


Biological Parameters, Phenology and Temperature Requirements of Halyomorpha halys (Hemiptera: Pentatomidae) in the Sub-Mediterranean Climate of Western Slovenia




Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Phenology, Lifetime Fecundity and Mortality Assessment in Outdoor Conditions
2.2. Life-Tables Analyses
2.3. Data Analysis
3. Results
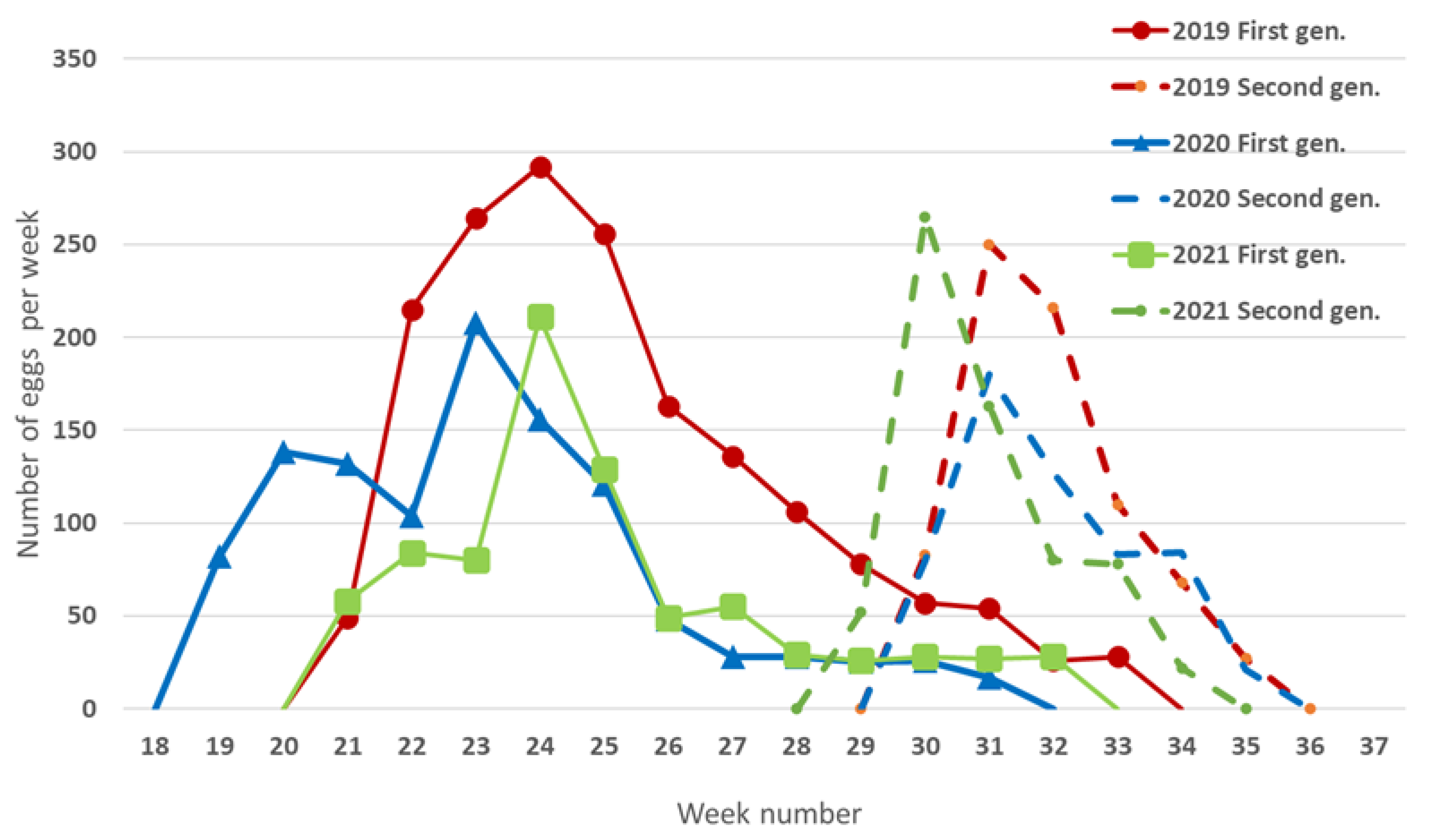
3.1. Overwintering Survival and Seasonal Phenology
3.2. Egg to Adult Development under a Natural Fluctuating Temperature Regime
3.3. Life Table Parameters for Halyomorpha Halys Population in 2019
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maistrello, L.; Dioli, P.; Dutto, M.; Volani, S.; Pasquali, S.; Gilioli, G. Tracking the Spread of Sneaking Aliens by Integrating Crowdsourcing and Spatial Modeling: The Italian Invasion of Halyomorpha Halys. BioScience 2018, 68, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Hoebeke, E.; Carter, M.E. Halyomorpha halys (Stål) (Heteroptera: Pentatomidae): A Polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2003, 105, 225–237. [Google Scholar]
- Yan, J.; Pal, C.; Anderson, D.; Vétek, G.; Farkas, P.; Burne, A.; Fan, Q.-H.; Zhang, J.; Gunawardana, D.N.; Balan, R.K.; et al. Genetic Diversity Analysis of Brown Marmorated Stink Bug, Halyomorpha halys Based on Mitochondrial COI and COII Haplotypes. BMC Genom. Data 2021, 22, 7. [Google Scholar] [CrossRef]
- Wermelinger, B.; Forster, B.; Wyniger, D. First records of an invasive bug in Europe: Halyomorpha halys Stål (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitt. Schweiz. Entomol. 2008, 81, 1–8. [Google Scholar]
- Cesari, M.; Maistrello, L.; Piemontese, L.; Bonini, R.; Dioli, P.; Lee, W.; Park, C.-G.; Partsinevelos, G.K.; Rebecchi, L.; Guidetti, R. Genetic Diversity of the Brown Marmorated Stink Bug Halyomorpha halys in the Invaded Territories of Europe and Its Patterns of Diffusion in Italy. Biol. Invas. 2018, 20, 1073–1092. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Musolin, D.L.; Konjević, A.; Karpun, N.N.; Zakharchenko, V.Y.; Zhuravleva, E.N.; Tavella, L.; Bruin, A.; Haye, T. Diversity and Distribution of Cytochrome Oxidase I (COI) Haplotypes of the Brown Marmorated Stink Bug, Halyomorpha halys Stål (Hemiptera, Pentatomidae), along the Eastern Front of Its Invasive Range in Eurasia. NeoBiota 2021, 68, 53–77. [Google Scholar] [CrossRef]
- Faúndez, E.; Rider, D. The Brown Marmorated Stink Bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) in Chile. Arquivos Entomol. 2017, 17, 305–307. [Google Scholar]
- Van der Heyden, T.; Saci, A.; Dioli, P. First recordof the brown marmorated stink bug Halyomorpha halys (Stål, 1855) in Algeria and its presence in North Africa (Heteroptera: Pentatomidae). Rev. Gadit. Entomol. 2021, 12, 147–154. [Google Scholar]
- Bergmann, E.J.; Venugopal, P.D.; Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Host plant use by the Invasive Halyomorpha halys (Stål) on woody ornamental trees and shrubs. PLoS ONE 2016, 11, e0149975. [Google Scholar] [CrossRef]
- Ludwick, D.; Morrison, W.R.; Acebes-Doria, A.L.; Agnello, A.M.; Bergh, J.C.; Buffington, M.L.; Hamilton, G.C.; Harper, J.K.; Hoelmer, K.A.; Krawczyk, G.; et al. Invasion of the Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) into the United States: Developing a National Response to an Invasive Species Crisis Through Collaborative Research and Outreach Efforts. J. Integr. Pest Manag. 2020, 11, 4. [Google Scholar] [CrossRef]
- Leskey, T.C.; Short, B.D.; Butler, B.R.; Wright, S.E. Impact of the invasive brown marmorated stink bug, Halyomorpha halys (Stål), in mid-Atlantic tree fruit orchards in the United States: Case studies of commercial management. J. Entomol. 2012, 2012, 535062. [Google Scholar] [CrossRef] [Green Version]
- Kuhar, T.P.; Kamminga, K.L.; Whalen, J.; Dively, G.P.; Brust, G.; Hooks, C.R.R.; Hamilton, G.; Herbert, D.A. The pest potential of brown marmorated stink bug on vegetable crops. Plant Health Prog. 2012, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Maistrello, L.; Vaccari, G.; Caruso, S.; Costi, E.; Bortolini, S.; Macavei, L.; Foca, G.; Ulrici, A.; Bortolotti, P.; Nannini, R.; et al. Monitoring of the Invasive Halyomorpha halys, a New Key Pest of Fruit Orchards in Northern Italy. J. Pest Sci. 2017, 90, 1231–1244. [Google Scholar] [CrossRef]
- Bariselli, M.; Bugiani, R.; Maistrello, L. Distribution and damage caused by Halyomorpha halys in Italy. EPPO Bull. 2016, 46, 332–334. [Google Scholar] [CrossRef]
- Bosco, L.; Moraglio, S.T.; Tavella, L. Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J. Pest Sci. 2018, 91, 661–670. [Google Scholar] [CrossRef]
- CSO Italy. Estimation of Damage from Brown Marmorated Stink Bug and Plant Pathologies Related to Climate Change. 2020. Available online: www.csoservizi.com (accessed on 20 February 2021).
- Stoeckli, S.; Felber, R.; Haye, T. Current distribution and voltinism of the brown marmorated stink bug, Halyomorpha halys, in Switzerland and its response to climate change using a high-resolution CLIMEX model. Int. J. Biometeorol. 2020, 64, 2019–2032. [Google Scholar] [CrossRef]
- Vétek, G.; Korányi, D. Severe damage to vegetables by the invasive brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), in Hungary. Period. Biol. 2017, 119, 131–135. [Google Scholar] [CrossRef]
- Damos, P.; Soulopoulou, P.; Thomidis, T. First Record and Current Status of the Brown Marmorated Sting Bug Halyomorpha halys Damaging Peaches and Olives in Northern Greece. J. Plant Protect. Res. 2020, 60, 323–326. [Google Scholar] [CrossRef]
- Musolin, D.; Konjevic, A.; Karpun, N.; Protsenko, V.; Ayba, L.; Saulich, A. Invasive Brown Marmorated Stink Bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Serbia: History of Invasion, Range Expansion, Early Stages of Establishment, and First Records of Damage to Local Crops. Arthropod Plant Interact. 2018, 12, 517–529. [Google Scholar] [CrossRef]
- Murvanidze, M.; Krawczyk, G.; Inasaridze, N.; Dekanoidze, L.; Samsonadze, N.; Macharashvili, M.; Khutsishvili, S.; Shengelaia, S. Preliminary data on the biology of brown marmorated stink bug Halyomorpha halys (Hemiptera, Pentatomidae) in Georgia. Tur. J. Zool. 2018, 42, 6. [Google Scholar] [CrossRef]
- Rot, M.; Devetak, M.; Carlevaris, B.; Žežlina, J.; Žežlina, I. First record of brown marmorated stink bug (Halyomorpha halys Stål, 1855) (Hemiptera: Pentatomidae) in Slovenia. Acta Entomol. Slov. 2018, 26, 5–12. [Google Scholar]
- Rot, M.; Devetak, M.; Žigon, P.; Ferlež Rus, A.; Matko, B.; Peterlin, A. Marmorirana smrdljivka (Halyomorpha halys (Stål, 1855) [Hemiptera, Pentatomidae]; pojav in razširjenost nove invazivne, tujerodne stenice v Sloveniji. In Proceedings of the Zbornik Predavanj in Referatov 14. Slovenskega Posvetovanja o Varstvu Rastlin z Mednarodno Udeležbo, Maribor, Slovenia, 5–6 March 2019; Trdan, S., Ed.; Društvo za Varstvo Rastlin Slovenije: Ljubljana, Slovenia, 2019; pp. 134–141. (In Slovenian) [Google Scholar]
- Kistner, E.J. Climate Change Impacts on the Potential Distribution and Abundance of the Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) with Special Reference to North America and Europe. Environ. Entomol. 2017, 46, 1212–1224. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Tobin, P.C.; Nagarkatti, S.; Loeb, G.; Saunders, M.C. Historical and projected interactions between climate change and insect voltinism in a multivoltine species. Glob. Chang. Biol. 2008, 14, 951–957. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C. Seasonal occurrence and impact of Halyomorpha halys (Hemiptera: Pentatomidae) in tree fruit. J. Econ. Entomol. 2009, 102, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Bu, W.; Gao, Y.; Liu, G. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef]
- Haye, T.; Abdallah, S.; Gariepy, T.; Wyniger, D. Phenology, life table analysis and temperature requirements of the invasive brown marmorated stink bug, Halyomorpha halys, in Europe. J. Pest Sci. 2014, 87, 407–418. [Google Scholar] [CrossRef]
- Costi, E.; Haye, T.; Maistrello, L. Biological parameters of the invasive brown marmorated stink bug, Halyomorpha halys, in Southern Europe. J. Pest Sci. 2017, 90, 1059–1067. [Google Scholar] [CrossRef]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef]
- Nielsen, A.; Chen, S.; Fleischer, S. Coupling Developmental Physiology, Photoperiod, and Temperature to Model Phenology and Dynamics of an Invasive Heteropteran, Halyomorpha Halys. Front. Physiol. 2016, 7, 165. [Google Scholar] [CrossRef] [Green Version]
- Bakken, A.J.; Schoof, S.C.; Bickerton, M.; Kamminga, K.L.; Jenrette, J.C.; Malone, S.; Abney, M.A.; Herbert, D.A.; Reisig, D.; Kuhar, T.P.; et al. Occurrence of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) on Wild Hosts in Nonmanaged Woodlands and Soybean Fields in North Carolina and Virginia. ENVE 2015, 44, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Leskey, T.C.; Agnello, A.; Bergh, J.C.; Dively, G.P.; Hamilton, G.C.; Jentsch, P.; Khrimian, A.; Krawczyk, G.; Kuhar, T.P.; Lee, D.-H.; et al. Attraction of the Invasive Halyomorpha halys (Hemiptera: Pentatomidae) to Traps Baited with Semiochemical Stimuli across the United States. J. Econ. Entomol. 2015, 44, 746–756. [Google Scholar] [CrossRef]
- Ingels, C.A.; Daane, K.M. Phenology of Brown Marmorated Stink Bug in a California Urban Landscape. J. Econ. Entomol. 2018, 111, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Funayama, K. Nutritional Status of Overwintering Adults of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae), Is Affected by Cone Counts of Japanese Cedar, Cryptomeria Japonica, in Northern Japan. Appl. Entomol. Zool. 2014, 50, 117–121. [Google Scholar] [CrossRef]
- Medal, J.; Smith, T.; Fox, A.; Cruz, A.S.; Poplin, A.; Hodges, A. Rearing the Brown Marmorated Stink Bug Halyomorpha halys (Heteroptera: Pentatomidae). Fla. Entomol. 2012, 95, 800–802. [Google Scholar] [CrossRef]
- Bellows, T.S., Jr.; Van Driesche, R.G.; Elkinton, J.S. Life-table construction and analysis in the evaluation of natural enemies. Ann. Rev. Entomol. 1992, 37, 587–612. [Google Scholar] [CrossRef]
- Van Driesche, R.; Hoddle, M.; Center, T. 2008 Control of Pests and Weeds by Natural Enemies: An Introduction to Biological Control; Blackwell Publishing: Malden, MA, USA, 2008. [Google Scholar]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Gutierrez, A.P.; Mackauer, M. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar]
- Reznik, S.; Karpun, N.; Zakharchenko, V.; Shoshina, Y.; Dolgovskaya, M.; Saulich, A.; Musolin, D. To Every Thing There Is a Season: Phenology and Photoperiodic Control of Seasonal Development in the Invasive Caucasian Population of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae). Insects 2022, 13, 580. [Google Scholar] [CrossRef]
- Zhang, C.T.; Li, D.L.; Su, H.F.; Xu, G.L. A study on the biological characteristics of Halyomorpha picus and Erthesina fullo. For. Res. 1993, 6, 271–275. [Google Scholar]
- Hegazi, E.M.; Khafagi, W.E.; Agamy, E. Do non-diapausing insects respond to different photoperiods: Spodoptera littoralis? J. Entomol. Nematol. 2021, 13, 26–34. [Google Scholar]
- Saulich, A.K.; Musolin, D.L. Diapause in the seasonal cycle of stink bugs (Heteroptera: Pentatomidae) from the temperate zone. Entomol. Rev. 2012, 92, 1–26. [Google Scholar] [CrossRef]
- Musolin, D.L.; Dolgovskaya, M.Y.; Protsenko, V.Y.; Karpun, N.N.; Reznik, V.Y.; Saulich, A.K. Photoperiodic and temperature control of nymphal growth and adult diapause induction in the invasive Caucasian population of the brown marmorated stink bug, Halyomorpha halys. J. Pest Sci. 2019, 92, 621–631. [Google Scholar] [CrossRef]
- McDougall, R.N.; Ogburn, E.C.; Walgenbach, J.F.; Nielsen, A.L. Diapause Termination in Invasive Populations of the Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) in Response to Photoperiod. Environ. Entomol. 2021, 50, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Ogburn, E.C.; Ohmen, T.M.; Huseth, A.S.; Reisig, D.D.; Kennedy, G.G.; Walgenbach, J.F. Temperature-Driven Differences in Phenology and Habitat Suitability for Brown Marmorated Stink Bug, Halyomorpha halys, in Two Ecoregions of North Carolina. J. Pest Sci. 2022, 1–15. [Google Scholar] [CrossRef]
- Grisafi, F.; Papa, G.; Barbato, M.; Tombesi, S.; Negri, I. Influence of Microclimate Factors on Halyomorpha halys Dehydration. Insects 2021, 12, 897. [Google Scholar] [CrossRef]
- Scaccini, D.; Vanishvili, L.; Tirello, P.; Walton, V.M.; Duso, C.M.; Pozzebon, A. Lethal and sub-lethal effects of low-temperature exposures on Halyomorpha halys (Hemiptera: Pentatomidae) adults before and after overwintering. Sci. Rep. 2020, 10, 15231. [Google Scholar] [CrossRef]
- Funayama, K. Nutritional States of Post-Overwintering Adults of the Brown-Marmorated Stink Bug, Halyomorpha halys (Stål) (Heteroptera: Pentatomidae). Jap. J. Appl. Entomol. Zool. 2012, 56, 12–15. [Google Scholar] [CrossRef] [Green Version]
- Burne, A.R. Pest Risk Assessment: Halyomorpha Halys (Brown Marmorated Stink Bug); Version 1; Ministry for Primary Industries: Wellington, New Zealand, 2019. [Google Scholar]
- Kawada, H.; Kitamura, C. The reproductive behavior of the brown marmorated stink bug, Halyomorpha mista Uhler (Heteroptera: Pentatomidae). I. Observation of mating behavior and multiple copulation. Appl. Entomol. Zool. 1983, 18, 234–242. [Google Scholar] [CrossRef]
- Govindan, B.N.; Hutchison, W.D. Influence of Temperature on Age-Stage, Two-Sex Life Tables for a Minnesota-Acclimated Population of the Brown Marmorated Stink Bug (Halyomorpha halys). Insects 2020, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, A.L.; Hamilton, G.C.; Matadha, D. Developmental rate estimation and life table analysis for Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2008, 37, 348–355. [Google Scholar] [CrossRef]
- Johnson, C.A.; Coutinho, R.M.; Berlin, E.; Dolphin, K.E.; Heyer, J.; Kim, B.; Leung, A.; Sabellon, J.L.; Amarasekare, P. Effects of Temperature and Resource Variation on Insect Population Dynamics: The Bordered Plant Bug as a Case Study. Funct. Ecol. 2016, 30, 1122–1131. [Google Scholar] [CrossRef] [Green Version]
- Rot, M.; Maistrello, L.; Costi, E.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Trdan, S. Native and Non-Native Egg Parasitoids Associated with Brown Marmorated Stink Bug (Halyomorpha halys [Stål, 1855]; Hemiptera: Pentatomidae) in Western Slovenia. Insects 2021, 12, 505. [Google Scholar] [CrossRef]
- Yonow, T.; Kriticos, D.J.; Ota, N.; Avila, G.A.; Hoelmer, K.A.; Chen, H.; Caron, V. Modelling the Potential Geographic Distribution of Two Trissolcus Species for the Brown Marmorated Stink Bug, Halyomorpha Halys. Insects 2021, 12, 491. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Oviposition Period | Total No. of Eggs Laid Per Week (ex) | Proportion of Total No. Eggs Laid (%) (ex × 100/E) | No. of N2 Followed | % Mortality at Different Stages | % Total Mortality | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Eggs | N1 | N2 | N3 | N4 | N5 | |||||
| 27 May–2 June | 49 | 2.8 | 30 | 18.37 | 17.50 | 16.67 | 8.00 | 0.00 | 0.00 | 46.94 |
| 3–6 June | 215 | 12.5 | 40 | 11.82 | 11.34 | 20.00 | 6.25 | 6.67 | 7.69 | 25.07 |
| 10–16 June | 264 | 15.3 | 40 | 20.79 | 15.00 | 17.50 | 15.15 | 7.14 | 19.23 | 39.87 |
| 17–23 June | 292 | 16.9 | 40 | 16.36 | 26.37 | 30.00 | 17.86 | 4.35 | 4.55 | 44.93 |
| 24–30 June | 256 | 14.8 | 40 | 18.18 | 30.00 | 32.50 | 11.11 | 0.00 | 8.33 | 49.76 |
| 1–7 July | 163 | 9.5 | 40 | 14.29 | 8.33 | 20.00 | 6.25 | 3.33 | 13.79 | 30.63 |
| 8–14 July | 136 | 7.9 | 40 | 11.83 | 14.63 | 22.50 | 12.90 | 11.11 | 33.33 | 42.38 |
| 15–21 July | 106 | 6.1 | 40 | 14.15 | 17.58 | 30.00 | 39.29 | 0.00 | 0.00 | 50.94 |
| 22–28 July | 78 | 4.5 | 40 | 12.82 | 5.88 | 15.00 | 0.00 | 0.00 | 5.88 | 28.21 |
| 29 July–4 August | 57 | 3.3 | 30 | 15.79 | 20.83 | 10.00 | 0.00 | 3.70 | 3.85 | 42.11 |
| 5–11 August | 54 | 3.1 | 30 | 40.74 | 12.50 | 13.33 | 3.85 | 0.00 | 0.00 | 57.41 |
| 12–18 August | 26 | 1.5 | 18 | 30.77 | 0.00 | 55.56 | 12.50 | 0.00 | 14.29 | 76.92 |
| 19–25 August | 28 | 1.6 | 24 | 14.29 | 0.00 | 8.33 | 22.73 | 29.41 | 50.00 | 78.57 |
| Mean (SE) | 18.48 ± 2.32 | 13.84 ± 2.51 | 22.41 ± 3.47 | 11.99 ± 2.93 | 5.06 ± 2.37 | 12.38 ± 4.07 | ||||
| Total (E) | 1724 | |||||||||
| Oviposition Period | Total No. of Eggs Laid Per Week (ex) | Proportion of Total No. Eggs Laid (%) (ex × 100/E) | N2 Followed | % Mortality at Different Stages | % Total Mortality | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Eggs | N1 | N2 | N3 | N4 | N5 | |||||
| 29 July–4 August | 83 | 10.9 | 30 | 18.07 | 7.35 | 20.00 | 0.00 | 0.00 | 0.00 | 31.33 |
| 5–11 August | 231 | 30.4 | 40 | 7.27 | 24.51 | 35.00 | 11.54 | 13.04 | 5.00 | 37.79 |
| 12–18 August | 241 | 31.7 | 40 | 19.44 | 18.39 | 22.50 | 16.13 | 19.23 | 4.76 | 40.48 |
| 19–25 August | 110 | 14.5 | 40 | 9.09 | 4.00 | 20.00 | 21.88 | 8.00 | 56.52 | 33.64 |
| 26 August–1 September | 68 | 8.9 | 40 | 7.35 | 6.35 | 10.00 | 14.29 | 0.00 | 16.67 | 20.59 |
| 2–8 September | 27 | 3.6 | 16 | 25.93 | 20.00 | 12.50 | 100.00 | 100.00 | ||
| Mean (SE) | 14.53 ± 2.69 | 13.43 ± 3.50 | 20.00 ± 3.59 | 27.30 ± 14.84 | 8.05 ± 3.74 | 16.59 ± 10.35 | ||||
| Total (E) | 760 | |||||||||
| Year | The Length of Pre-Oviposition Period (Days) | Egg-To-Adult Development Time (Days) | ||||||
|---|---|---|---|---|---|---|---|---|
| Overwintering Generation | Mean Temp. (°C) | Summer Generation | Mean Temp. (°C) | Overwintering Generation | Mean Temp. (°C) | Summer Generation | Mean Temp. (°C) | |
| 2019 | 40.80 ± 1.59 | 13.6 | 17.30 ± 0.42 | 24.8 | 48.17 ± 0.42 | 23.8 | 59.23 ± 1.24 | 19.4 |
| 2020 | 33.20 ± 2.23 | 15.9 | 18.60 ± 0.58 | 23.8 | 59.33 ± 0.60 | 22.7 | 53.95 ± 0.90 | 20.8 |
| 2021 | 44.30 ± 1.14 | 13.3 | 14.00 ± 1.11 | 24.7 | 48.93 ± 0.59 | 24.1 | 54.48 ± 0.74 | 21.3 |
| Mean | 39.43 ± 3.28 | 16.63 ± 1.37 | 52.14 ± 3.60 | 55.89 ± 1.68 | ||||
| Year | Pre-Oviposition Period (DD12.2) | Egg-To-Adult Development (DD12.2) | ||
|---|---|---|---|---|
| Overwintering Generation | Summer Generation | Overwintering Generation | Summer Generation | |
| 2019 | 113.1 | 205.40 | 553.3 | 523.5 |
| 2020 | 130.2 | 183.9 | 510.5 | 534.2 |
| 2021 | 113.0 | 147.7 | 528.8 | 577.8 |
| Mean | 118.77 ± 7.72 | 179.00 ± 16.84 | 530.87 ± 12.40 | 545.17 ± 16.61 |
| Stage | lx | dx | Apparent Mortality qx (dx/lx) | Real Mortality rx (dx/l0) | k Value (−log (1 − qx)) | % of Generational Mortality (100 kx/Ks) |
|---|---|---|---|---|---|---|
| Egg | 1724.00 | 291.00 | 0.1688 | 0.1688 | 0.0803 | 19.66 |
| N1 | 1433.00 | 250.30 | 0.1747 | 0.1452 | 0.0834 | 20.42 |
| N2 | 1182.70 | 272.10 | 0.2301 | 0.1578 | 0.1135 | 27.81 |
| N3 | 910.60 | 110.49 | 0.1213 | 0.0641 | 0.0562 | 13.76 |
| N4 | 800.11 | 38.93 | 0.0487 | 0.0226 | 0.0217 | 5.31 |
| N5 | 761.17 | 87.84 | 0.1154 | 0.0510 | 0.0533 | 13.04 |
| New generation adults | 673.33 | |||||
| Sex ratio | 0.5 | |||||
| Adult females | 336.67 | |||||
| Realized fecundity | 76.0 | |||||
| Realized F1 progeny | 25,586.92 | |||||
| R0 | 14.84 | |||||
| Total mortality | 60.94% | Ks = 0.4084 |
| Stage | lx | dx | Apparent Mortality qx (dx/lx) | Real Mortality rx (dx/l0) | k Value (−log (1 − qx)) | % of Generational Mortality (100 kx/Ks) |
|---|---|---|---|---|---|---|
| Egg | 760.00 | 100.66 | 0.1324 | 0.1324 | 0.0617 | 6.21 |
| N1 | 659.34 | 105.20 | 0.1596 | 0.1384 | 0.0755 | 7.59 |
| N2 | 554.14 | 131.94 | 0.2381 | 0.1736 | 0.1181 | 11.88 |
| N3 | 422.19 | 70.32 | 0.1666 | 0.0925 | 0.0791 | 7.96 |
| N4 | 351.87 | 36.73 | 0.1044 | 0.0483 | 0.0479 | 4.82 |
| N5 | 315.14 | 46.79 | 0.1485 | 0.0616 | 0.0698 | 7.02 |
| Diapausing adults | 268.35 | 211.67 | 0.7129 | 0.2785 | 0.5420 | 54.52 |
| Overwintering adults | 77.04 | |||||
| Sex ratio | 0.5 | |||||
| Adult females | 38.52 | |||||
| Realized fecundity | 111,3 | |||||
| Realized F1 progeny | 4287.47 | |||||
| R0 | 5.64 | |||||
| Total mortality | 89.86 % | Ks = 0.9941 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rot, M.; Maistrello, L.; Costi, E.; Trdan, S. Biological Parameters, Phenology and Temperature Requirements of Halyomorpha halys (Hemiptera: Pentatomidae) in the Sub-Mediterranean Climate of Western Slovenia. Insects 2022, 13, 956. https://doi.org/10.3390/insects13100956
Rot M, Maistrello L, Costi E, Trdan S. Biological Parameters, Phenology and Temperature Requirements of Halyomorpha halys (Hemiptera: Pentatomidae) in the Sub-Mediterranean Climate of Western Slovenia. Insects. 2022; 13(10):956. https://doi.org/10.3390/insects13100956
Chicago/Turabian StyleRot, Mojca, Lara Maistrello, Elena Costi, and Stanislav Trdan. 2022. "Biological Parameters, Phenology and Temperature Requirements of Halyomorpha halys (Hemiptera: Pentatomidae) in the Sub-Mediterranean Climate of Western Slovenia" Insects 13, no. 10: 956. https://doi.org/10.3390/insects13100956
APA StyleRot, M., Maistrello, L., Costi, E., & Trdan, S. (2022). Biological Parameters, Phenology and Temperature Requirements of Halyomorpha halys (Hemiptera: Pentatomidae) in the Sub-Mediterranean Climate of Western Slovenia. Insects, 13(10), 956. https://doi.org/10.3390/insects13100956