

Prediction of the Current and Future Distributions of the Hessian Fly, Mayetiola destructor (Say), under Climatic Change in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
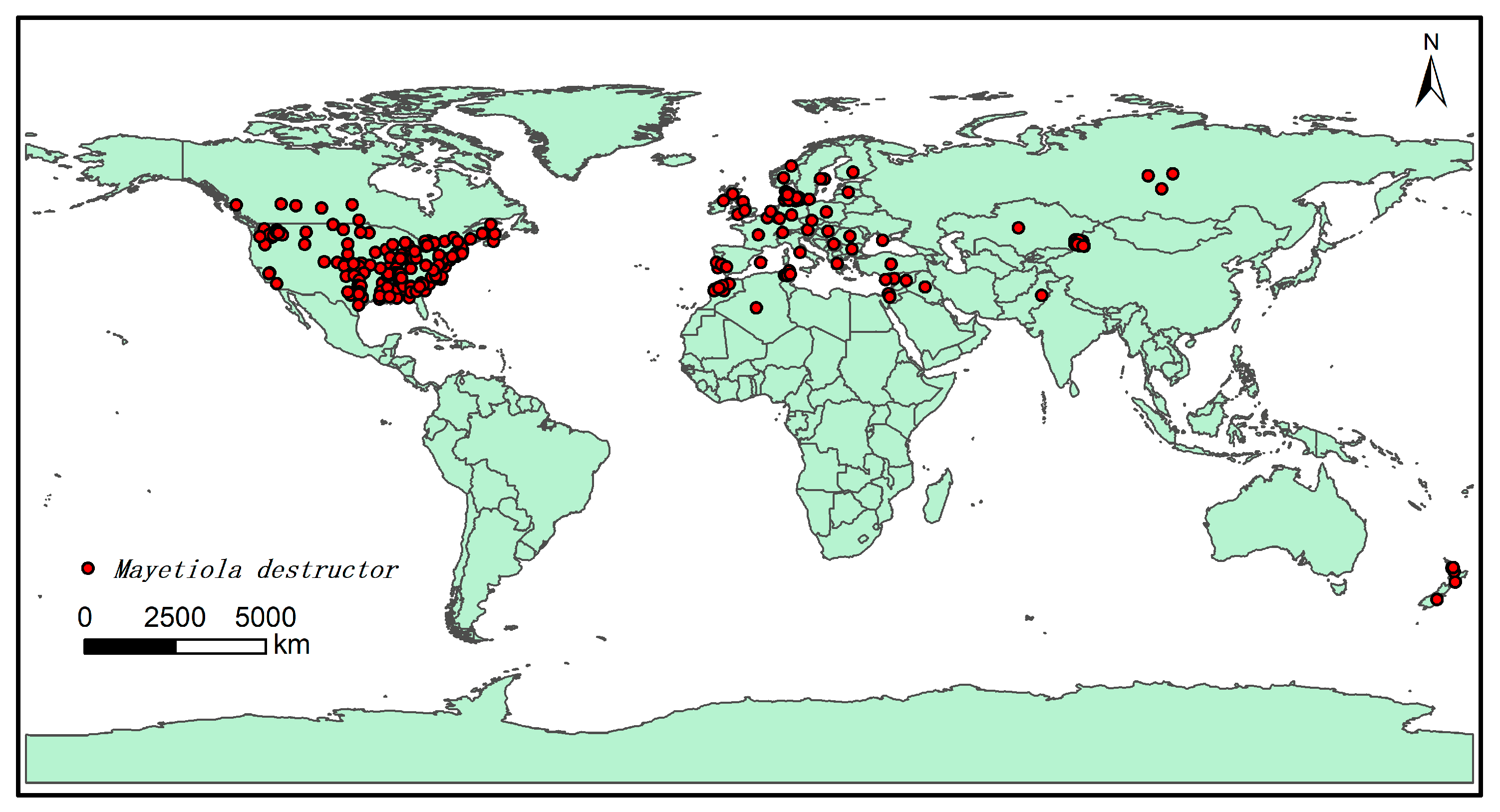
2.1. Data and Processing of Species Presence Records
2.2. Environmental Parameters
2.3. MaxEnt Model Construction and Parameter Optimization
3. Results
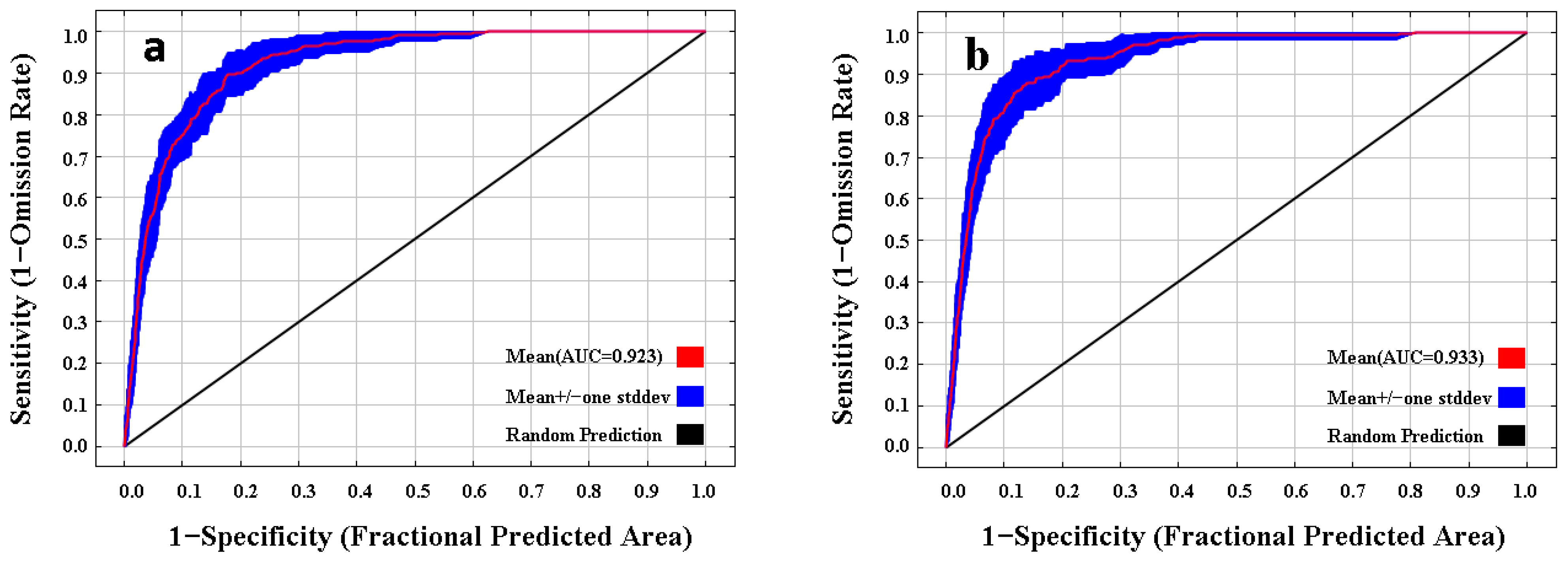
3.1. Model Performance
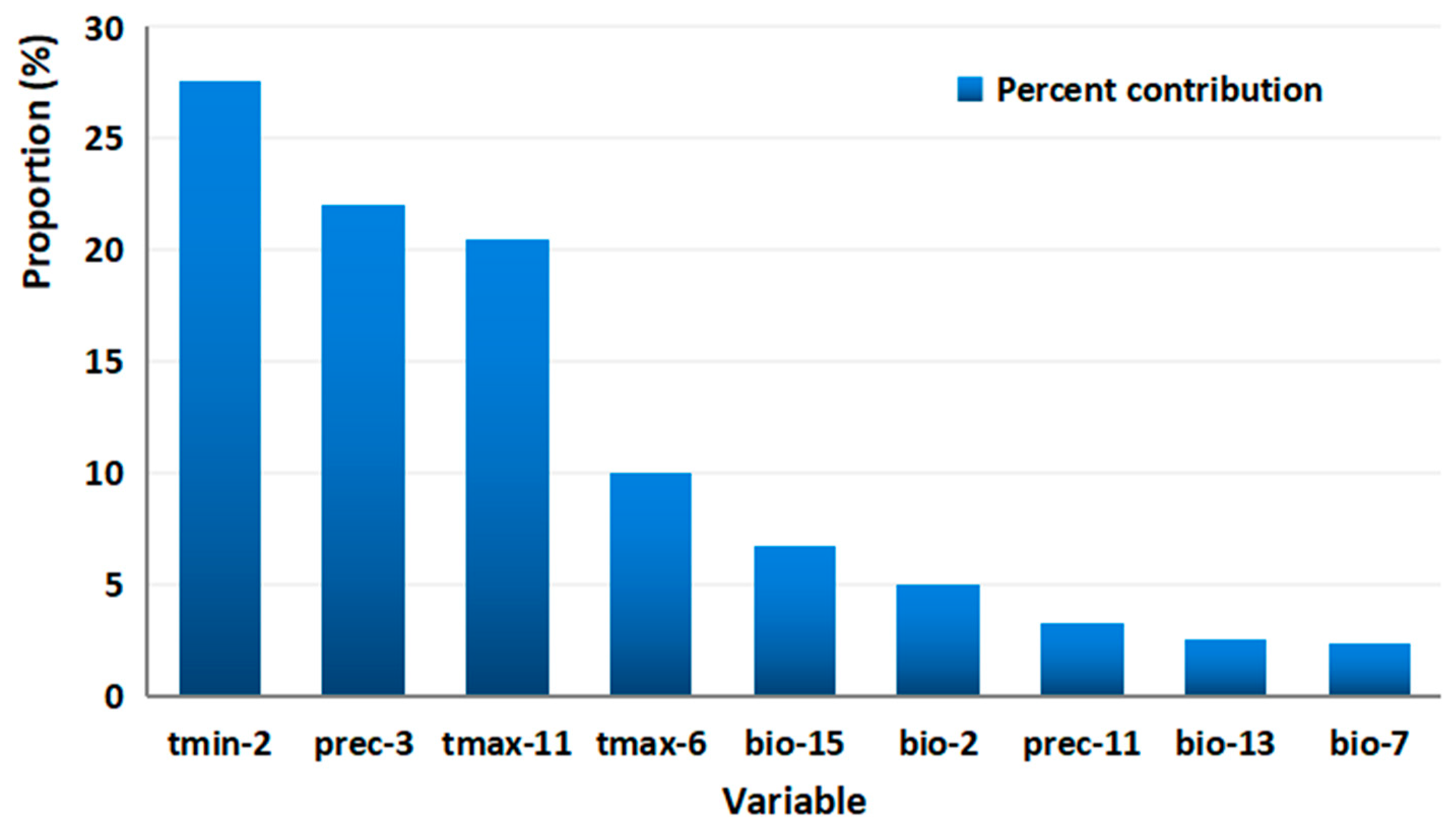
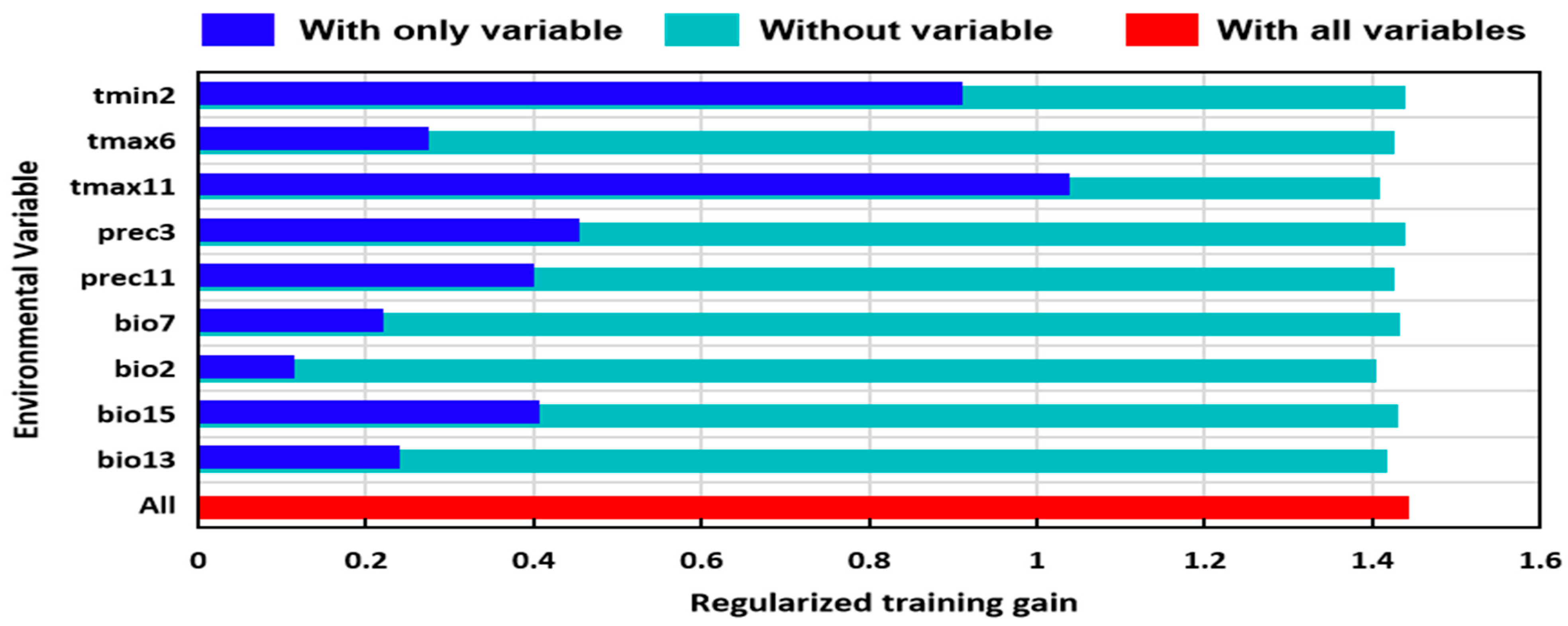
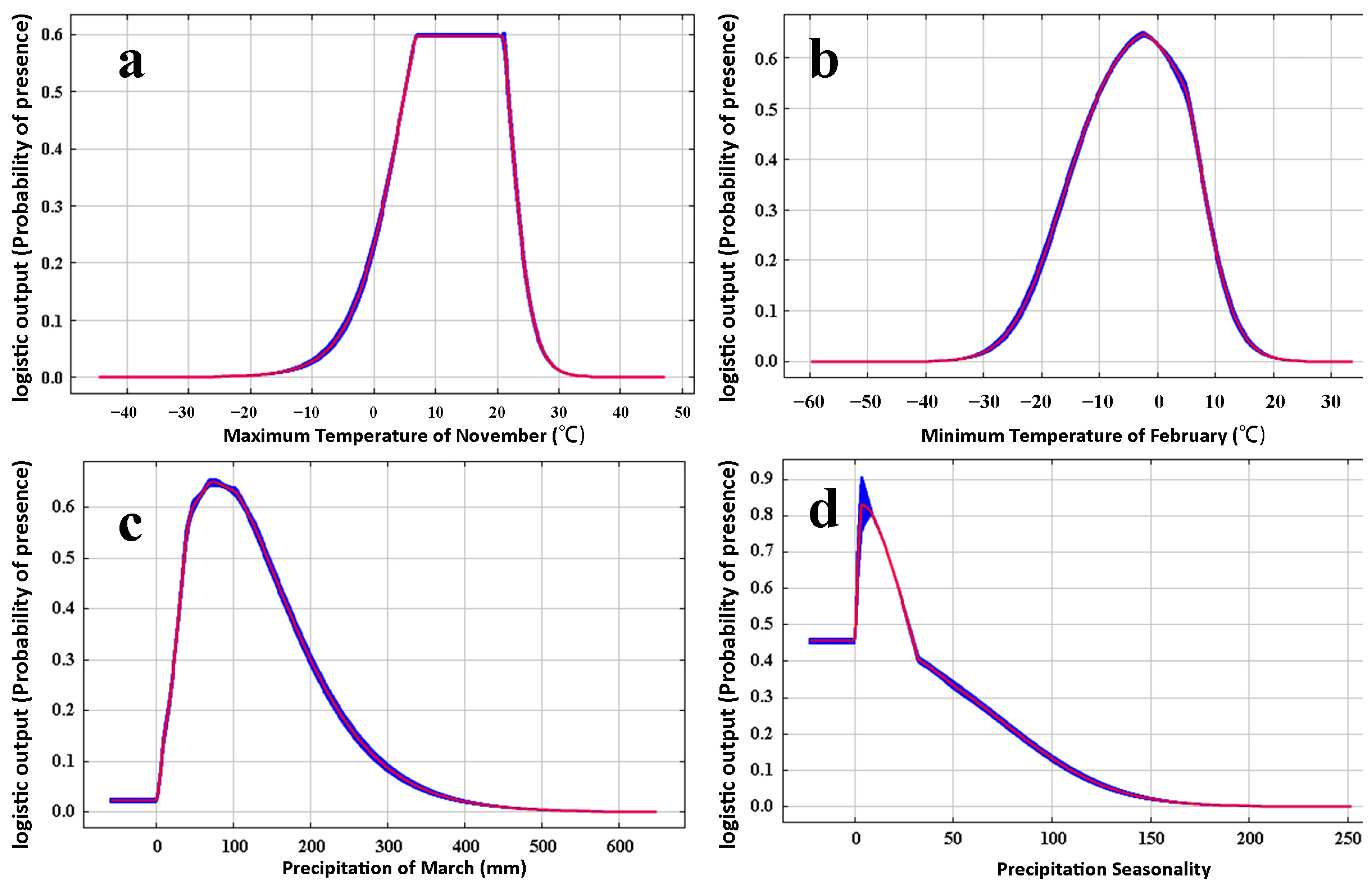
3.2. Dominant Environmental Factors Affecting Distribution
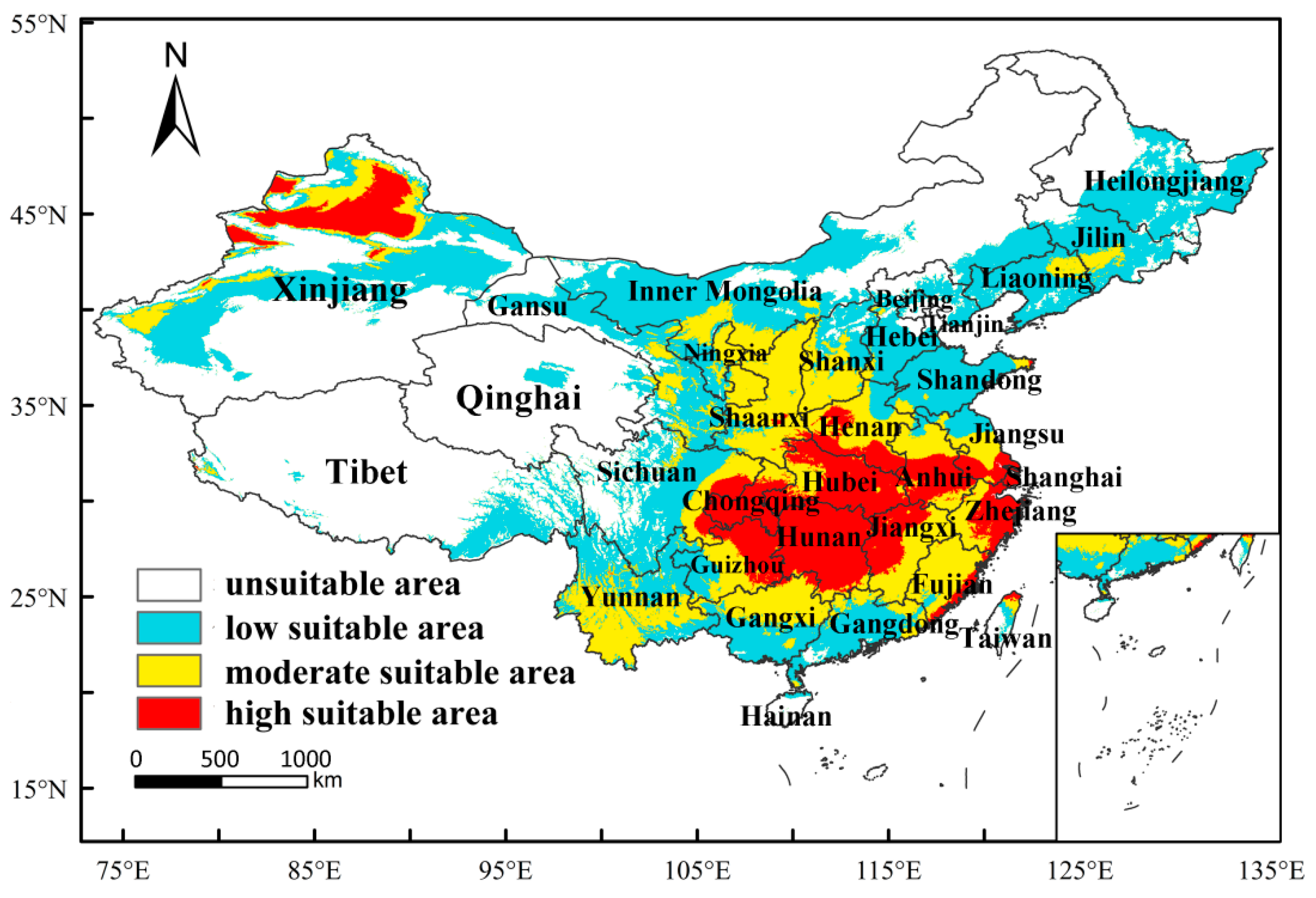
3.3. Predicting the Current Distribution of M. destructor in China
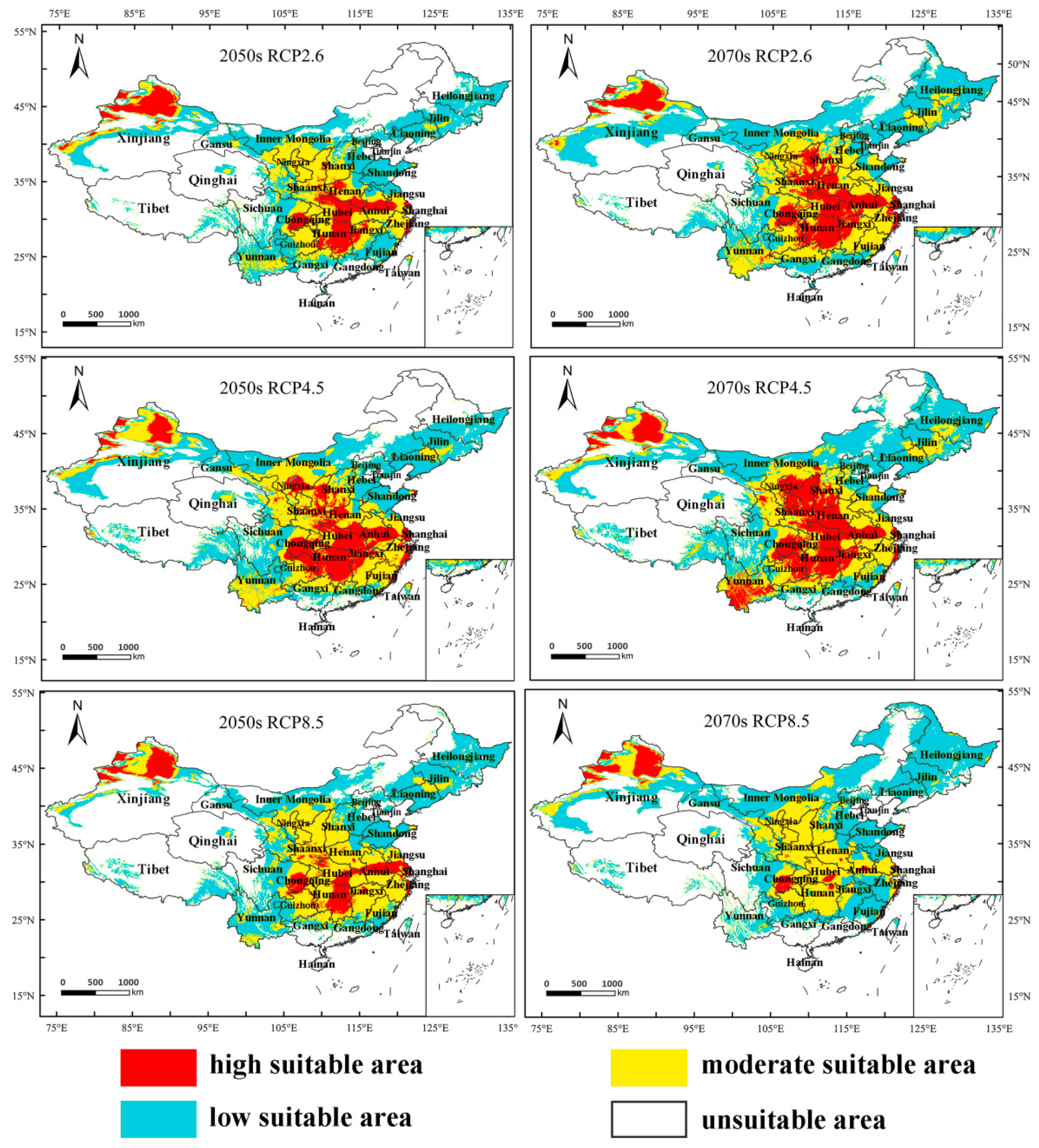
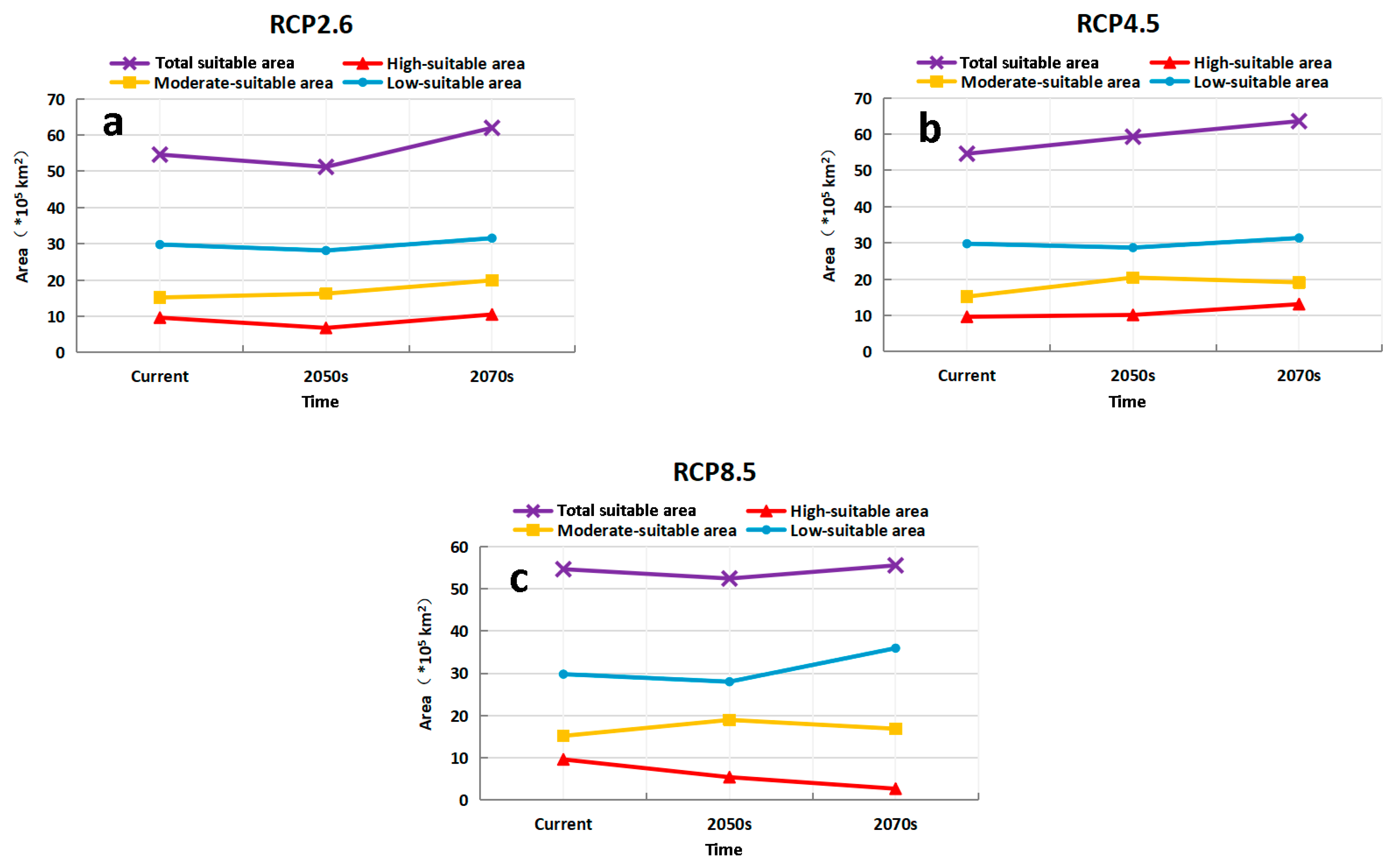
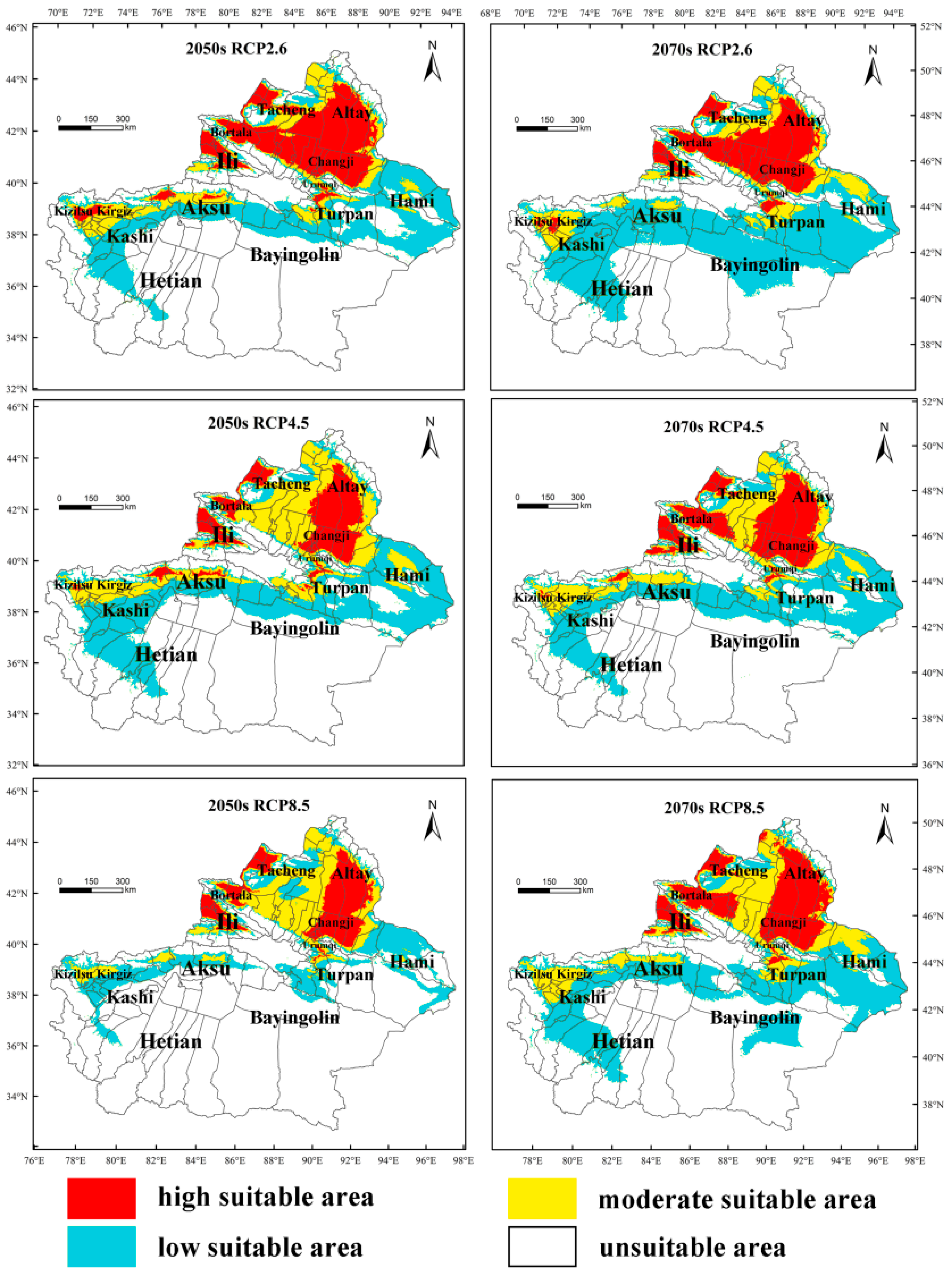
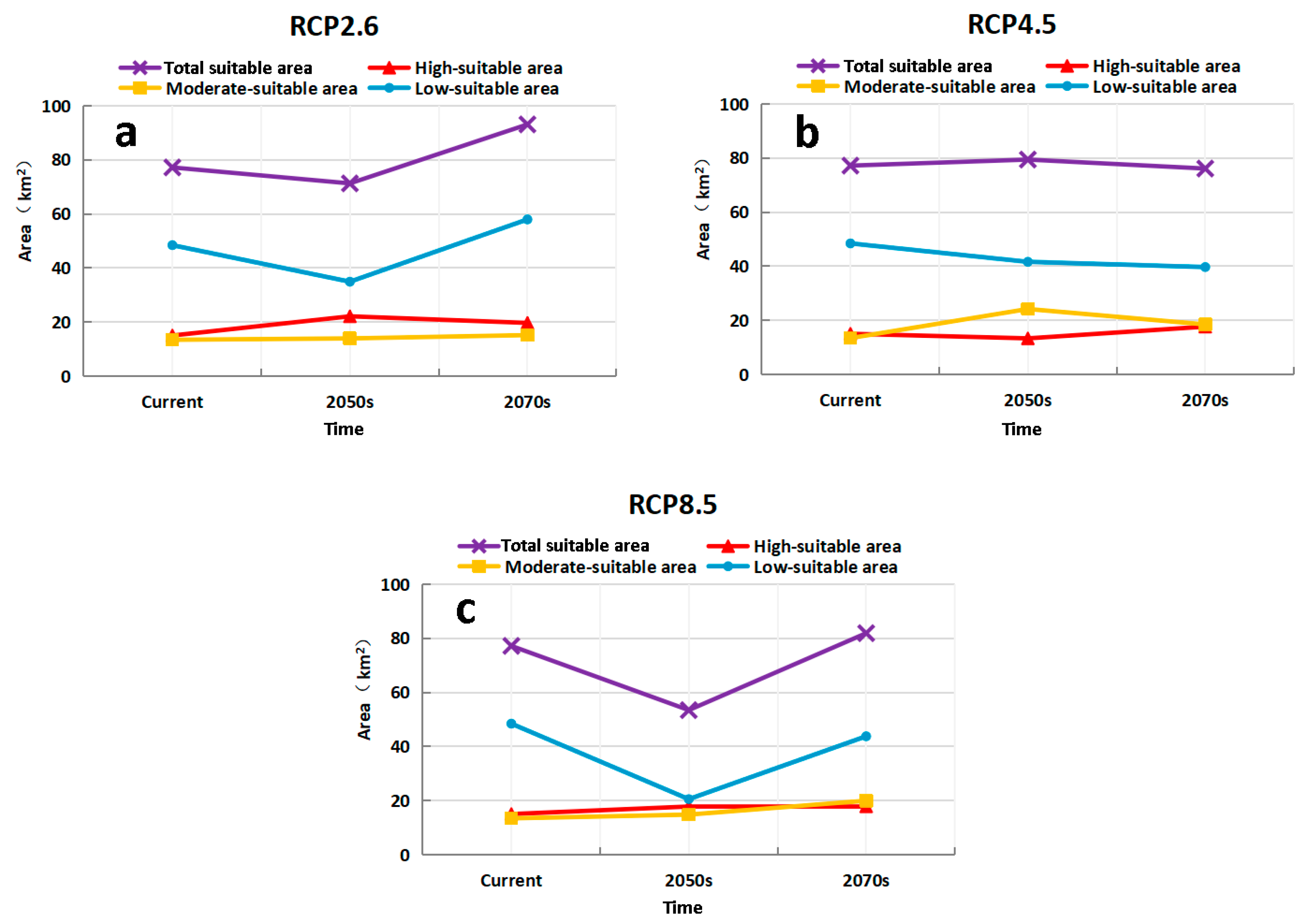
3.4. Predicting the Future Suitable Climatic Distribution of M. destructor in China
4. Discussion
4.1. The MaxEnt Model
4.2. Environmental Factors Affecting the Potential Distribution of M. destructor
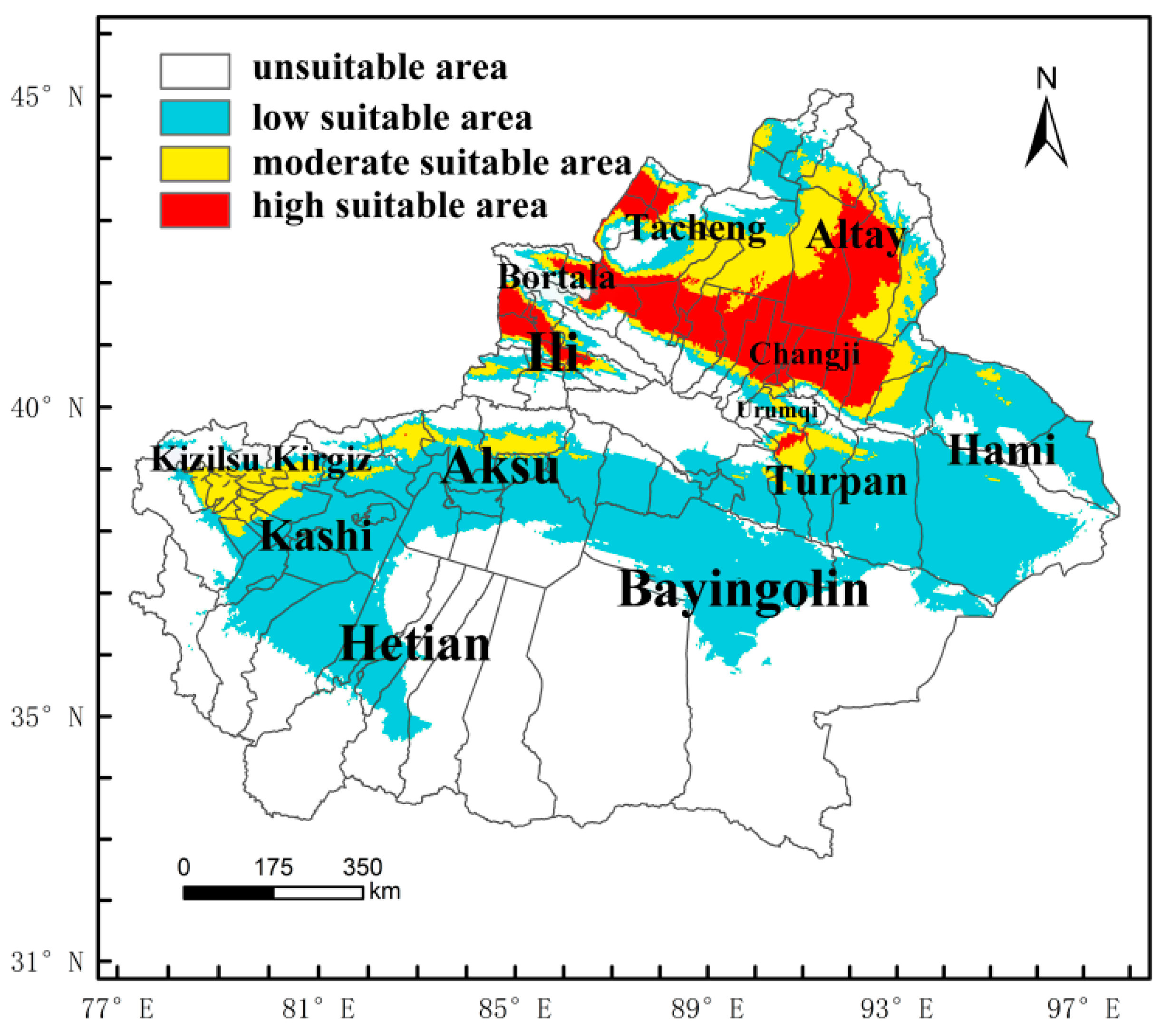
4.3. The Potential Distribution of M. destructor in China and in Xinjiang Only
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sentis, A.; Desneux, N. Editorial overview: Global change: Integrating ecological and evolutionary consequences across time and space. Curr. Opin. Insect Sci. 2019, 35, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Sattar, Q.; Maqbool, M.E.; Ehsan, R.; Akhtar, S. Review on climate change and its effect on wildlife and ecosystem. J. Environ. Biol. 2021, 6, 8–14. [Google Scholar] [CrossRef]
- Shrestha, S. Effects of climate change in agricultural insect pest. Acta Sci. Agric. 2019, 3, 74–80. [Google Scholar] [CrossRef]
- Commission of the European Communities. Communication from the Commission to the Council, the European Parliament, the European Economic and Social Committee and the Committee of the Regions: Winning the Battle against Global Climate Change. COM 35 Final; Commission of the European Communities: Brussels, Belgium, 2005.
- Bellard, C.; Jeschke, J.M.; Leroy, B.; Mace, G.M. Insights from modeling studies on how climate change affects invasive alien species geography. Ecol. Evol. 2018, 8, 5688–5700. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Ma, G.; Pincebourde, S. Survive a warming climate: Insect responses to extreme high temperatures. Annu. Rev. Entomol. 2021, 7, 163–184. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Buffo, E.; Larsson, S. A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Glob. Chang. Biol. 2006, 12, 662–671. [Google Scholar] [CrossRef]
- Monteith, J. Agricultural meteorology: Evolution and application. Agric. For. Meteorol. 2000, 103, 5–9. [Google Scholar] [CrossRef]
- Aidoo, O.F.; Souza, P.G.C.; da Silva, R.S.; Santana Júnior, P.A.; Picanço, M.C.; Kyerematen, R.; Sétamou, M.; Ekes, S.; Borgemeister, C. Climate-induced range shifts of invasive species (Diaphorina citri Kuwayama). Pest Manag. Sci. 2022, 78, 2534–2549. [Google Scholar] [CrossRef]
- Aidoo, O.F.; Souza, P.G.C.; da Silva, R.S.; Santana Júnior, P.A.S.; Picanço, M.C.; Osei-Owusu, J.; Sétamou, M.; Ekesi, S.; Borgemeister, C. A machine learning algorithm-based approach (MaxEnt) for predicting invasive potential of Trioza erytreae on a global scale. Ecol. Inf. 2022, 71, 101792. [Google Scholar] [CrossRef]
- Zhang, H.; Song, J.; Zhao, H.; Li, M.; Han, W. Predicting the distribution of the invasive species Leptocybe invasa: Combining MaxEnt and Geodetector models. Insects. 2021, 12, 92. [Google Scholar] [CrossRef]
- Tadesse, W.; El-Hanafi, S.; El-Fakhouri, K.; Imseg, I.; Rachdad, F.E.; El-Gataa, Z.; El-Bouhssini, M. Wheat breeding for Hessian fly resistance at ICARDA. Crop J. 2022, in press. [Google Scholar] [CrossRef]
- Harris, M.O.; Sandanayaka, M.; Griffin, A. Oviposition preferences of the Hessian fly and their consequences for the survival and reproductive potential of offspring. Ecol. Entomol. 2001, 26, 473–486. [Google Scholar] [CrossRef]
- Chen, M.S.; Liu, X.; Wang, H.; Bouhssini, M. Hessian fly (Diptera: Cecidomyiidae) interactions with barley, rice, and wheat seedlings. J. Econ. Entomol. 2009, 102, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Naber, N.; Bouhssini, M.; Labhilili, M.; Udupa, S.; Nachit, M.M.; Baum, M.; Lhaloui, S.; Benslimane, A.; Abbouyi, H. Genetic variation among populations of the Hessian fly, Mayetiola destructor (Diptera: Cecidomyiidae), in Morocco and Syria. Bull. Entomol. Res. 2000, 90, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, H.F. Gall Midges of Economic Importance. Vol. VII. Gall Midges of Cereal Crops; Crosby Lockwood & Son: London, UK, 1956; Volume 7, pp. 261–526. [Google Scholar]
- Wellso, S.G. Aestivation and phenology of the Hessian fly (Diptera: Cecidomyiidae) in Indiana. Environ. Entomol. 1991, 20, 795–801. [Google Scholar] [CrossRef]
- Schmid, R.B.; Allen, K.; Giles, K.L.; Brian, P.M. Hessian fly (Diptera: Cecidomyiidae) biology and management in wheat. J. Integr. Pest Manag. 2018, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Gagne, R.J.; Hatchett, J.H. Instars of the Hessian fly (Diptera: Cecidomyiidae). Ann. Entomol. Soc. Am. 1989, 82, 73–79. [Google Scholar] [CrossRef]
- Whitworth, R.J.; Sloderbeck, P.E.; Davis, H.; Cramer, G. Kansas crop pests: Hessian fly. Kans. State Univ. Agric. Exp. Stn. Coop. Ext. Serv. 2009, MF-2866, 1–3. [Google Scholar]
- Bergh, J.C.; Harris, M.O.; Rose, S. Temporal patterns of emergence and reproductive-behavior of the Hessian fly (Diptera: Cecidomyiidae). Ann. Entomol. Soc. Am. 1990, 83, 998–1004. [Google Scholar] [CrossRef]
- Withers, T.M.; Harris, M.O.; Madie, C. Dispersal of mated female Hessian flies (Diptera: Cecidomyiidae) in field arrays of host and nonhost plants. Environ. Entomol. 1997, 26, 1247–1257. [Google Scholar] [CrossRef]
- Foster, J.E.; Taylor, P.L. Thermal-unit requirements for development of the Hessian fly under controlled environments. Environ. Entomol. 1975, 4, 195–202. [Google Scholar] [CrossRef]
- Buntin, G.; Chapin, J.W. Biology of Hessian fly (Diptera: Cecidomyiidae) in the southeastern United States: Geographic variation and temperature-dependent phenology. J. Econ. Entomol. 1990, 83, 1015–1024. [Google Scholar] [CrossRef]
- McColloch, J.W. The Hessian fly in Kansas. Kans. Agric. Exp. Stn. Bull. 1923, 11, 57–58. [Google Scholar]
- Stuart, J.J.; Chen, M.S.; Shukle, R.; Harris, M.O. Gall midges (Hessian flies) as plant pathogens. Annu. Rev. Phytopathol. 2012, 50, 339–357. [Google Scholar] [CrossRef]
- Zhang, X.Z. The discovery and investigation of Mayetiola destructor in Xinjiang. J. Plant Protect. 1983, 10, 1–10. (In Chinese) [Google Scholar] [CrossRef]
- Dai, A.M.; Liang, Q.L.; Zhang, H.; Lu, P. Outbreak and the reasons of Hessian fly in Bortala Mongol Autonomous Prefecture, Xinjiang. Plant Quar. 2014, 28, 60–63. (In Chinese) [Google Scholar] [CrossRef]
- Hu, W.F.; Chen, L.L.; Yao, J.Q.; Ji, S.Y.; Sun, N. Analysis of the temporal and spatial evolution of temperature and precipitation in Xinjiang under the background of climate change. J. Fuyang Norm. Univ. Nat. Sci. 2020, 37, 90–95. [Google Scholar] [CrossRef]
- Franklin, J. Species distribution models in conservation biogeography: Developments and challenges. Divers. Distrib. 2013, 19, 1217–1223. [Google Scholar] [CrossRef]
- Li, Y.; Cao, W.; He, X.; Chen, W.; Xu, S. Prediction of suitable habitat for lycophytes and ferns in Northeast China: A case study on Athyrium Brevifrons. Chin. Geogr. Sci. 2019, 29, 1011–1023. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Zhuo, Z.; Wang, R.; Ye, M.; Pu, B. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Glob. Ecol. Conserv. 2019, 19, e00691. [Google Scholar] [CrossRef]
- Sarquis, J.A.; Cristaldi, M.A.; Arzamendia, V.; Bellini, G.; Giraudo, A.R. Species distribution models and empirical test: Comparing predictions with well-understood geographical distribution of Bothrops alternatus in Argentina. Ecol. Evol. 2018, 8, 10497–10509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.L.; Wang, M.T.; Luo, J.D.; Liu, Y.; Wu, S.Q.; Wen, G.; Li, Q. The analysis of climate suitability and regionalization of Actinidia deliciosa by using MaxEnt model in China. J. Yunnan Agric. Univ. Nat. Sci. 2019, 34, 522–531. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Ji, W.; Gao, G.; Wei, J. Potential global distribution of Daktulosphaira vitifoliae under climate change based on MaxEnt. Insects 2021, 12, 347. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhang, H.; Li, M.; Han, W.; Yin, Y.; Lei, J. Prediction of spatiotemporal invasive risk of the red import fire ant, Solenopsis invicta (Hymenoptera: Formicidae), in China. Insects 2021, 12, 874. [Google Scholar] [CrossRef] [PubMed]
- Pascoe, E.L.; Marcantonio, M.; Caminade, C.; Foley, J.E. Modeling potential habitat for Amblyomma tick species in California. Insects 2019, 10, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, D.L.; Wright, A.N.; Seifert, S.N.; Shaffer, H.B. Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern. Divers. Distrib. 2014, 20, 334–343. [Google Scholar] [CrossRef]
- Jia, X.; Wang, C.; Jin, H.; Zhao, Y.; Liu, L.J.; Chen, Q.H.; Li, B.Y.; Xiao, Y.; Yin, H. Assessing the suitable distribution area of Pinus koraiensis based on an optimized MaxEnt model. Chin. J. Ecol. 2019, 38, 7. [Google Scholar] [CrossRef]
- Zou, Y.; Ge, X.; Guo, S.; Zhou, Y.; Wang, T.; Zong, S. Impacts of climate change and host plant availability on the global distribution of Brontispa longissima (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2020, 76, 244–256. [Google Scholar] [CrossRef]
- Wu, W.; Li, Z.H.; Hang, X. The predictive research on the potential adaptable areas of Mayetiola destructor in China based on CLIMEX. Plant Quar. 2015, 29, 20–24. (In Chinese) [Google Scholar]
- Li, X.Y.; Xu, D.P.; Jin, Y.W.; Zhuo, Z.H.; Yang, H.J.; Hu, J.M.; Wang, R.L. Predicting the current and future distributions of Brontispa longissima (Coleoptera: Chrysomelidae) under climate change in China. Glob. Ecol. Conserv. 2021, 25, e01444. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013.
- Li, A.; Wang, J.; Wang, R.; Yang, H.; Yang, W.; Yang, C.; Jin, Z. MaxEnt modeling to predict current and future distributions of Batocera lineolata (Coleoptera: Cerambycidae) under climate change in China. Ecoscience 2019, 27, 23–31. [Google Scholar] [CrossRef]
- Zhu, G.P.; Qiao, H.J. Effect of the Maxent model’s complexity on the prediction of species potential distributions. Biodivers. Conserv. 2016, 24, 1189–1196. [Google Scholar] [CrossRef]
- Liu, X.Y.; Zhao, C.Y.; Li, F.; Zhu, J.F.; Gao, K.X.; Hu, Y.P. Prediction of potential geographical distribution of Solenopsis invicta Buren in China based on MaxEnt. Plant Quar. 2019, 33, 70–76. (In Chinese) [Google Scholar] [CrossRef]
- Wang, Y.S.; Xie, B.Y.; Wan, F.H.; Xiao, Q.M.; Dai, L.Y. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models. Biodiv. Sci. 2007, 4, 365–372. [Google Scholar]
- Zhang, X.; Li, X.; Feng, Y.; Liu, Z. The use of ROC and AUC in the validation of objective image fusion evaluation metrics. Signal Process. 2015, 115, 38–48. [Google Scholar] [CrossRef]
- Waage, J.K.; Reaser, J.K. A global strategy to defeat invasive species. Science. 2001, 292, 1486. [Google Scholar] [CrossRef]
- Zeng, Y.; Low, B.W.; Yeo, D.C.J. Novel methods to select environmental variables in MaxEnt: A case study using invasive crayfish. Ecol. Model. 2016, 341, 5–13. [Google Scholar] [CrossRef]
- Babasaheb, B.F.; Shashank, P.R.; Suroshe, S.S.; Chandrashekar, K.; Meshram, N.M.; Timmanna, H.N. Invasion risk of the South American tomato pinworm Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in India: Predictions based on MaxEnt ecological niche modelling. Int. J. Trop. Insect Sci. 2020, 40, 561–571. [Google Scholar] [CrossRef]
- Soberon, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Soberon, J.M. Niche and area of distribution modeling: A population ecology perspective. Ecography 2010, 33, 159–167. [Google Scholar] [CrossRef]
- Hamilton, E.W. Hessian fly larval strain responses to simulated weather conditions in the greenhouse and laboratory. J. Econ. Entomol. 1966, 59, 535–538. [Google Scholar] [CrossRef]
- Anonymous. The Hessian Fly; Xinjiang People’s Press: Urumqi, China, 1986; pp. 34–46. [Google Scholar]
- Morgan, G.; Sansone, C.; Knutson, A. Hessian fly in Texas wheat. Tex. AM Agrilife Ext. Serv. Ext. Pub. 2005, E-350, 1–7. Available online: https://hdl.handle.net/1969.1/87300 (accessed on 15 October 2022).
- Prestidge, R.A. Population biology and parasitism of Hessian fly (Mayetiola destructor) (Diptera: Cecidomyiidae) on Bromus willdenowii in New Zealand. N. Z. J. Agric. Res. 1992, 35, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.M.; Liu, J. Analysis of spatial-temporal dynamic change of wheat planting structure of China. Chin. Agric. Sci. Bull. 2019, 35, 12–23. [Google Scholar]
- Harris, M.O.; Rose, S. Factors influencing the onset of egglaying in a cecidomyiid fly. Physiol. Entomol. 1991, 16, 183–190. [Google Scholar] [CrossRef]
- McColloch, J.W. Wind as a factor in the dispersion of the Hessian fly. Econ. Entomol. 1917, 10, 162–168. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Factors |
|---|---|
| Bio1 | Annual mean temperature (°C) |
| Bio2 | Mean diurnal range (°C) (Monthly mean [max temp − min temp]) |
| Bio3 | Isothermality ([BIO2/BIO7] [×100]) |
| Bio4 | Temperature seasonality (standard deviation ×100) (°C) |
| Bio5 | Maximum temperature of the warmest month (°C) |
| Bio6 | Minimum temperature of the coldest month (°C) |
| Bio7 | Temperature annual range (BIO5-BIO6) (°C) |
| Bio8 | Mean temperature of wettest quarter (°C) |
| Bio9 | Mean temperature of driest quarter (°C) |
| Bio10 | Mean temperature of warmest quarter (°C) |
| Bio11 | Mean temperature of coldest quarter (°C) |
| Bio12 | Annual precipitation (mm) |
| Bio13 | Precipitation of the wettest month (mm) |
| Bio14 | Precipitation of the driest month (mm) |
| Bio15 | Precipitation seasonality (coefficient of variation) |
| Bio16 | Precipitation of the wettest quarter (mm) |
| Bio17 | Precipitation of the driest quarter (mm) |
| Bio18 | Precipitation of the warmest quarter (mm) |
| Bio19 | Precipitation of the coldest quarter (mm) |
| Tmin | Average monthly minimum temperature (°C) |
| Tmax | Average monthly maximum temperature (°C) |
| Tmean | Average monthly mean temperature (°C) |
| Prec | Average monthly precipitation (mm) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Guo, J.-L.; Guo, Y.; Guo, Z.; Lu, P.; Hu, X.-S.; Zhang, H.; Liu, T.-X. Prediction of the Current and Future Distributions of the Hessian Fly, Mayetiola destructor (Say), under Climatic Change in China. Insects 2022, 13, 1052. https://doi.org/10.3390/insects13111052
Ma Q, Guo J-L, Guo Y, Guo Z, Lu P, Hu X-S, Zhang H, Liu T-X. Prediction of the Current and Future Distributions of the Hessian Fly, Mayetiola destructor (Say), under Climatic Change in China. Insects. 2022; 13(11):1052. https://doi.org/10.3390/insects13111052
Chicago/Turabian StyleMa, Qi, Jin-Long Guo, Yue Guo, Zhi Guo, Ping Lu, Xiang-Shun Hu, Hao Zhang, and Tong-Xian Liu. 2022. "Prediction of the Current and Future Distributions of the Hessian Fly, Mayetiola destructor (Say), under Climatic Change in China" Insects 13, no. 11: 1052. https://doi.org/10.3390/insects13111052
APA StyleMa, Q., Guo, J.-L., Guo, Y., Guo, Z., Lu, P., Hu, X.-S., Zhang, H., & Liu, T.-X. (2022). Prediction of the Current and Future Distributions of the Hessian Fly, Mayetiola destructor (Say), under Climatic Change in China. Insects, 13(11), 1052. https://doi.org/10.3390/insects13111052