
Systematic Position of the Fossil Burrower Bug Eocenocydnus lisi Popov, 2019 (Hemiptera, Heteroptera, Cydnidae) Revealed by a Parallel/Cross-Eyed Viewing Method Used for Obtaining Three-Dimensional Images


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
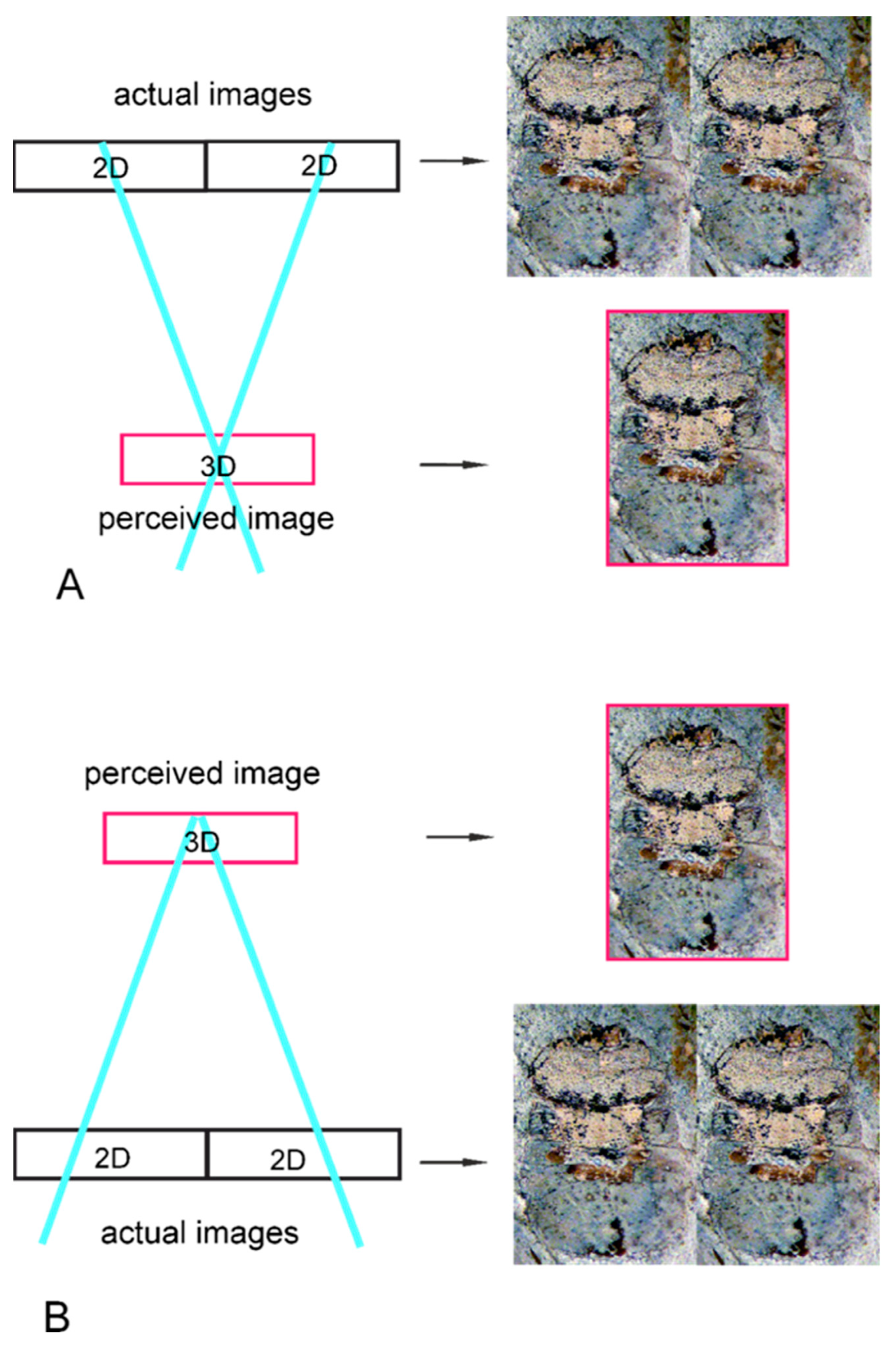
2. Materials and Methods
3. Results and Discussion
3.1. Redefinition of the Genus Eocenocydnus
3.2. Systematic Position of the Genus Eocenocydnus
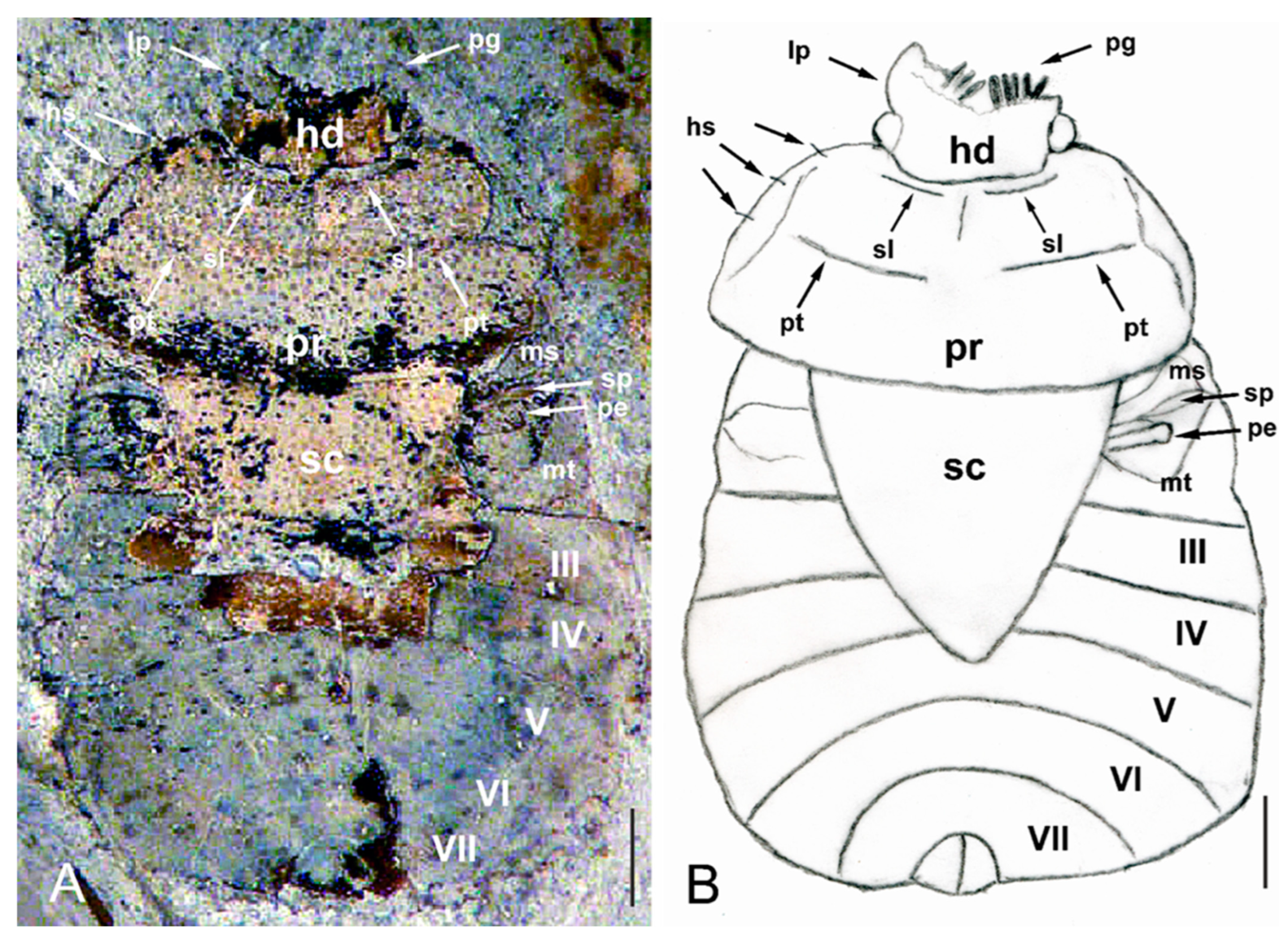
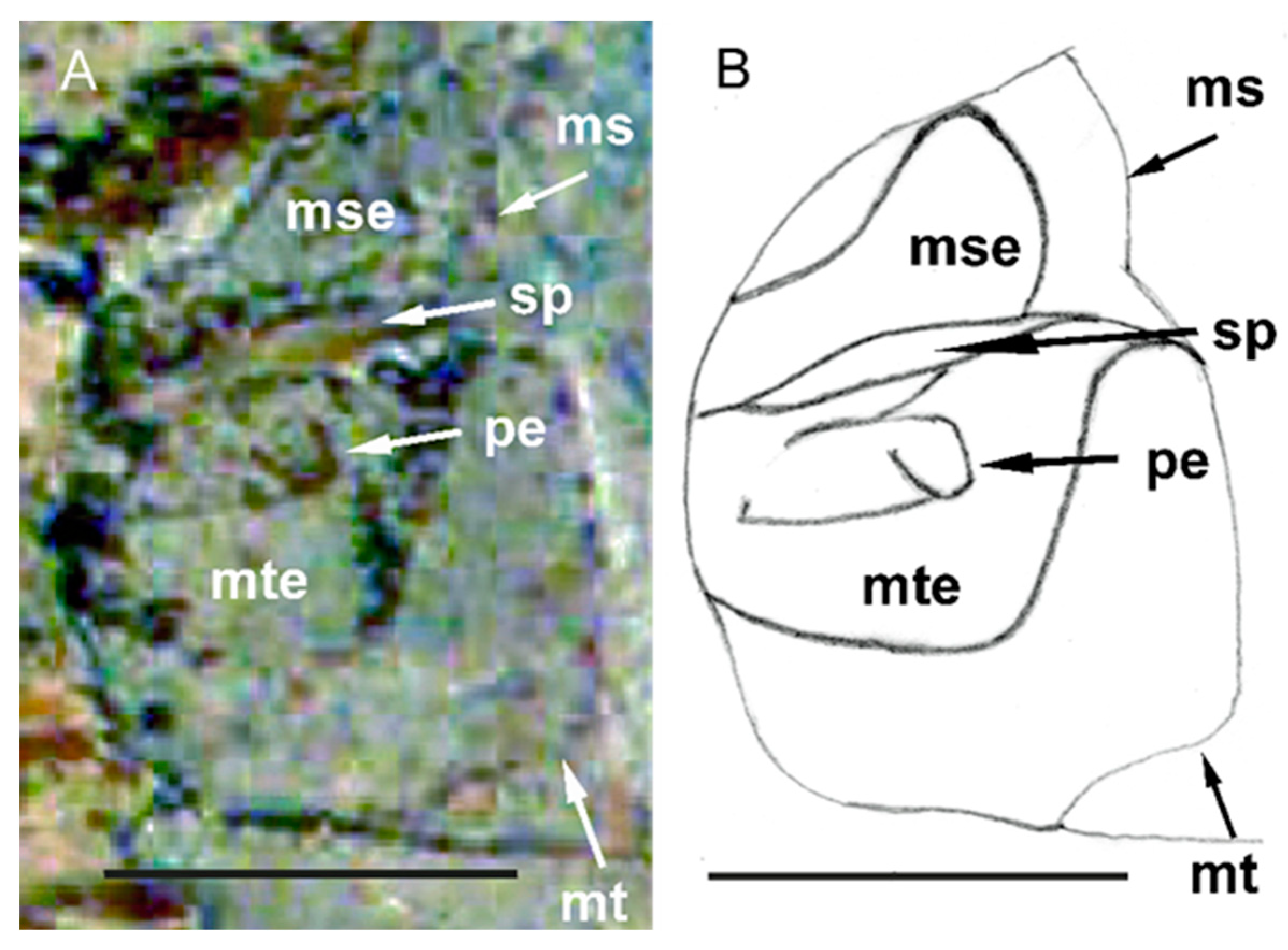
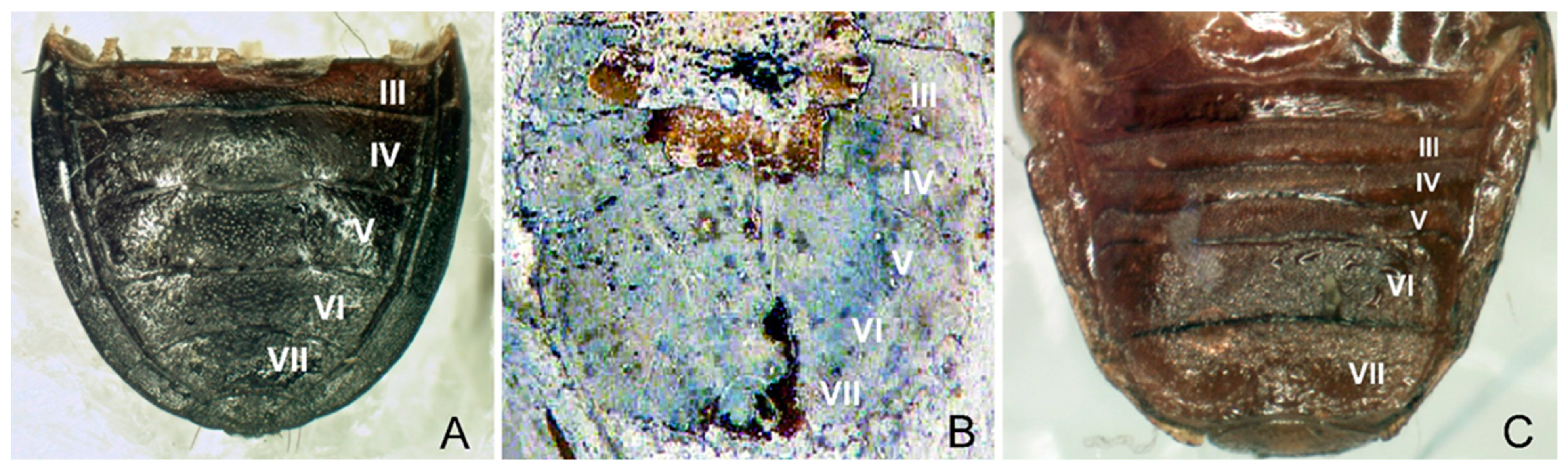
3.3. Redescription of Eocenocydnus lisi
4. Systematic Palaeontology
- -
- The body does not exceed 13.00 mm in length;
- -
- The head margins are armed predominantly by pegs and peg-like setae, although hair-like setae are also present;
- -
- The scutellum is short, triangular, usually not reaching the half-length of the hemelytra, and broader than or as broad as long;
- -
- The peritreme is extended laterally in the form of a polished band, which is sometimes strongly recurved at its end.
5. Conclusions
- The fossil burrower bug genus Eocenocydnus, placed in the original description [7] tentatively within the subfamily Sehirinae, was transferred to the Cydninae tribe Cydnini.
- A parallel, cross-eyed viewing method used for the first time in insect fossil morphological studies allowed the recognition of more morphological details than when directly viewing a two-dimensional flat image.
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vršanský, P.; Lis, J.A.; Schlögl, J.; Guldan, M.; Mlynský, T.; Barna, P.; Štys, P. Partially disarticulated new Miocene burrower bug (Hemiptera: Heteroptera: Cydnidae) from Cerová (Slovakia) documents occasional preservation of terrestrial arthropods in deep-marine sediments. Eur. J. Entomol. 2015, 112, 844–854. [Google Scholar] [CrossRef] [Green Version]
- Lis, J.A.; Lis, B.; Heiss, E. Chilamnestocoris mixtus gen. et spec. nov., the first burrower bug (Hemiptera: Pentatomoidea: Cydnidae) in Upper Cretaceous Burmese amber. Cretac. Res. 2018, 91, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Du, S.L.; Yao, Y.Z.; Ren, D. A new genus and species of burrower bugs (Heteroptera: Cydnidae) from the mid-Cretaceous Burmese amber. Zootaxa 2019, 4585, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Lis, J.A.; Roca-Cusachs, M.; Lis, B.; Jung, S.H. Pullneyocoris dentatus gen. et sp. nov. (Hemiptera: Pentatomoidea: Cydnidae), the third representative of the subfamily Amnestinae from mid-Cretaceous amber of northern Myanmar. Cretac. Res. 2020, 115, 104562. [Google Scholar] [CrossRef]
- Du, S.; Gu, L.; Engel, M.S.; Ren, D.; Yao, Y. Morphological phylogeny of new Cretaceous fossils elucidates the early history of soil dwelling among bugs. Front. Ecol. Evol. 2022, 10, 908044. [Google Scholar] [CrossRef]
- Statz, G.; Wagner, E. Geocorisae (landwanzen) aus den Oberoligocanen Ablagerungen von Rott. Palaeontogr. Abteil. A 1950, 98, 97–136. [Google Scholar]
- Szwedo, J.; Drohojowska, J.; Popov, Y.; Simon, E.; Wegierek, P. Aphids, true hoppers, jumping plant-lice, scale insects, true bugs and whiteflies (Insecta: Hemiptera) from the Insect Limestone (latest Eocene) of the Isle of Wight, UK. Earth Environ. Sci. Trans. R. Soc. Edinb. 2019, 110, 331–396. [Google Scholar] [CrossRef]
- Schuh, R.T.; Weirauch, C.H. True bugs of the World (Hemiptera: Heteroptera). Classification and Natural History, 2nd ed.; Monograph Series; Siri Scientific Press: Manchester, UK, 2020; Volume 8, 767p. [Google Scholar]
- Munt, M.; Museum of Isle of Wight Geology, Sandown, UK. Personal communication, 2022.
- Adams, L.P. Stereoscopic viewing of image pairs with the naked eyes. Photogramm. Rec. 1974, 8, 229–230. [Google Scholar] [CrossRef]
- Green, N.M. Stereo viewing simplified. Nature 1983, 301, 279. [Google Scholar] [CrossRef]
- Green, N.M. Stereoimages—A practical approach. Structure 1994, 2, 85–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAllister, D.F. Stereo and 3-D Display Technology. In Encyclopedia of Imaging Science and Technology; John Wiley & Sons: Hoboken, NJ, USA, 2002; pp. 1327–1344. [Google Scholar]
- Çöltekin, A. Foveation for 3D Visualization and Stereo Imaging. Ph.D. Thesis, Helsinki University of Technology, Espoo, Finland, 3 February 2006. [Google Scholar]
- Ono, H.; Lillakas, L.; Wade, N.J. Seeing double and depth with Wheatstone’s stereograms. Perception 2007, 36, 1611–1623. [Google Scholar] [CrossRef] [PubMed]
- Imura, J. New and obscure species of the genus Chilocorus Mayr in eastern Asia, with the proposal of a “nitidus-group” concept (Hemiptera: Heteroptera: Cydnidae). Zootaxa 2011, 3012, 31–47. [Google Scholar] [CrossRef]
- Lis, J.A.; Pluot-Sigwalt, D. Nymphal and adult cephalic chaetotaxy of the Cydnidae (Hemiptera: Heteroptera), and its adaptive, taxonomic and phylogenetic significance. Eur. J. Entomol. 2002, 99, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Kment, P.; Vilímová, J. Thoracic scent efferent system of Pentatomoidea (Hemiptera: Heteroptera): A review of terminology. Zootaxa 2010, 2706, 1–77. [Google Scholar] [CrossRef] [Green Version]
- Lis, J.A. Studies on Cydnidae of the Australian Region V. The genus Chilocoris Mayr in Australia, Tasmania and Moluccas (Heteroptera). Bonn. Zool. Beitr. 1994, 45, 225–230. [Google Scholar]
- Lis, J.A. Studies on Cydnidae of the Australian Region. X. Two new species of the genus Chilocoris Mayr from New Guinea (Hemiptera: Heteroptera). Int. J. Invertebr. Taxon. Genus 1997, 8, 583–589. [Google Scholar]
- Lis, J.A. Three new Australian genera of burrower bugs with four new species (Heteroptera: Cydnidae). Pol. Pismo ent. 1997, 66, 189–201. [Google Scholar]
- Lis, J.A. The genus Chilocoris (Heteroptera: Cydnidae) in Australia. Acta Soc. Zool. Bohem. 1999, 63, 139–155. [Google Scholar]
- Froeschner, R.C. Cydnidae of the Western Hemisphere. Proc. U. S. Natl. Mus. 1960, 111, 337–680. [Google Scholar] [CrossRef]
- Josifov, M.V. Fauna Bulgarica 12. Heteroptera, Pentatomoidea; Academia Scientiarum Bulgariae: Sofia, Bulgaria, 1981; 205p. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lis, J.A. Systematic Position of the Fossil Burrower Bug Eocenocydnus lisi Popov, 2019 (Hemiptera, Heteroptera, Cydnidae) Revealed by a Parallel/Cross-Eyed Viewing Method Used for Obtaining Three-Dimensional Images. Insects 2023, 14, 22. https://doi.org/10.3390/insects14010022
Lis JA. Systematic Position of the Fossil Burrower Bug Eocenocydnus lisi Popov, 2019 (Hemiptera, Heteroptera, Cydnidae) Revealed by a Parallel/Cross-Eyed Viewing Method Used for Obtaining Three-Dimensional Images. Insects. 2023; 14(1):22. https://doi.org/10.3390/insects14010022
Chicago/Turabian StyleLis, Jerzy A. 2023. "Systematic Position of the Fossil Burrower Bug Eocenocydnus lisi Popov, 2019 (Hemiptera, Heteroptera, Cydnidae) Revealed by a Parallel/Cross-Eyed Viewing Method Used for Obtaining Three-Dimensional Images" Insects 14, no. 1: 22. https://doi.org/10.3390/insects14010022
APA StyleLis, J. A. (2023). Systematic Position of the Fossil Burrower Bug Eocenocydnus lisi Popov, 2019 (Hemiptera, Heteroptera, Cydnidae) Revealed by a Parallel/Cross-Eyed Viewing Method Used for Obtaining Three-Dimensional Images. Insects, 14(1), 22. https://doi.org/10.3390/insects14010022