


Comparison of Two Different Morphological Methods to Study the Pronotum of Cimicidae: Bed Bugs, Bat Bugs, and Swallow Bugs
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collections
2.2. Specimen Mounting, Photography, and Morphological Identification
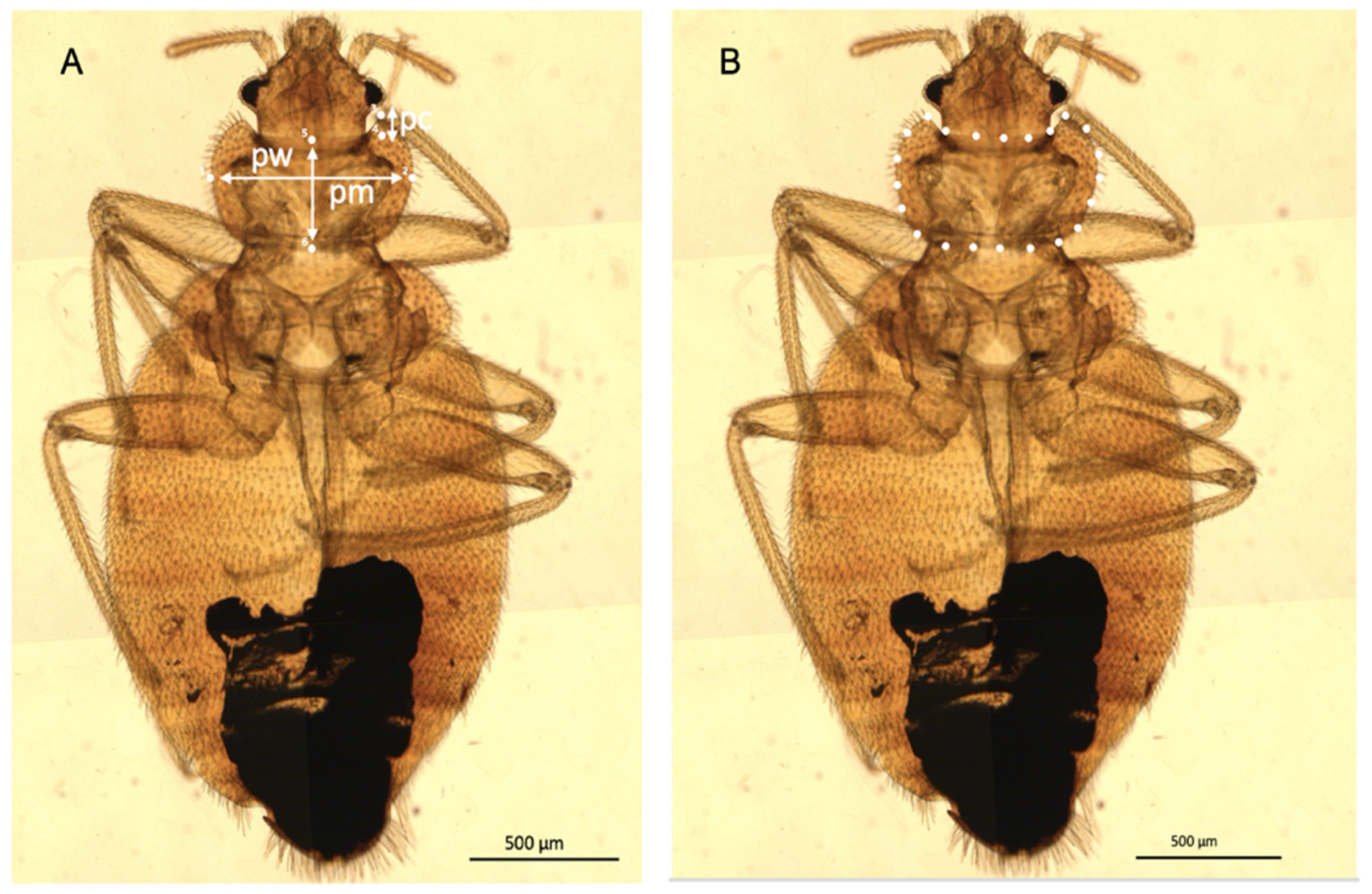
2.3. Linear Morphometric Analysis
2.4. Outline-Based Morphometrics
2.5. Clustering and Reclassification
2.6. Allometry
2.7. Software
3. Results
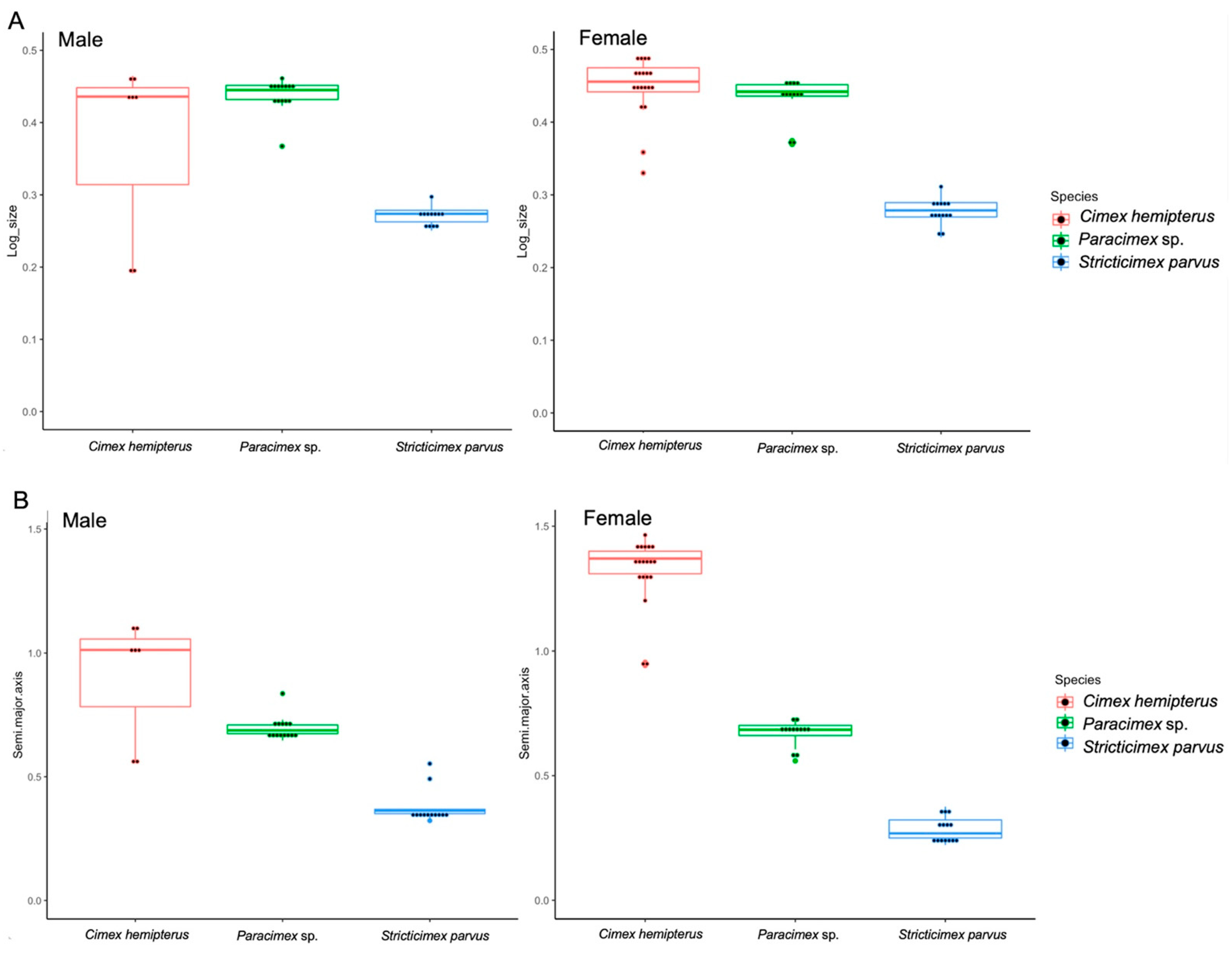
3.1. Size Variation of the Ectoparasites
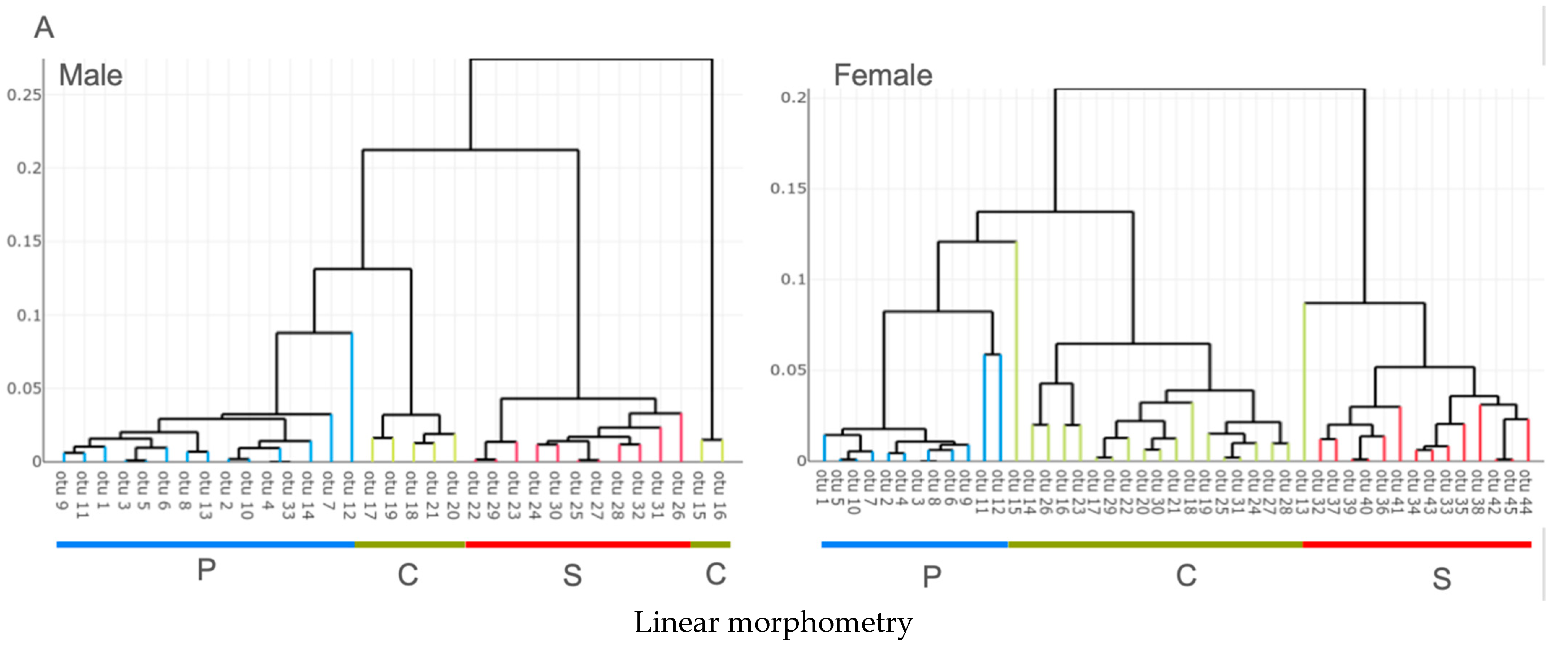
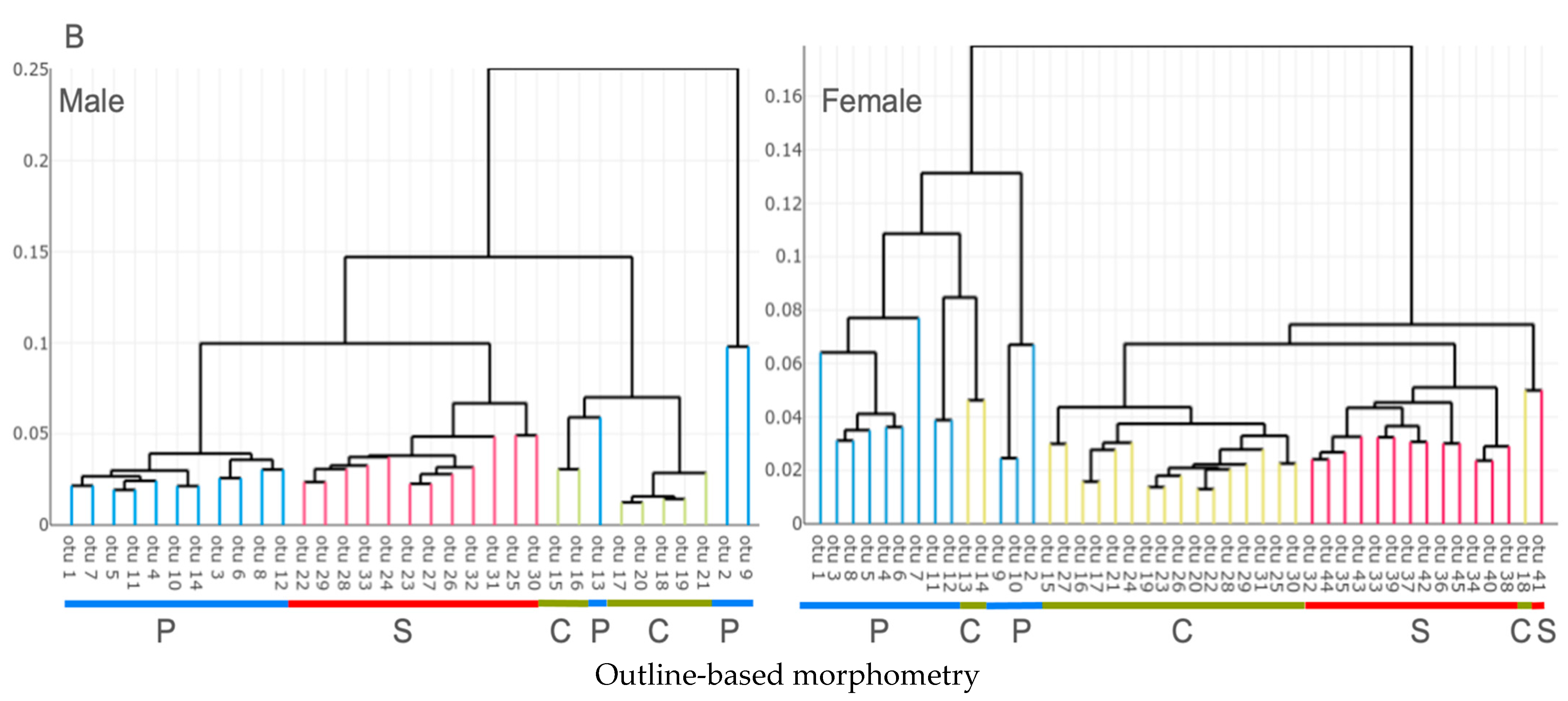
3.2. Unsupervised Classification
3.3. Supervised Classification
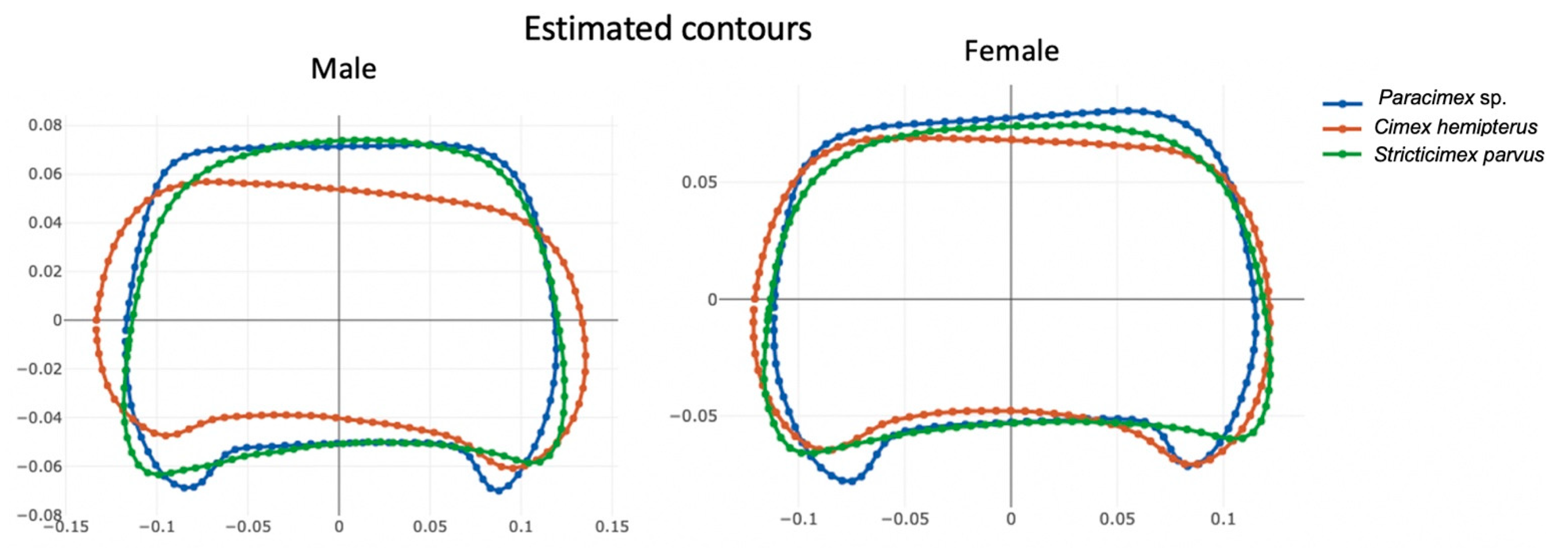
3.4. Allometry
4. Discussion
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burton, G.J. Bedbugs in relation to transmission of human diseases: Review of the literature. Public Health Rep. 1963, 78, 513–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietri, J.E. Case not closed: Arguments for new studies of the interactions between bed bugs and human pathogens. Am. J. Trop. Med. Hyg. 2020, 103, 619–624. [Google Scholar] [CrossRef]
- Sheele, J.M.; Libertin, C.R.; Pritt, B.S.; Wysokinska, E.M.; Pietri, J.E. Investigating the association of bed bugs with infectious diseases: A retrospective case-control study. Heliyon 2021, 7, e08107. [Google Scholar] [CrossRef] [PubMed]
- Doggett, S.L.; Dwyer, D.E.; Peñas, P.F.; Russell, R.C. Bed bugs: Clinical relevance and control options. Clin. Microbiol. Rev. 2012, 25, 164–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaunay, P.; Blanc, V.; Del Giudice, P.; Levy-Bencheton, A.; Chosidow, O.; Marty, P.; Brouqui, P. Bedbugs and infectious diseases. Clin. Infect. Dis. 2011, 52, 200–210. [Google Scholar] [CrossRef]
- Williams, J.E.; Imlarp, S.; Top, F.H., Jr.; Cavanaugh, D.C.; Russell, P.K. Kaeng Khoi Virus from naturally infected bedbugs (Cimicidae) and immature free-tailed bats. Bull. World Health Organ. 1976, 53, 365–369. [Google Scholar]
- Beatty, N.L.; Klotz, S.A.; Elliott, S.P. Hematophagous Ectoparasites of Cliff Swallows Invade a Hospital and Feed on Humans. Clin. Infect. Dis. 2017, 65, 2119–2121. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, K.; Harder, A.; Holland, S.; Hooper, J.; Leake-Lyall, C. Who knows the bed bug? Knowledge of adult bed bug appearance increases with people’s age in three counties of Great Britain. J. Med. Entomol. 2008, 45, 956–958. [Google Scholar] [CrossRef]
- Goddard, J.; Baker, G.T.; Ferrari, F.G.; Ferrari, C. Bed bugs (Cimex lectularius) and bat bugs (several Cimex species): A confusing issue. Outlooks Pest Manag. 2012, 23, 125–127. [Google Scholar] [CrossRef]
- Johnson, C. Taxonomic characters, variability, and relative growth in Cimex lectularius L. and C. columbarius Jenyns (Heteropt. Cimicidae). Trans. R. Entomol. Soc. Lond. 1939, 89, 543–568. [Google Scholar] [CrossRef]
- Kassianoff, L. Étude morphologique et biologique de la famille des cimicidés. Ann. Parasitol. Hum. Comparée 1937, 15, 97–124. [Google Scholar] [CrossRef]
- Usinger, R.L. Monograph of Cimicidae (Hemiptera, Heteroptera); Entomological Society of America: Annapolis, MA, USA, 1966. [Google Scholar]
- Balvín, O.; Munclinger, P.; Kratochvíl, L.; Vilímová, J. Mitochondrial DNA and morphology show independent evolutionary histories of bedbug Cimex lectularius (Heteroptera: Cimicidae) on bats and humans. J. Parasitol. Res. 2012, 111, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Ossa, G.; Johnson, J.S.; Puisto, A.I.; Rinne, V.; Sääksjärvi, I.E.; Waag, A.; Vesterinen, E.J.; Lilley, T.M. The Klingon batbugs: Morphological adaptations in the primitive bat bugs, Bucimex chilensis and Primicimex cavernis, including updated phylogeny of Cimicidae. Ecol. Evol. 2019, 9, 1736–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, S.; Hadani, A.; Lavi, A.G.; Berman, E.; Bendheim, U.; Hisham, A. The occurrence of the tropical bedbug (Cimex hemipterus, Fabricius) in poultry barns in Israel. Avian Pathol. 1987, 16, 339–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, T. Biodiversity of Heteroptera. In Insect Biodiversity: Science and Society; Foottit, R., Adler, P., Eds.; Blackwell Publishing: Oxford, UK, 2009; pp. 223–254. [Google Scholar]
- Stutt, A.D.; Siva-Jothy, M.T. Traumatic insemination and sexual conflict in the bed bug Cimex lectularius. Proc. Natl. Acad. Sci. USA 2001, 98, 5683–5687. [Google Scholar] [CrossRef] [Green Version]
- Ueshima, N. Experiments on reproductive isolation in Cimex lectularius and Cimex columbarius. Pan-Pac. Entomol. 1964, 4, 47–53. [Google Scholar]
- Potiwat, R.; Sungvornyothin, S.; Samung, Y.; Payakkapol, A.; Apiwathnasorn, C. Identification of Bat Ectoparasite Leptocimex inordinatus from Bat-dwelling Cave, Kanchanaburi Province, Thailand. Southeast Asian J. Trop. Med. Public Health 2016, 47, 16–22. [Google Scholar]
- Hornok, S.; Szőke, K.; Boldogh, S.A.; Sándor, A.D.; Kontschán, J.; Tu, V.T.; Halajian, A.; Takács, N.; Görföl, T.; Estók, P. Phylogenetic analyses of bat-associated bugs (Hemiptera: Cimicidae: Cimicinae and Cacodminae) indicate two new species close to Cimex lectularius. Parasites Vectors 2017, 10, 439. [Google Scholar] [CrossRef] [Green Version]
- Roth, S.; Balvín, O.; Siva-Jothy, M.T.; Di Iorio, O.; Benda, P.; Calva, O.; Faundez, E.I.; Khan, F.A.A.; McFadzen, M.; Lehnert, M.P.; et al. Bedbugs evolved before their bat hosts and did not co-speciate with ancient humans. Curr. Biol. 2019, 29, 1847–1853. [Google Scholar] [CrossRef]
- Balvin, O.; Roth, S.; Vilimova, J. Molecular evidence places the swallow bug genus Oeciacus Stål within the bat and bed bug genus Cimex Linnaeus (Heteroptera: Cimicidae). Syst. Entomol. 2015, 40, 652–665. [Google Scholar] [CrossRef]
- Balvín, O.; Vilímová, J.; Kratochvíl, L. Batbugs (Cimex pipistrelli group, Heteroptera: Cimicidae) are morphologically, but not genetically differentiated among bat hosts. J. Zool. Syst. Evol. Res. 2013, 51, 287–295. [Google Scholar] [CrossRef]
- Benkacimi, L.; Gazelle, G.; El Hamzaoui, B.; Bérenger, J.-M.; Parola, P.; Laroche, M. MALDI-TOF MS identification of Cimex lectularius and Cimex hemipterus bedbugs. Infect. Genet. Evol. 2020, 85, 104536. [Google Scholar] [CrossRef] [PubMed]
- Chabriere, E.; Bassène, H.; Drancourt, M.; Sokhna, C. MALDI-TOF MS and point of care are disruptive diagnostic tools in Africa. New Microbes New Infect. 2018, 26, S83–S88. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, K.; Jacobs, D. Abundance of Cacodmus villosus (Stål, 1855) (Heteroptera: Cimicidae) on its host, Neoromicia capensis (Chiroptera: Vespertillionidae). Afr. Entomol. 2006, 14, 398–400. Available online: https://hdl.handle.net/10520/EJC32679 (accessed on 1 November 2022).
- Campbell, B.; Koehler, P.; Buss, L.; Baldwin, R. Recent Documentation of the Tropical Bed Bug (Hemiptera: Cimicidae) in Florida Since the Common Bed Bug Resurgence. Fla. Entomol. 2016, 99, 549–551. [Google Scholar] [CrossRef] [Green Version]
- Klingenberg, C.P. Multivariate allometry. In Advances in Morphometrics; Springer: Boston, MA, USA, 1996; pp. 23–49. [Google Scholar] [CrossRef]
- Darroch, J.N.; Mosimann, J.E. Canonical and principal components of shape. Biometrika 1985, 72, 241–252. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Marcus, L.F. A revolution morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Dujardin, J.-P. Modern morphometrics of medically important insects. In Genetics and Evolution of Infectious Disease; Elsevier: Amsterdam, The Netherlands, 2011; pp. 473–501. [Google Scholar] [CrossRef]
- Santillán-Guayasamín, S.; Villacís, A.G.; Grijalva, M.J.; Dujardin, J.-P. The modern morphometric approach to identify eggs of Triatominae. Parasites Vectors 2017, 10, 55. [Google Scholar] [CrossRef] [Green Version]
- Deku, G.; Combey, R.; Doggett, S.L. Morphometrics of the Tropical Bed Bug (Hemiptera: Cimicidae) From Cape Coast, Ghana. J. Med. Entomol. 2022, 59, 1534–1547. [Google Scholar] [CrossRef]
- Dujardin, J.-P.; Slice, D.E. Contributions of Morphometrics to Medical Entomology. In Encyclopedia of Infectious Diseases; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2007; pp. 435–447. [Google Scholar] [CrossRef]
- Chittsamart, B.; Samruayphol, S.; Sungvorayothin, S.; Pothiwat, R.; Samung, Y.; Apiwathnasorn, C. Phlebotomine sand flies of edible-nest swiftlet cave of Lang Ga Jiew Island, Chumphon province, Thailand. Trop. Biomed. 2015, 32, 402–406. [Google Scholar]
- Ueshima, N. New species and records of Cimicidae with keys (Hemiptera). Pan-Pac. Entomol. 1968, 44, 264–279. [Google Scholar]
- Cielecka, D.; Salamatin, R.; Garbacewicz, A. Usage of the Hoyer’s medium for diagnostics and morphological studies of some parasites. Wiad. Parazytol. 2009, 55, 265–270. [Google Scholar] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Dujardin, S.; Dujardin, J.-P. Geometric morphometrics in the cloud. Infect. Genet. Evol. 2019, 70, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, F.P.; Giardina, C.R. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process. 1982, 18, 236–258. [Google Scholar] [CrossRef]
- Dujardin, J.-P.; Dujardin, S.; Kaba, D.; Santillán-Guayasamin, S.; Villacís, A.G.; Piyaselakul, S.; Sumruayphol, S.; Samung, Y.; Vargas, R.M. The maximum likelihood identification method applied to insect morphometric data. Zool. Syst. 2017, 42, 46–58. [Google Scholar]
- Rumelhart, D.E.; Hinton, G.E.; Williams, R.J. Learning representations by back-propagating errors. Nature 1986, 323, 533–536. [Google Scholar] [CrossRef]
- Mancuso, S. Elliptic Fourier Analysis (EFA) and Artificial Neural Networks (ANNs) for the identification of grapevine (Vitis vinifera L.) genotypes. Vitis 1999, 38, 73–78. [Google Scholar] [CrossRef]
- Sumruayphol, S.; Siribat, P.; Dujardin, J.-P.; Dujardin, S.; Komalamisra, C.; Thaenkham, U. Fasciola gigantica, F. hepatica and Fasciola intermediate forms: Geometric morphometrics and an artificial neural network to help morphological identification. PeerJ 2020, 8, e8597. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, G.H. Multivariate analysis and the study of form, with special reference to canonical variate analysis. Am. Zool. 1980, 20, 679–693. [Google Scholar] [CrossRef]
- Soda, K.; Slice, D.; Naylor, G. Artificial neural networks and geometric morphometric methods as a means for classification: A case-study using teeth from Carcharhinus sp. (Carcharhinidae). J. Morphol. 2017, 278, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Jenyns, L. On three undescribed species of the genus Cimex, closely allied to the common bed bug. Ann. Mag. Nat. Hist. 1839, 3, 241–244. [Google Scholar] [CrossRef] [Green Version]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; The Press Syndicate of the University of Cambridge: Cambridge, UK, 1997. [Google Scholar]
- Dujardin, J.-P.; Kaba, D.; Solano, P.; Dupraz, M.; McCoy, K.D.; Jaramillo-O, N. Outline-based morphometrics, an overlooked method in arthropod studies? Infect. Genet. Evol. 2014, 28, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Tawatsin, A.; Thavara, U.; Chompoosri, J.; Phusup, Y.; Jonjang, N.; Khumsawads, C.; Bhakdeenuan, P.; Sawanpanyalert, P.; Asavadachanukorn, P.; Mulla, M.S. Insecticide resistance in bedbugs in Thailand and laboratory evaluation of insecticides for the control of Cimex hemipterus and Cimex lectularius (Hemiptera: Cimicidae). J. Med. Entomol. 2011, 48, 1023–1030. [Google Scholar] [CrossRef]
- Fagre, A.C.; Kading, R.C. Can bats serve as reservoirs for arboviruses? Viruses 2019, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Monath, T.; Lazuick, J.; Cropp, C.; Rush, W.; Calisher, C.; Kinney, R.; Trent, D.; Kemp, G.; Bowen, G.; Francy, D. Recovery of Tonate Virus (“Bijou Bridge” strain), a member of the Venezuelan Equine Encephalomyelitis Virus complex, from cliff swallow nest bugs (Oeciacus vicarius) and nestling birds in North America. Am. J. Trop. Med. Hyg. 1980, 29, 969–983. [Google Scholar] [CrossRef]
- Rush, W.A.; Francy, D.B.; Smith, G.C.; Cropp, C.B. Transmission of an Arbovirus by a Member of the Family Cimicidae. Ann. Entomol. Soc. Am. 1980, 73, 315–318. [Google Scholar] [CrossRef]
- Dujardin, J.-P.A.; Kaba, D.; Henry, A.B. The exchangeability of shape. BMC Res. Notes 2010, 3, 266. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Linear Morphometry | Outline-Based Morphometry | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Paracimex sp. | 0.44 ± 0.02 a (n = 14) | 0.43 ± 0.03a (n = 12) | 0.70 ± 0.05 a (n = 14) | 0.70 ± 0.05 a (n = 12) |
| Cimex hemipterus | 0.37 ± 0.12 a (n = 7) | 0.45 ± 0.04 a,* (n = 19) | 0.90 ± 0.24 a (n = 7) | 1.32 ± 0.14 b,* (n = 19) |
| Stricticimex parvus | 0.30 ± 0.01 b (n = 12) | 0.30 ± 0.02 b (n = 14) | 0.40 ± 0.07 b (n = 12) | 0.30 ± 0.04 c,* (n = 14) |
| Validated Reclassification Scores | Linear Morphometry | Outline-Based Morphometry | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Maximum likelihood size-based reclassification score | 61% (20/33) | 71% (32/45) | 85% (28/33) | 82% (37/45) |
| Multilayer perceptron shape-based reclassification score | 99% ± 0.7% | 94% ± 0.5% | 96% ± 0.5% | 98% ± 0.7% |
| Males | Females | |
|---|---|---|
| Linear morphometry | ||
| PC1 | 91% | 91% |
| PC2 | 8% | 5% |
| Outline-based morphometry | ||
| PC1 | 15% | 2% |
| PC2 | 66% | 37% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajonhson, D.M.; Raksakoon, C.; Payakkapol, A.; Dujardin, S.; Dujardin, J.-P.; Potiwat, R. Comparison of Two Different Morphological Methods to Study the Pronotum of Cimicidae: Bed Bugs, Bat Bugs, and Swallow Bugs. Insects 2022, 13, 1155. https://doi.org/10.3390/insects13121155
Rajonhson DM, Raksakoon C, Payakkapol A, Dujardin S, Dujardin J-P, Potiwat R. Comparison of Two Different Morphological Methods to Study the Pronotum of Cimicidae: Bed Bugs, Bat Bugs, and Swallow Bugs. Insects. 2022; 13(12):1155. https://doi.org/10.3390/insects13121155
Chicago/Turabian StyleRajonhson, Dora M., Chadchalerm Raksakoon, Anon Payakkapol, Sébastien Dujardin, Jean-Pierre Dujardin, and Rutcharin Potiwat. 2022. "Comparison of Two Different Morphological Methods to Study the Pronotum of Cimicidae: Bed Bugs, Bat Bugs, and Swallow Bugs" Insects 13, no. 12: 1155. https://doi.org/10.3390/insects13121155
APA StyleRajonhson, D. M., Raksakoon, C., Payakkapol, A., Dujardin, S., Dujardin, J. -P., & Potiwat, R. (2022). Comparison of Two Different Morphological Methods to Study the Pronotum of Cimicidae: Bed Bugs, Bat Bugs, and Swallow Bugs. Insects, 13(12), 1155. https://doi.org/10.3390/insects13121155