1. Introduction
The planthopper
Pentastiridius leporinus belongs to the family Cixiidae in the infraorder Fulgoromorpha. Cixiid species are small hemipterans that feed as adult stages on aboveground plant material, whereas their nymph stages feed below the ground on root sap. Many cixiid species are economically important because they transmit bacterial pathogens to plants.
P. leporinus is a vector of the bacterial yellowing disease Syndrome des Basses Richesses (SBR) in sugar beet (
Beta vulgaris), and large populations of this insect are found in sugar beet crops [
1]. The symptoms of SBR include yellowing of older leaves, lancet-shaped leaf deformations, and necrosis of the vascular tissue, which significantly reduces the sugar content of root tissue and results in lower yields. SBR was first identified in France [
2,
3] but is now spreading rapidly in southern and eastern Germany, threatening the profitability of the sugar beet industry [
4].
In central Europe,
P. leporinus produces one generation per year. After mating at the end of June, beginning of July, the females deposit their egg mass in the soil and the nymphs continue to develop in the topsoil. Adult females and nymphs produce filamentous wax structures that protect them and the eggs from predation [
5]. Sugar beet is harvested in fall, and winter wheat is sown shortly afterwards, providing the
P. leporinus nymphs with an overwinter/spring food source until they emerge as adults and seek a new sugar beet crop [
6].
SBR is associated with two different bacterial pathogens,
Candidatus Arsenophonus phytopathogenicus and
Candidatus Phytoplasma solani, both of which are transmitted by insect vectors and limited to the phloem [
7,
8].
Candidatus Arsenophonus phytopathogenicus (phylum Gammaproteobacteria), hereafter abbreviated to SBRars, is an intracellular bacterium that is transmitted vertically from
P. leporinus females to their offspring with an efficiency of ~30%, and with a lower efficiency by males due to the presence of fewer bacteria in the male salivary gland [
9].
Candidatus Phytoplasma solani, hereafter abbreviated to SBRps, is commonly known as stolbur phytoplasma. Phytoplasmas are bacterial pathogens (family
Acholeplasmataceae) that lack a cell wall and colonize the phloem sieve tubes of plants, resulting in various economically important crop diseases [
10]. In Serbia, stolbur phytoplasma is also associated with a different disease of sugar beet, known as rubbery tap root disease [
11].
There are currently no effective measures against either of the SBR pathogens or the insect vector. Effective vector control requires a comprehensive knowledge of the distribution and behavior of P. leporinus, but the lack of mass breeding protocols makes it difficult to assess life cycle and life history traits in order to model the spread of the disease and test biological and environmentally safe alternative control measures in the laboratory and greenhouse. We therefore set out to develop the first mass rearing methods for this insect species. Furthermore, we gathered information on the flight behavior of the planthopper in different regions to determine the most suitable application period for any new control measures and investigated the subterranean movement of nymphs as a function of soil temperature in order to design strategies that interrupt the P. leporinus life cycle in the soil. Finally, we screened for both SBR pathogens at four sites in Rhineland-Palatinate and Hesse to monitor the spread of the disease in Germany. A better understanding of P. leporinus biology and the SBR pathogens will facilitate the development of environmentally friendly control strategies and assist with pest management decisions to control the spread of SBR.
2. Materials and Methods
2.1. Field Collection of Adults
We collected P. leporinus adults from commercial sugar beet fields during July 2020 in Biblis (49°41′52.6″ N 8°27′12.8″ E) and during June and July 2021 in Bickenbach (49°45′46.0″ N 8°33′36.2″ E). Both fields are located in the Rhine Valley close to the border of the Odenwald (89 m above sea level) in Hesse. Adults were collected using a sweep net and were immediately transferred to the greenhouse for rearing.
2.2. Rearing
Field-collected P. leporinus adults were placed directly on 8-week-old sugar beet plants (KWS Annarosa) growing in Göttinger 3-L pots (Lamprecht-Verpackungen, Göttingen, Germany) filled with Frühstorfer potting soil LD 80 (Heinrichs, Ingelheim, Germany) and covered with expanded clay (Floragard, Oldenburg, Germany). The insects were kept in groups of ~400 in a Cavea PopUp size M net cage (Howitec, Bolsward, Netherlands). The sugar beet plant in the cage was changed every 7 days. Plants were watered moderately twice weekly. The temperature in the greenhouse chamber was set to 22/16 °C day/night with a 16 h photoperiod.
In 2020, egg clusters were left inside the pot, allowing the nymphs to develop on the sugar beet roots. When the plant wilted (due to SBR infection), the nymphs were transferred to a new sugar beet plant. In 2021, sugar beet plants from the adult cages were carefully screened for eggs, which were transferred to a climate chamber (continuous 22 °C and 60% relative humidity, 16 h photoperiod). Mass rearing was carried out in GODMORGEN plastic boxes (IKEA Deutschland, München, Germany) filled with the granular soil additive GeoHumus (GeoHumus, Frankfurt/Main, Germany) to maintain humidity. Young sugar beet plants were placed in the boxes as a food source and were replaced weekly.
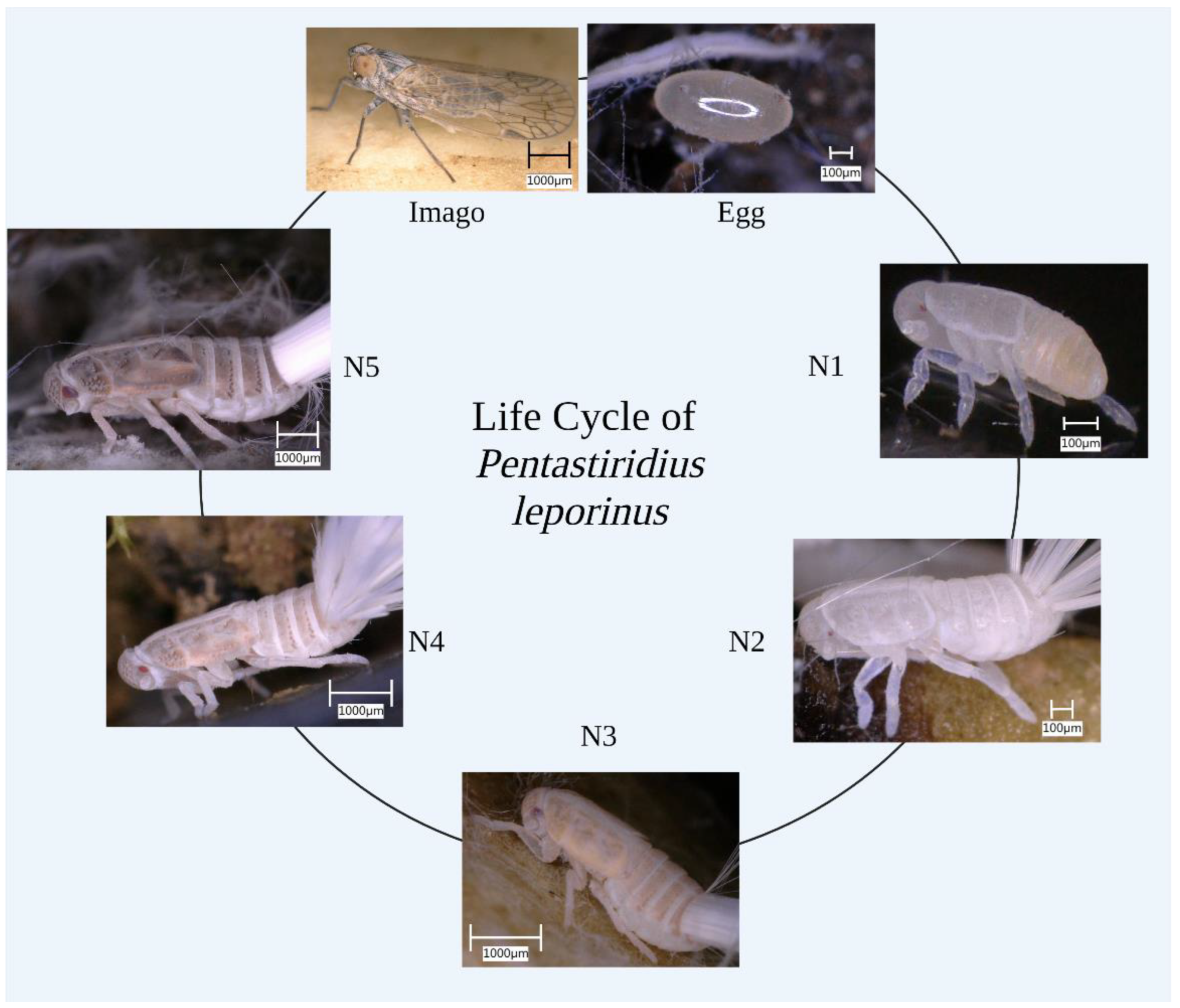
2.3. Life Cycle Data
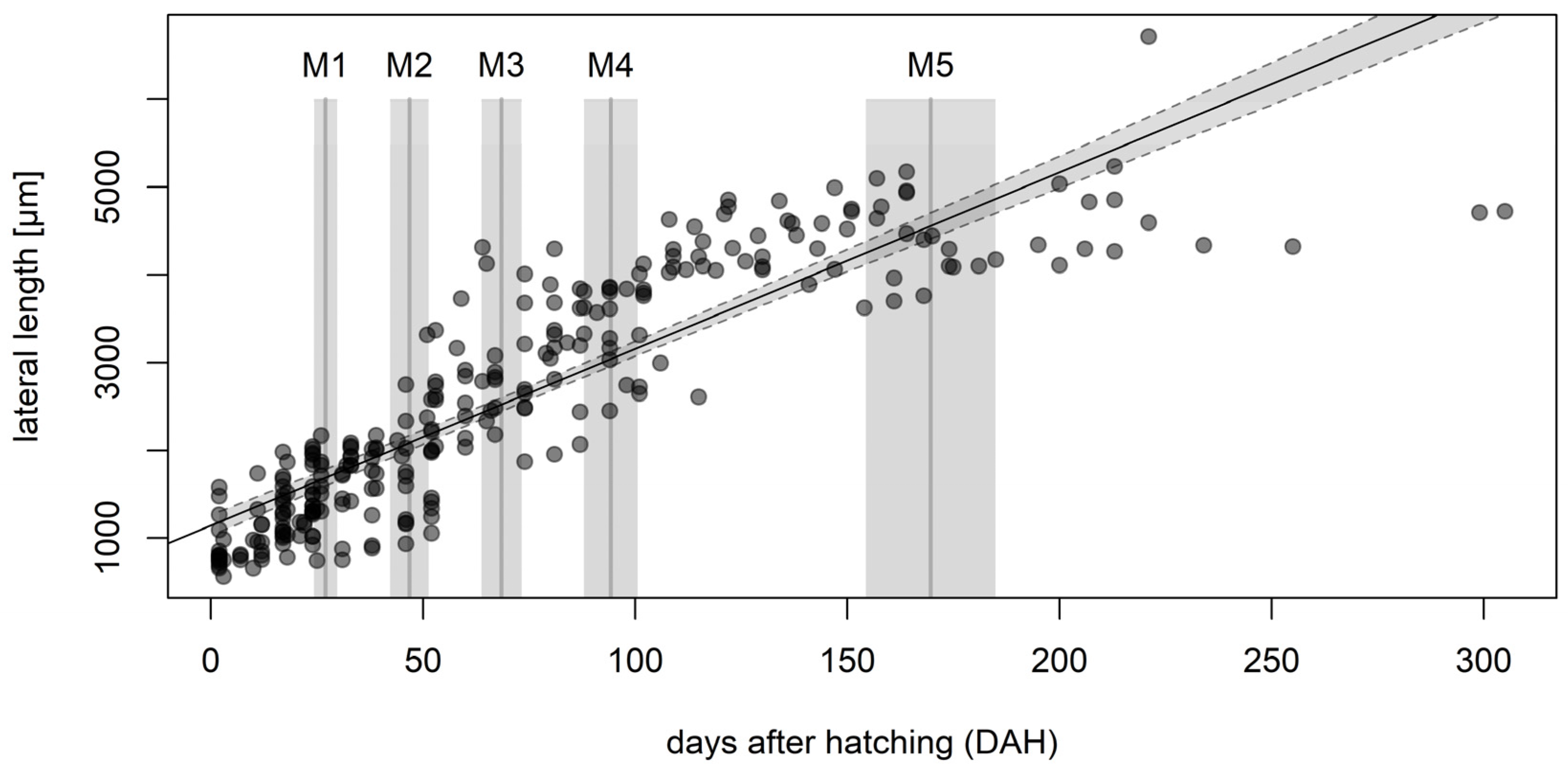
Solitary nymphs were maintained in 60 mM Petri dishes with GeoHumus substrate and 1 × 1 × 1 cm cubes of young sugar beet roots as nutrition. The developmental stages were documented and measured using a VHX-5000 digital microscope (Keyence Deutschland, Neu-Isenburg, Germany) on a weekly basis for 34 weeks. The body size of individual nymphs was measured along the lateral axis [
12] and the pigmentation of the nymphs was used for further characterization.
2.4. Host Plant Choice Experiments
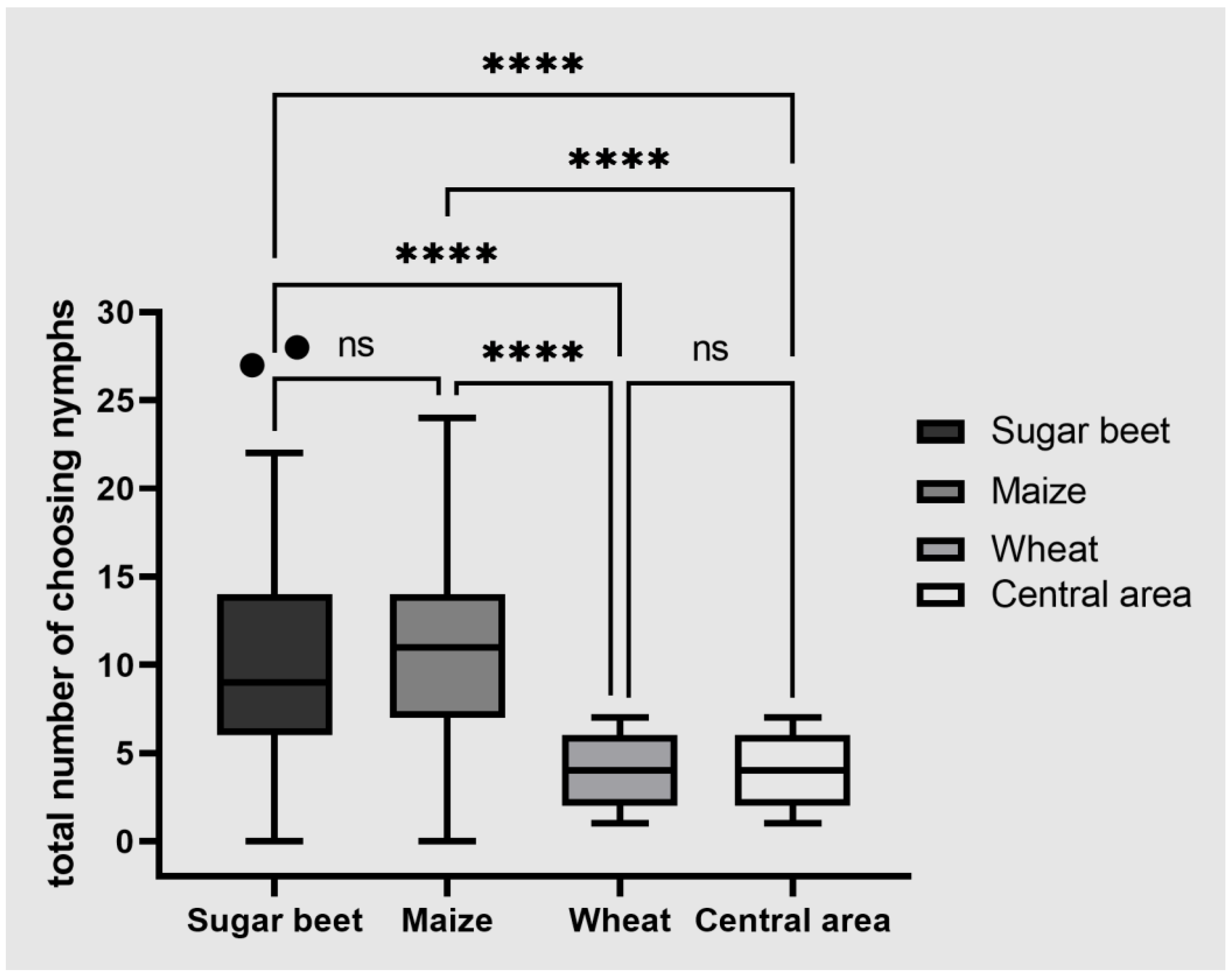
Sugar beet, wheat and maize were presented to P. leporinus nymphs (the first progeny of field-collected adults) to determine their host plant preference. The host choice experiments were carried out using five replicate cohorts of 30 nymphs. The number of nymphs on each host plant was assessed after 1, 3, 6, 12, 24 and 48 h. The experimental setup consisted of four areas; a central area created by a plastic container (8 × 10.5 × 9 cm) from which three transparent plastic pipes (1.7 cm diameter) led to three separate plastic containers holding the different plants. The nymphs were released to the central area. The stalks of the plants were wrapped with 2 cm foam strips and fixed in the perforated lids of the containers, to make the roots available for the nymphs.
The host plants were grown in a hydroponic system. Maize and wheat seeds were transferred to a sandwich culture [
13] and grown in 1 mM CaSO
4, whereas sugar beet seed was grown in 1 mM CaSO
4 containing 10 µM boric acid. Before germination, all cultures were coated with the seed treatment Maxim XL (Syngenta, Basel, Switzerland) containing the fungicides mefenoxam and fludioxonil. Sandwich cultures were grown under controlled conditions from day 4 (26/18 °C day/night, 16-h photoperiod). On day 8, the seedlings were transferred to 2.5-L containers filled with semi-concentrated nutrient solution containing the following nutrient salts: calcium nitrate tetrahydrate, potassium sulfate, magnesium sulfate heptahydrate, calcium chloride dihydrate, monobasic potassium phosphate, ferric EDTA, boric acid, manganese(II) sulfate monohydrate, zinc sulfate solution, copper(II) sulfate pentahydrate, ammonium heptamolybdate tetrahydrate and nickel(II) sulfate hexahydrate [
14]. The containers were maintained under controlled conditions for a further 4 days (21/16 °C day/night, 16 h photoperiod). The stalks of the seedlings were then wrapped with 2 cm foam strips and fixed in the perforated lids of the containers, into which aeration tubes were inserted. An Osaga LK 60-piston compressor (Osaga, Buchholz, Germany) was used to circulate air in the nutrient solutions. On day 12, the plants were refreshed with fully-concentrated nutrient solution, which was changed weekly. The host choice experiments were initiated when the plants reached BBCH stage 15 (sugar beet and maize) or 31 (wheat).
2.5. Abundance of Adults in the Field
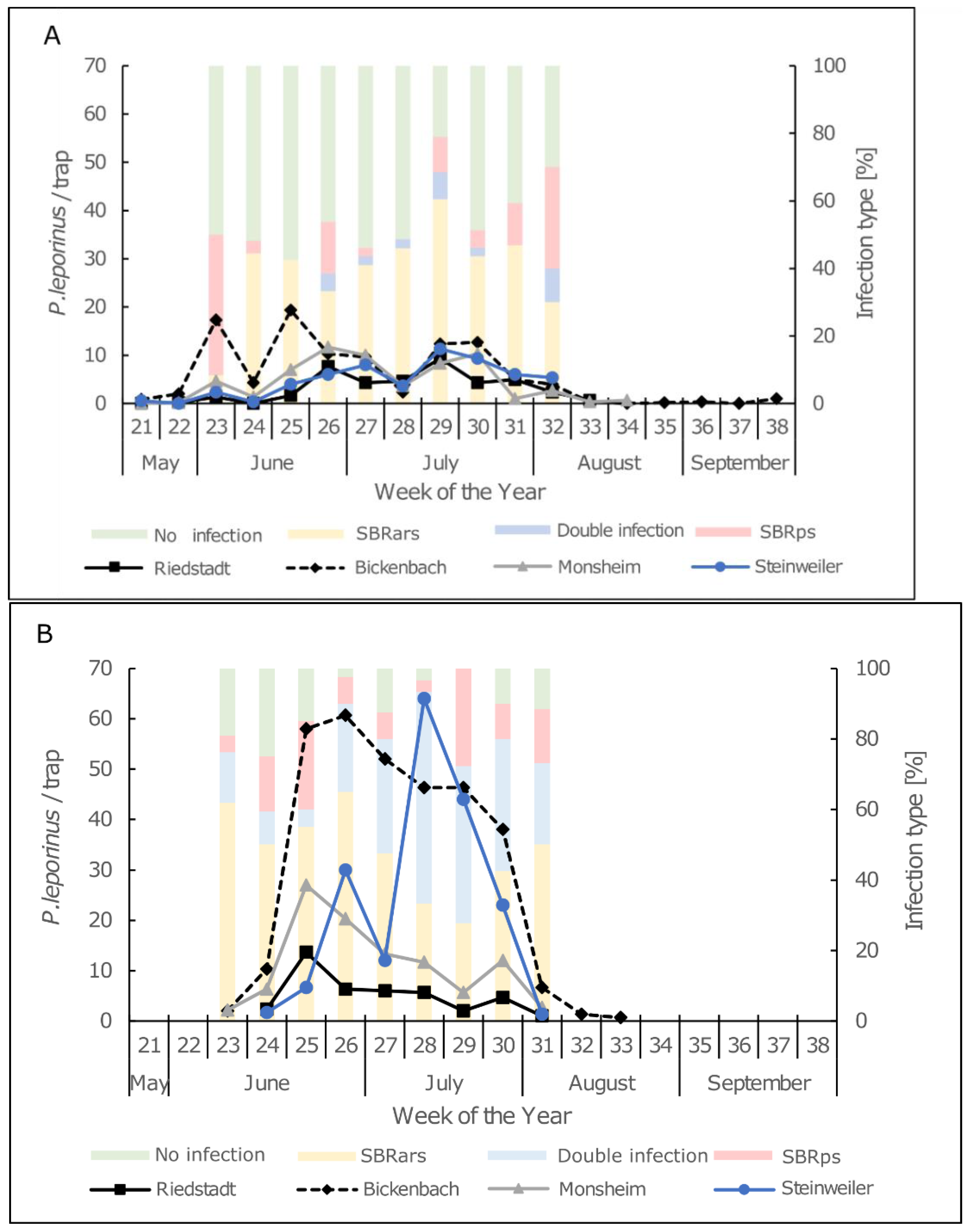
To measure the abundance of P. leporinus, 10 × 25 cm yellow sticky traps (Aeroxon, Waiblingen, Germany) were set up at four locations in Rhineland-Palatinate and south Hesse in the summer of 2020 and 2021. The sites were commercial sugar beet fields in Riedstadt (2020, 49°51′20.34″ N 8°23′47.67″ E; 2021, 49°52′18.2″ N 8°24′53.1″ E), Bickenbach (2020, 49°46′04.4″ N 8°33′30.9″ E; 2021, 49°46′03.2″ N 8°33′33.0″ E), Monsheim (2020, 49°37′59.2″ N 8°14′02.0″ E; 2021, 49°38′15.1″ N 8°14′42.9″ E) and Steinweiler (2020, 49°07′07.0″ N 8°09′03.6″ E; 2021, 49°06′29.8″ N 8°09′29.6″ E). Three traps were set up per site at a distance of 10, 30 and 50 m from the edge of the field on a pole at a height of 10 cm above the sugar beet vegetation. The panels were set up in the first week of May and the sampling was finished after two continuous weeks without catches. The sticky traps were replaced weekly and the number of adult P. leporinus on each trap was determined.
2.6. Subterranean Nymph Movement
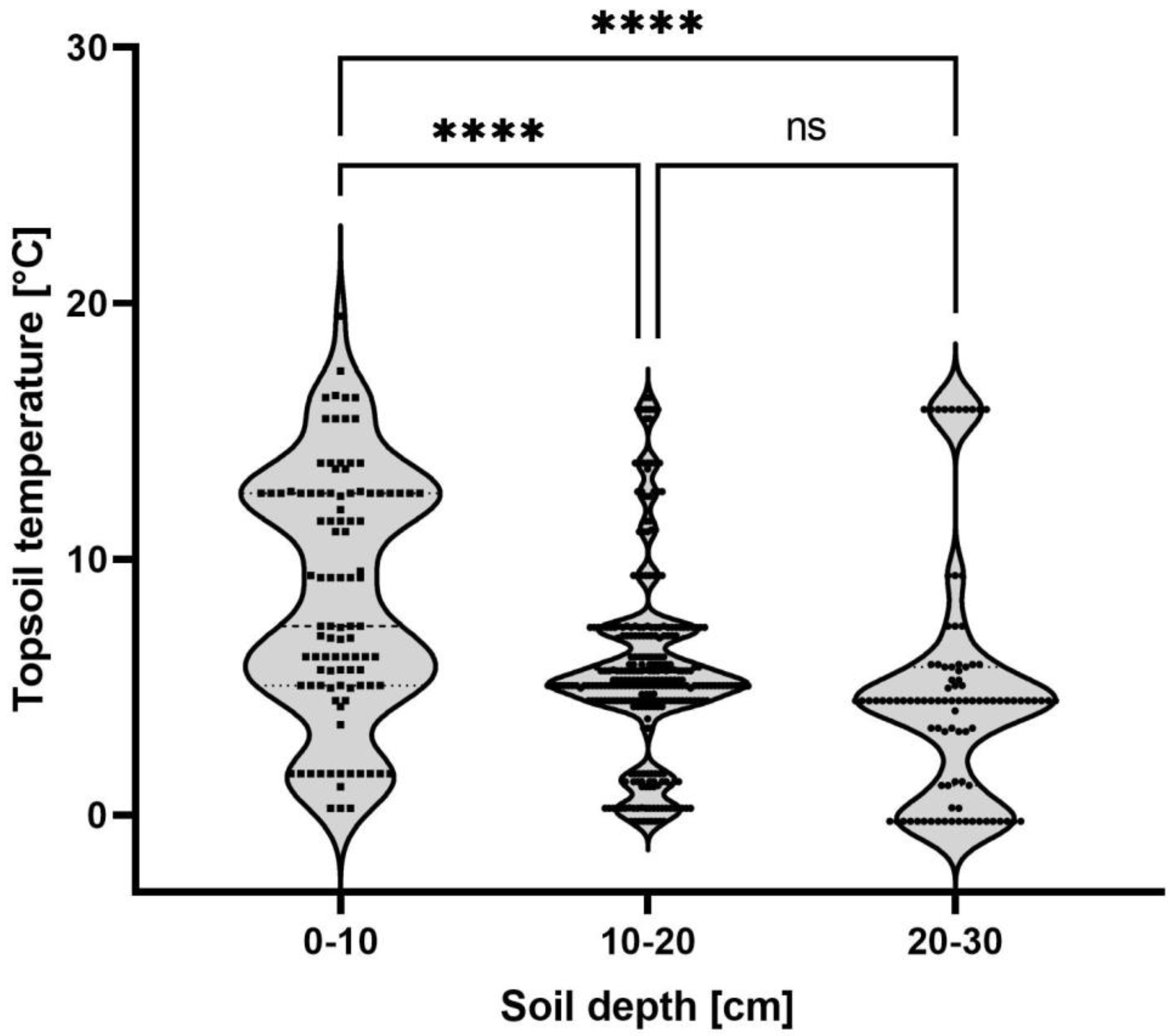
From mid-November 2020 until June 2021, we excavated soil from winter wheat fields following a heavily SBR-infested sugar beet crop (2020, 49°37′06.4″ N 8°07′45.5″ E). After the adult planthoppers had left the field in June 2021, the study was continued in a nearby sugar beet field (2021, 49°37′51.2″ N 8°07′00.3″ E) until March 2022, where peas were sown in spring after the sugar beet harvest. Once a week, six soil samples (25 × 25 × 25 cm) were taken at marked points to determine the whereabouts of nymphs during the course of the year. Each soil sample was sliced in 5 cm increments and the number of P. leporinus nymphs per slice was counted. After each survey, the sampling points were moved 1 m in different directions on six sampling lines. The study was conducted for 66 weeks in total. The soil temperature was recorded using a Tinytag Plus 2 TPG-4017 (Gemini Data Loggers, Chichester, UK) at 5–10 cm depth.
2.7. Detection of SBRars and SBRps by qRT-PCR
Trapped P. leporinus specimens were tested for infection with SBRars and SBRps in 2020 and 2021 by examining 343 and 296 planthoppers, respectively. In the flight monitoring, up to 10 planthoppers per week collected from four sites were transferred to Eppendorf tubes containing 70% ethanol, and DNA was extracted using the Blood&Tissue Kit (Qiagen, Hilden, Germany). The detection of SBRars and SBRps was determined by qRT-PCR (Applied Biosystems, Foster City, CA, USA). For SBRars, we used primers SBRars_qRTforKL437 (5′-CAC AAG CGG TGG AGC ATG TG-3′) and SBRars_qRTrevKL438 (5′-ACT CCC ATG GTG TGA CGG-3′) with the FAM/TAMRA-labeled probe SBRars_qRTprobeKL457 (5′-FAM-TCT CAA TTA CTC ATA GTG AGA T-TamRa-3′). For SBRps, we used primers SBRps_qRTforKL464 (5′-TGG AGG TTA TCA GAA GCA CAG-3′) and SBRps_qRTrevKL465 (5′-TGC TAA AGT CCC CAA CTT AAT G-3′) with the FAM/TAMRA-labeled probe SBRps_qRTprobeKL466 (5′-FAM-ATG TTG GGT TAA GTC CCG CAA CGA GCG CAA CC-TamRa-3′). Targets were amplified using the Luna Universal Probe qPCR master mix kit (New England Biolabs, Ipswich, MA, USA). The total DNA amount of the samples was adjusted to 10 ng/µL. A plasmid carrying the SBRars and SBRps target was used to prepare a standard curve including 10-fold dilution series. The samples were heated to 55 °C for 10 min, then 95 °C for 1 min, followed by 40 cycles of 95 °C for 10 s and 60 °C for 30 s in the amplification and fluorescence detection steps, respectively. The bacterial load was calculated against the standard curve using ABI 7500 System SDS Software (Applied Biosystems). In brief, we considered a threshold of 10 copies for SBRars and a threshold of 3 copies for SBRps, or for both cases a CT value higher than 35 as negative results.
2.8. Statistical Analysis
For growth rate analysis, a linear model was fitted to the lateral length data plotted against the number of days after hatching. Molting events were determined by observing the shed cuticle remnants, called exuviae, and were assigned to one of five developmental transitions based on the characteristic physical attributes of the nymphs. Host plant choice data were analyzed in GraphPad Prism v9.1.2 for Windows (GraphPad Software, San Diego, CA, USA) using Friedman’s test followed by Dunn’s multiple comparisons test. Subterranean nymph movement data were assessed by one-way analysis of variance (ANOVA) followed by Bonferroni’s multiple comparisons test, also in GraphPad Prism. Adult abundance data were analyzed in R v4.1.2 (R Core Team, Vienna, Austria, 2021) using a Mann–Whitney U-test and a Kruskal–Wallis test for data with non-normal distribution.
4. Discussion
The development of P. leporinus mass rearing protocols not only facilitated the life cycle studies described herein but will also allow the analysis of trait parameters that can be used to predict the future spreading and behavior of this pest. Furthermore, the mass rearing of P. leporinus is necessary to establish bioassays for the development of biological and environmentally friendly control methods. We developed a P. leporinus mass rearing method that was satisfactory in two different settings: a climate-controlled cabinet and greenhouse. Rearing in the climate-controlled cabinet required more effort than the greenhouse, but made it easier to collect adults and nymphs, and to trace them through their developmental stages. A key factor underpinning the success of rearing was the moderate and stringent watering regime and the use of GeoHumus or expanded clay to provide hiding places for oviposition and molting. The absence of an obligate diapause during the development of P. leporinus was in line with the field studies, where nymphs at different soil depths remained active even at low temperatures during winter.
The results of the host plant preference tests were important for several reasons. First, they showed that sugar beet is an attractive host for nymphs as well as adults, so there is no need to provide different host plant material during mass rearing. Second, they confirmed that nymphs are polyphagous, and indeed show no preference when offered a choice between sugar beet and maize. Winter wheat is the most common following crop after sugar beets, and it has been hypothesized that winter wheat might be a suitable host plant for
P. leporinus nymphs. However, our host plant preference test did not confirm a high attractiveness of winter wheat. One reason for the lack of attractiveness of wheat could be the amount and composition of volatiles released by the single plants. For further evaluation of the attractiveness for
P. leporinus nymphs solid-phase micro-extraction-gas chromatography-mass spectrometry could be performed [
15].
The initial host plant of
P. leporinus was the common reed
Phragmites australis, but the circumstances surrounding the switch from reed to sugar beet are unclear [
16]. It would be interesting to check for
P. leporinus strains that are still endemic in aquatic or semi-aquatic
P. australis and to compare them with strains that have adapted to sugar beet. Planthoppers have colonized subterranean habitats, as exemplified by the cavern-dwelling genus
Oliarus, and might serve as a model for rapid subterranean speciation [
17,
18].
Nymph nutrition and movement are important factors to consider when evaluating pest control strategies based on limiting the food supply of insect pests or the use of agronomic practices that disrupt their life cycle [
19]. We found that the nymphs are not restricted to sugar beet, only showing a weak preference for sugar beet over winter wheat and no preference between sugar beet and maize. The suggestion to switch from winter wheat to maize as a post-beet crop is therefore unlikely to inhibit
P. leporinus development. In terms of movement, it was shown that
P. leporinus nymphs develop underground and move through different soil layers based on sampling carried out in September, before sugar beet harvesting, and in May, just before adult emergence [
1]. Our results from weekly sampling show that nymphs migrate to the topsoil (0–10 cm) when the temperature reaches an average of 8.6 °C, providing an opportunity to use ploughing as a means to disrupt the life cycle as part of an integrated pest management program. Sugar beet is harvested between September and November, when the soil temperature in our study areas falls from an average of 13.7 °C to 6.6 °C, so ploughing soon after harvest would seem a feasible strategy to interfere with
P. leporinus movement. However, we also found that nymphs remained active even when the soil temperature dropped to 0 °C, similar to other fulgoromorph species such as
Hyalesthes obsoletus larvae found on lavender [
20].
We have provided the first experimental observations of
P. leporinus movement patterns in south Hesse and Rhineland-Palatinate. SBR was first detected in sugar beet roots in Heilbronn, Baden-Württemberg, in 2010 [
20], and the Palatinate has been infested with SBR since 2018 [
21]. No SBR infection was detected in the Riedstadt district of Groß-Gerau during yellowing disease monitoring in 2018 and 2019 [
4]. Infected sugar beet roots were first detected in the district of Groß-Gerau (south Hesse) in 2020 [
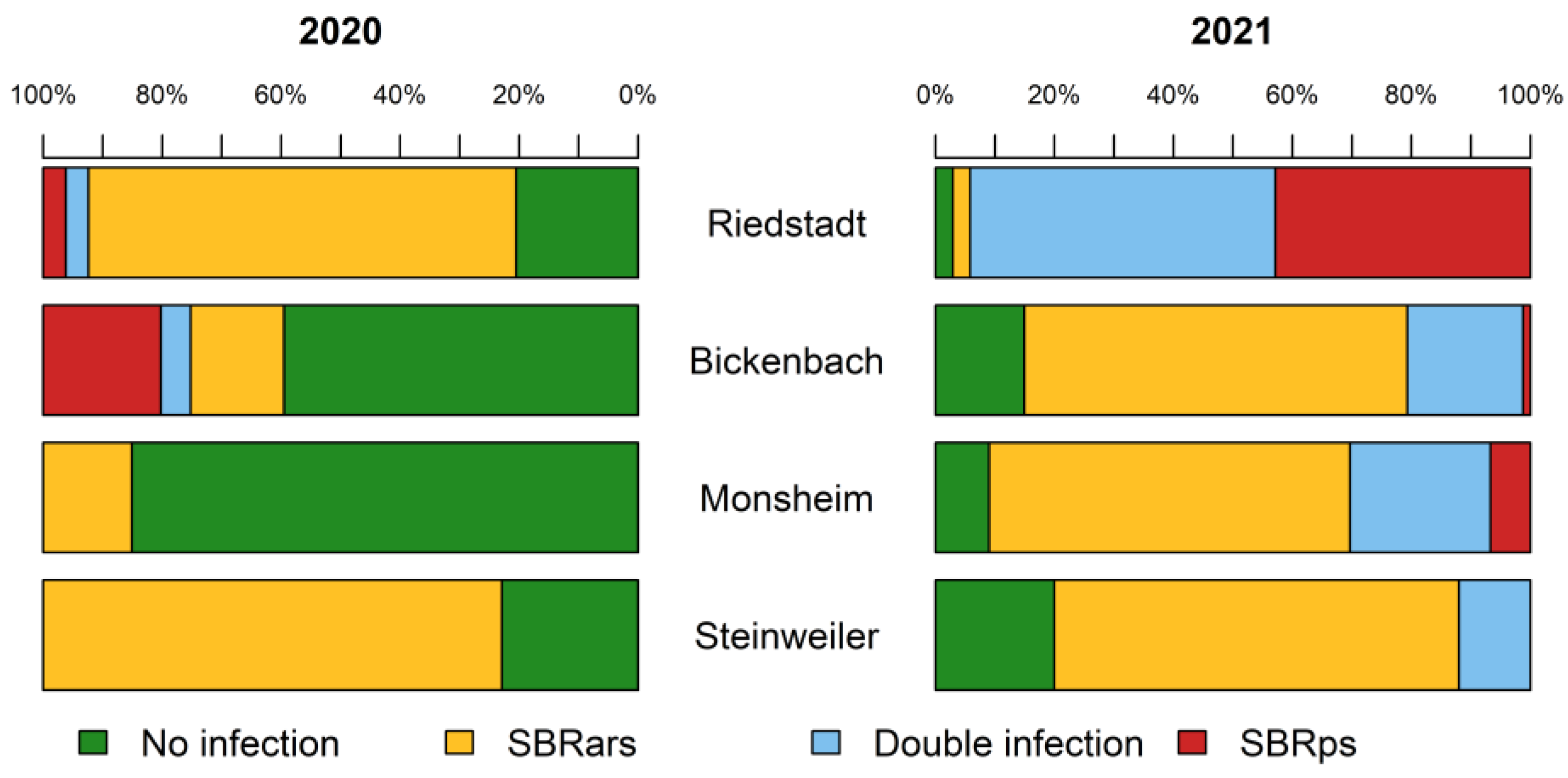
4]. The significant increase in the
P. leporinus population in south Hesse and the Palatinate between 2020 and 2021 may indicate the recent colonization of sugar beet crops in this area. Our field studies revealed that stolbur phytoplasma infection increased significantly between 2020 and 2021. To our knowledge, this is the first time a phytoplasma infection has exceeded 15% of a
P. leporinus population, which is alarming due to the key role of phytoplasma in SBR-induced damage to sugar beet, especially in south Hesse. Further studies are needed to understand the interactions between these pathogens and their host plant and insect vector. Including phytoplasma in future monitoring activity could help to predict the mass movement of
P. leporinus and thus prevent the spread of SBR by allowing the deployment of appropriate countermeasures. Another possibility is that the habitat is already infested the stolbur phytoplasma.
Candidatus Phytoplasma solani is also transmitted by
H. obsoletus in potato [
22]. In 2021, stolbur phytoplasma caused tremendous damage in south Hesse and the Palatinate by reducing the marketability of potato tubers (Michael Lenz, Regierungspräsidium Hessen, personal communication). The cross-infestation of potato and sugar beet by vector insects has yet to be investigated.
The movement pattern we observed was similar to that reported in Baden-Württemberg [
21] and Burgundy/Franche-Comté [
1]. The emergence of adults is subject to annual fluctuations. The main flight period is calendar weeks 23–31 with a peak during week 25. The long flight period of
P. leporinus is the main challenge when seeking control strategies.
P. leporinus females carrying SBRars were shown to remain infectious over their entire adult lifespan of 30 days [
9], whereas SBRps transmission assays by the same group were carried out using
H. obsoletus because no infected
P. leporinus could be found at the time of their study [
23]. The two pathogens affected sugar beet plants in different ways, with the sugar content (Brix) of the roots and the mean tap root biomass as a proportion of total plant biomass affected more severely by SBRps than SBRars [
24]. No data are yet available showing the effect of dual infections on sugar beet yields, but this information would facilitate the development of suitable SBR control measures.


 and
and 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}