1.50 (female); eye width in dorsal view 0.38 (male), 0.39–0.48 (female); width of interocular space 0.50 (male), 0.59–0.54 (female); width of interocellar space 0.10 (male), 0.18–0.20 (female); lengths of rostral segments I:II:III = 0.76:1.37:0.53 (male), 0.94–1.00:1.52–1.55:0.72–0.73 (female); lengths of antennal segments I:II:III:IV = 1.35:2.50:?:? (male), 1.29–1.38:2.67–2.89:2.13–?:2.51–? (female); length of anterior pronotal lobe 1.58 (male), 1.89–2.05 (female); length of posterior pronotal lobe 1.00 (male), 1.30–1.42 (female); width of anterior pronotal lobe 1.76 (male), 2.11–2.30 (female); width of posterior pronotal lobe 2.91 (male), 3.53–3.81 (female); scutellum length 1.40 (male), 1.29–? (female); maximum width of scutellum 1.52 (male), 1.35–1.85 (female); hemelytron length 8.70 (male), 10.42–10.52 (female).
Figure 11.
Sigicoris gracilis comb. nov., paratype female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Figure 11.
Sigicoris gracilis comb. nov., paratype female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Remarks: The holotype of this species is deposited in Rijksmuseum van Natuurlijke Historie, Leiden. We only examined four paratypes of this species preserved in NHM and there is no additional specimen for dissection and DNA extraction. This species is very allied to
S. brumalis comb. nov.; the characters currently used to separate it from the latter are the smaller body size, the paler coloration, the relatively larger ocellus, the straighter anterior margin of the pronotum with the less prominent collar process and the differences in the male genitalia as Miller illustrated [
23]. Here, we still treat them as two valid species, but more evidence such as molecular data and carefully dissected structures of male genitalia are needed to further clarify the relationship between them.
Figure 12.
Sigicoris gracilis comb. nov., paratypes. (A) Anterior part of body with antennae and legs removed, female, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Hemelytra, female, dorsal view. (E) Left fore leg without tarsus, female, ventral view. (F) Right mid tibia, female, ventral view. (G) Abdomen, female, ventral view. (H) Abdomen, male, ventral view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Figure 12.
Sigicoris gracilis comb. nov., paratypes. (A) Anterior part of body with antennae and legs removed, female, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Hemelytra, female, dorsal view. (E) Left fore leg without tarsus, female, ventral view. (F) Right mid tibia, female, ventral view. (G) Abdomen, female, ventral view. (H) Abdomen, male, ventral view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Brachysandalus sexguttatus Stål, 1866: 261; Maldonado-Capriles, 1990: 345. Insula Mysol (Indonesia: Misool Island).
Pirates sexguttatus: Walker, 1873: 123.
Pirates concinnus Walker, 1873: 124. New Guinea. syn. nov.
Pirates (
Brachysandalus)
sexguttatus: Stål, 1874: 60. [
34]
Peirates concinnus: Maldonado-Capriles, 1990: 364; Coscarón, 1997: 39, 41, excluded from Peirates without giving a new status.
Type material examined: Indonesia: Lectotype of Brachysandalus sexguttatus (designated by present study), male, “Typus” red rectangle label // “Stevens.” // “Mysol” // “sexguttatus Stål” // “NHRS-GULI 000000135” (NHRS); Holotype of Pirates concinnus, male, red-margined “Holotype” disc // green-margined “Type” disc // “N” disc // “Saunders. 65·13.” // “93. PIRATES CONCINNUS.” // “NHMUK 013587668” (NHM).
Other material examined: Indonesia: 1 ex (without abdomen), “Charles Lewis Mt. New Guinea. 98-203.” // “NHMUK 013587669” (NHM); 1 female, “Indonesia, West Papua ARFAK MTS, 1190 m alt DUEBEI ENV, 21. 1-8.2.2008 cca 20 km S of Warmere Manokwari distr, St Jakl lgt” (CAU).
Distribution: Indonesia (West Papua, Papua).
Figure 13.
Sigicoris sexguttatus comb. nov., lectotype male, habitus. (A) Dorsal view. (B) Lateral view. (C) Ventral view. Scale bar = 3.00 mm.
Figure 13.
Sigicoris sexguttatus comb. nov., lectotype male, habitus. (A) Dorsal view. (B) Lateral view. (C) Ventral view. Scale bar = 3.00 mm.
Diagnosis: Body dark brown, base of mid and hind tibiae and basal 1/3 of mid and hind femora yellowish white to pale yellow, hemelytron dark brown with three yellowish white spots, segment of connexivum with basal half yellowish white to yellow and apical half dark brown; width of interocellar space subequal to width of ocellus; first visible segment of rostrum longer than third segment; pronotum with anterior margin nearly straight, collar process prominent, lateral pronotal angle rounded; fore tibia with fossula spongiosa occupying half of tibial length, mid tibia with fossula spongiosa occupying 2/5 of tibial length.
Redescription: Macropterous form (
Figure 13,
Figure 14 and
Figure 15).
Coloration. Dark brown. Head, anterior lobe of pronotum, scutellum, pleura and sterna blackish brown; antennae brown; base of mid and hind tibiae and basal 1/3 of mid and hind femora yellowish white to pale yellow, tarsi yellowish brown (
Figure 13,
Figure 14 and
Figure 15); hemelytron dark brown except area between Pcu and Cu on corium and apical portion of membrane paler, three yellowish white spots present on hemelytron: a trapezoidal spot on conjunctive area of clavus, corium and membrane, a small, narrow, triangular spot on base of area between Sc and M on corium and a somewhat rectangular spot on base of costal area of membrane (
Figure 16F); segment of connexivum with basal half yellowish white to yellow and apical half dark brown (
Figure 13,
Figure 14,
Figure 15 and
Figure 16G).
Figure 14.
Sigicoris sexguttatus comb. nov., holotype of Pirates concinnus, male. (A) Habitus in dorsal view. (B) Habitus in ventral view. (C) Habitus in lateral view. (D) Legs stuck on the card. Scale bar = 3.00 mm.
Figure 14.
Sigicoris sexguttatus comb. nov., holotype of Pirates concinnus, male. (A) Habitus in dorsal view. (B) Habitus in ventral view. (C) Habitus in lateral view. (D) Legs stuck on the card. Scale bar = 3.00 mm.
Structure. Antennae, marginal area of head, disc of scutellum, coxal cavities, coxae, ventral surfaces and apex of femora, tibiae, lateral margin of corium and genitalic part of abdomen densely covered with golden, procumbent, short pubescence; first and second antennal segments, lateral margins of head and pronotum, femora and tibiae covered with brown, suberect to erect setae of varying lengths; apex of dorsal surfaces of tibiae and ventral surfaces of tarsi densely covered with yellow to yellowish brown, suberect setae; venter of abdomen sparsely covered with golden, long pubescence (
Figure 13,
Figure 14 and
Figure 15).
Head length is 1.21 times as long as width in male and 1.28 times as long as width in female; median short sulcus on interocular space deep (
Figure 16A); width of interocellar space subequal to width of ocellus (
Figure 16A); first visible segment of rostrum about 1.18 times longer than third segment (
Figure 16B). Pronotum with anterior margin nearly straight, collar process prominent (
Figure 16A); anterior lobe of pronotum 1.62 times longer than posterior lobe in male and 1.39 times longer than posterior lobe in female, lateral pronotal angle rounded (
Figure 16A). Fore tibia with fossula spongiosa occupying half of tibial length (
Figure 16D), mid tibia with fossula spongiosa occupying 2/5 of tibial length (
Figure 16E); hemelytron with Pcu and Cu on membrane slightly curved, not so straight as those in other species of
Sigicoris stat. nov. (
Figure 16F).
Figure 15.
Sigicoris sexguttatus comb. nov., female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Figure 15.
Sigicoris sexguttatus comb. nov., female, habitus. (A) Dorsal view. (B) Ventral view. (C) Lateral view. Scale bar = 3.00 mm.
Measurements [in mm, male (n = 1), female (n = 1)]: Body length 8.16 (male), 10.58 (female); maximum width of abdomen 2.44 (male), 3.31 (female); head length 1.22 (male), 1.59 (female); length of anteocular part 0.59 (male), 0.69 (female); length of postocular part 0.28 (male), 0.35 (female); head width 1.00 (male), 1.24 (female); eye width in dorsal view 0.29 (male), 0.35 (female); width of interocular space 0.41 (male), 0.55 (female); width of interocellar space 0.11 (male), 0.18 (female); lengths of rostral segments I:II:III = 0.52:0.91:0.49 (male), 0.79:0.95:0.61 (female); lengths of antennal segments I:II:III:IV = 0.73:1.64:?:? (male), 0.89:1.91:1.35:1.09 (female); length of anterior pronotal lobe 1.38 (male), 1.45 (female); length of posterior pronotal lobe 0.85 (male), 1.04 (female); width of anterior pronotal lobe 1.63 (male), 1.85 (female); width of posterior pronotal lobe 2.40 (male), 2.93 (female); scutellum length 0.88 (male), 0.98 (female); maximum width of scutellum 1.15 (male), 1.35 (female); hemelytron length 6.01 (male), 7.11 (female).
Figure 16.
Sigicoris sexguttatus comb. nov., female. (A) Anterior part of body with antennae and legs removed, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Left fore leg, ventral view. (E) Right mid leg, ventral view. (F) Right hemelytron, dorsal view. (G) Abdomen, ventral view; (H) ditto, caudal view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Figure 16.
Sigicoris sexguttatus comb. nov., female. (A) Anterior part of body with antennae and legs removed, dorsal view; (B) ditto, lateral view; (C) ditto, ventral view. (D) Left fore leg, ventral view. (E) Right mid leg, ventral view. (F) Right hemelytron, dorsal view. (G) Abdomen, ventral view; (H) ditto, caudal view. Scale bar = 2.00 mm. Abbreviations: 1A, first analis; Cu, cubitus; M, media; Pcu, postocubitus; R, radius; Sc, subcosta; IC, inner cell on membrane; OC, outer cell on membrane.
Remarks: The original data of the examined specimens of
Brachysandalus sexguttatus are “♂, Long. 8, Lat. 2 mill. Patria: Insula Mysol. (Mus. Holm.)”. Stål did not designate the holotype and did not mention the number of specimens he examined [
35]. We only found one male syntype of this species in NHRS and its label information matches the original data (
Figure 13). Here, we designate this specimen as the lectotype of
B. sexguttatus.
This species can be easily distinguished from other species of Sigicoris stat. nov. by the slightly curved Pcu and Cu on the membrane, the yellowish white spots on the hemelytron and the basal 1/3 of mid and hind femora yellowish white.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
















