


Labial Sensory Organs of Two Leptoglossus Species (Hemiptera: Coreidae): Their Morphology and Supposed Function


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Materials Examined
2.2. Light Microscopy Procedures
2.3. SEM Procedures
2.4. Measurements
2.5. Terminology for the Sensilla
3. Results
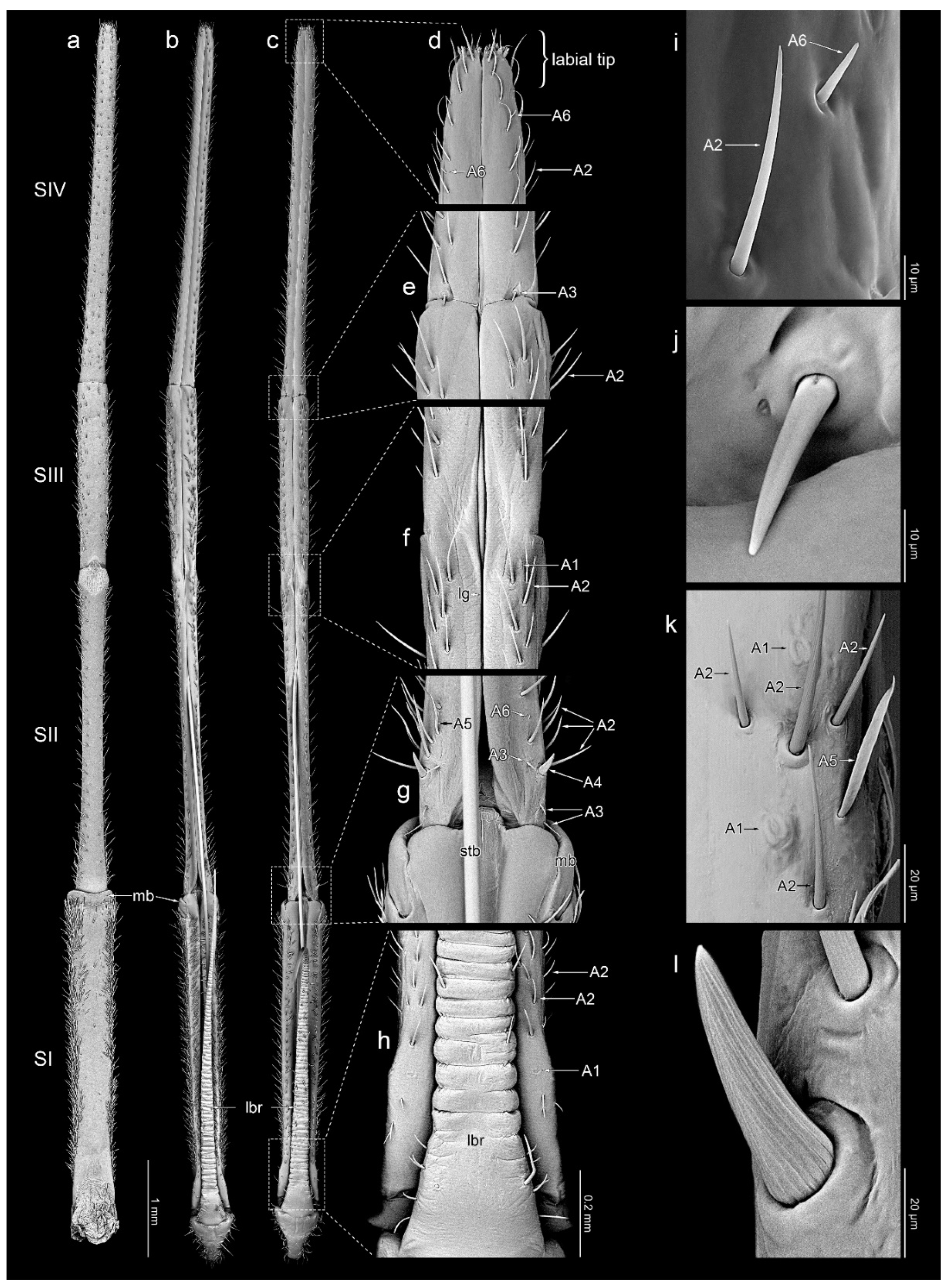
3.1. Morphology of Labium
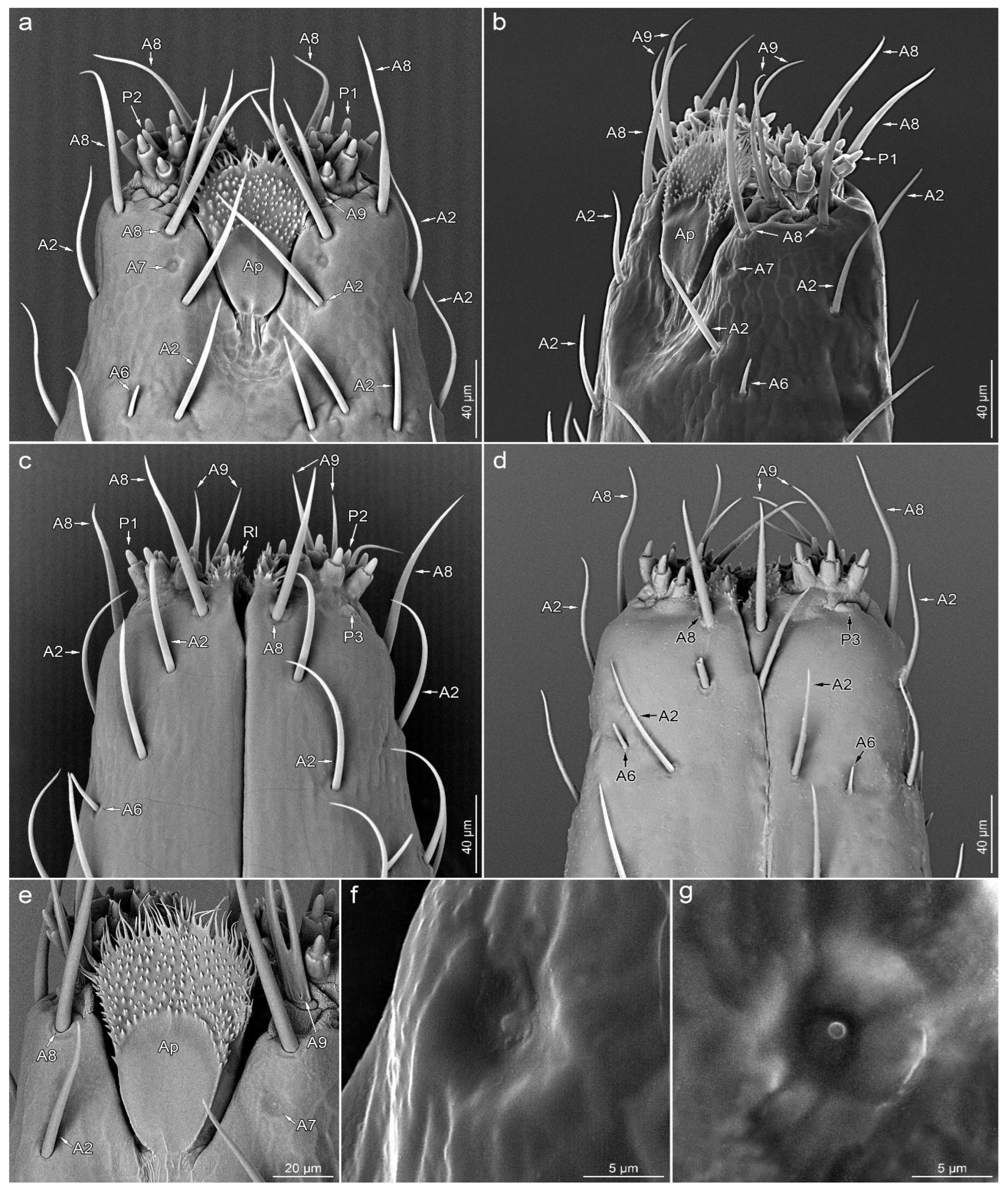
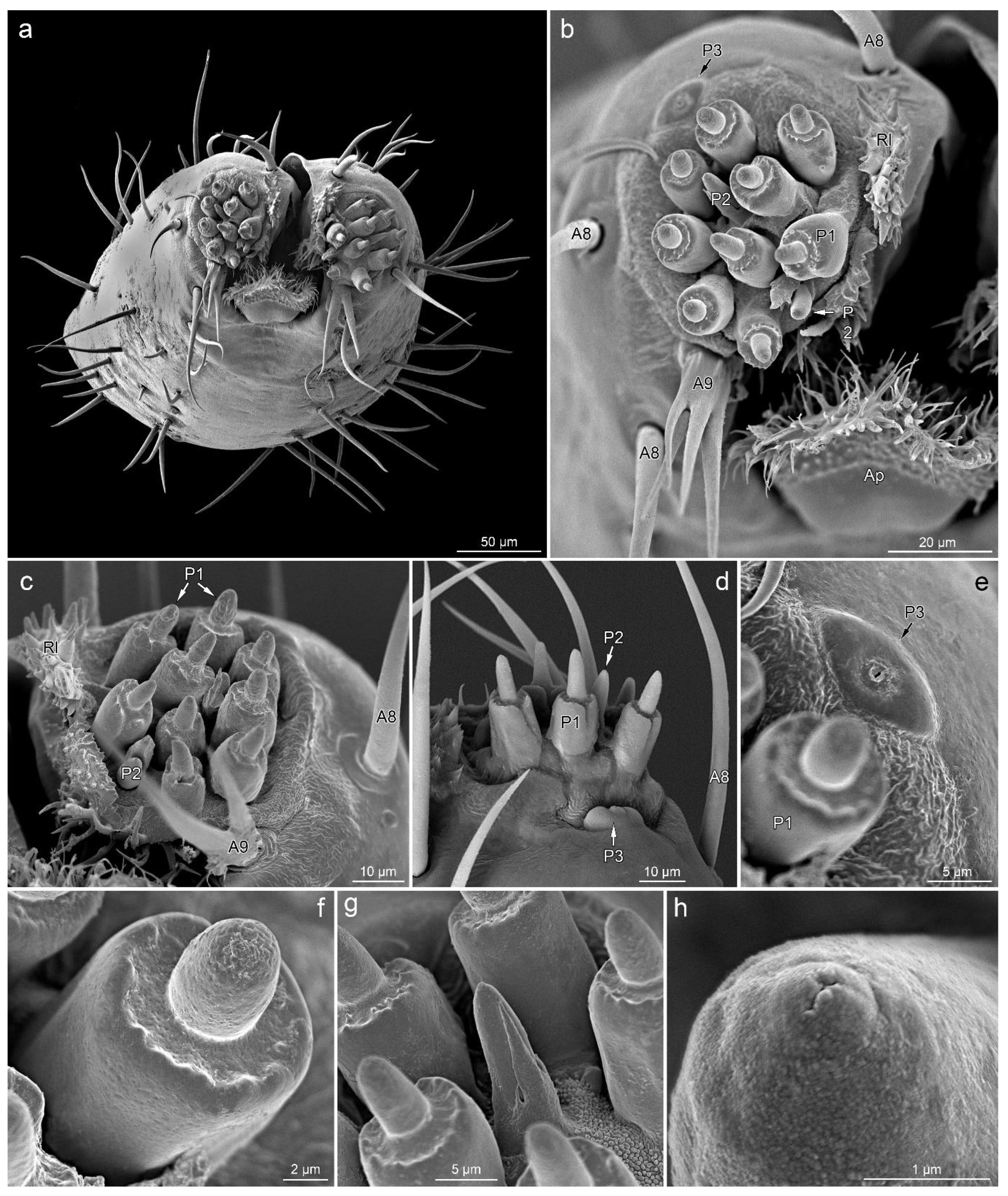
3.2. Sensilla Types
3.2.1. Aporous Sensilla
3.2.2. Porous Sensilla
3.3. Sensilla Arrangement
3.3.1. Segment I
3.3.2. Segment II
3.3.3. Segment III
3.3.4. Segment IV
3.4. Other Cuticular Structures
4. Discussion
4.1. Mechanoreception
4.2. Chemoreception
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Henry, T.J. Biodiversity of Heteroptera. In Insect Biodiversity: Science and Society; Foottit, R.G., Adler, P.H., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 279–335. [Google Scholar]
- Schuh, R.T.; Weirauch, C. True Bugs of the World (Hemiptera: Heteroptera): Classification and Natural History, 2nd ed.; Siri Scientific Press Monograph Series; Siri Scientific Press: Rochdale, UK, 2020; Volume 8, 800p. [Google Scholar]
- Schaefer, C.W.; Panizzi, A.R. (Eds.) Heteroptera of Economic Importance; CRC Press: London, UK, 2000. [Google Scholar]
- Millar, J.G.; Zou, Y.; Hall, D.R.; Halloran, S.; Pajares, J.A.; Ponce-Herrero, L.; Shates, T.; Wilson, H.; Daane, K.M. Identification and Synthesis of Leptotriene, a Unique Sesquiterpene Hydrocarbon from Males of the Leaffooted Bugs Leptoglossus zonatus and L. occidentalis. J. Nat. Prod. 2022, 85, 2062–2070. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.C. A revision of the genus Leptoglossus Guérin (Hemiptera: Coreidae). Entomol. Am. 1969, 45, 35–140. [Google Scholar]
- Brailovsky, H. Illustrated key for identification of the species included in the genus Leptoglossus (Hemiptera: Heteroptera: Coreidae: Coreinae: Anisoscelini), and descriptions of five new species and new synonyms. Zootaxa 2014, 3794, 143–178. [Google Scholar] [CrossRef] [Green Version]
- Heidemann, O. New species of Leptoglossus from North America. Proc. Entomol. Soc. Wash. 1910, 12, 191–197. [Google Scholar]
- Lesieur, V.; Lombaert, E.; Guillemaud, T.; Courtial, B.; Strong, W.; Roques, A.; Auger-Rozenberg, M.-A. The rapid spread of Leptoglossus occidentalis in Europe: A bridgehead invasion. J. Pest Sci. 2018, 92, 189–200. [Google Scholar] [CrossRef]
- CoreoideaSF Team. Coreoidea Species File Online. Version 5.0/5.0. Available online: http://Coreoidea.SpeciesFile.org (accessed on 14 September 2022).
- Bates, S.L.; Borden, J.H.; Kermode, A.R.; Bennett, R.G. Impact of Leptoglossus occidentalis (Hemiptera: Coreidae) on Douglas-Fir Seed Production. J. Econ. Entomol. 2000, 93, 1444–1451. [Google Scholar] [CrossRef]
- Strong, W. Lodgepole pine seedset increase by mesh bagging is due to Leptoglossus occidentalis (Hemiptera: Coreidae) exclusion. J. Entomol. Soc. Br. Columbia 2015, 112, 3–18. [Google Scholar]
- Mutke, S.; Calama, R.; Nasrallah Neaymeh, E.; Roques, A. Impact of the Dry Cone Syndrome on commercial kernel yield of stone pine cones. In Mediterranean Pine Nuts from Forests and Plantations; Carrasquinho, I., Correia, A.C., Mutke, S., Eds.; CIHEAM Options Méditerranéennes: Série A, Séminaires Méditerranéens; FAO: Rome, Italy, 2017; Volume 122, pp. 154–196. [Google Scholar]
- Dallas, W.S. List of the Specimens of Hemipterous Insects in the Collection of the British Museum; Trustees of the British Museum: London, UK, 1852; pp. 369–592. [Google Scholar]
- Xiao, Y.; Fadamiro, H.Y. Host preference and development of Leptoglossus zonatus (Hemiptera: Coreidae) on satsuma mandarin. J. Econ. Entomol. 2009, 102, 1908–1914. [Google Scholar] [CrossRef]
- Daane, K.M.; Yokota, G.Y.; Wilson, H. Seasonal Dynamics of the Leaffooted Bug Leptoglossus zonatus and Its Implications for Control in Almonds and Pistachios. Insects 2019, 10, 255. [Google Scholar] [CrossRef] [Green Version]
- McGavin, G.C. Food and Feeding. In Bugs of the World; Blandford: London, UK, 1993. [Google Scholar]
- Butt, F.H. Comparative study of mouth parts of representative Hemiptera-Heteroptera. Ann. Entomol. Soc. Am. 1943, 38, 124. [Google Scholar] [CrossRef]
- Cobben, R.H. Evolutionary trends in Heteroptera. Part II. Mouthpart—Structures and feeding strategies, Mededelingen. Landbouwhogesch Wagening 1978, 78, 5–401. [Google Scholar]
- Muir, F.; Kershaw, J.C. On the Homologies and Mechanism of the Mouth-Parts of Hemiptera. Psyche A J. Entomol. 1911, 18, 101638. [Google Scholar] [CrossRef] [Green Version]
- Chapman, R.F. Mechanoreception. Chemoreception. Mouthparts and feeding. In The Insects, Structure and Function; Chapman, R.F., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 610–652. [Google Scholar]
- Krenn, H.W. (Ed.) Insect Mouthparts: Form, Function, Development and Performance; Springer Nature: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Backus, E.A. Sensory systems and behaviours which mediate hemipteran plant-feeding: A taxonomic overview. J. Insect Physiol. 1988, 34, 151–165. [Google Scholar] [CrossRef]
- Faucheux, M.; Kristensen, N.P.; Yen, S.-H. The antennae of neopseustid moths: Morphology and phylogenetic implications, with special reference to the sensilla (Insecta, Lepidoptera, Neopseustidae). Zool Anz. 2006, 245, 131–142. [Google Scholar] [CrossRef]
- Taszakowski, A.; Nowińska, A.; Brożek, J. Morphological study of the labial sensilla in Nabidae (Hemiptera: Heteroptera: Cimicomorpha). Zoomorphology 2019, 138, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Brożek, J. Morphology and arrangement of the labial sensilla of the water bugs. Bull. Insectol. 2008, 61, 167–168. [Google Scholar]
- Brożek, J. Comparative Analysis and Systematic Mapping of the Labial Sensilla in the Nepomorpha (Heteroptera: Insecta). Sci. World J. 2013, 2013, 518034. [Google Scholar] [CrossRef]
- Brożek, J.; Zettel, H. A comparison of the external morphology and functions of labial tip sensilla in semiaquatic bugs (Hemiptera: Heteroptera: Gerromorpha). Eur. J. Entomol. 2014, 111, 275–297. [Google Scholar] [CrossRef] [Green Version]
- Avé, D.; Frazier, J.L.; Hatfield, L.D. Contact chemoreception in the tarnished plant bug Lygus lineolaris. Entomol. Exp. Appl. 1978, 24, 17–27. [Google Scholar] [CrossRef]
- Hatfield, L.D.; Frazier, J.L. Ultrastructure of the labial tip sensilla of the tarnished plant bug, Lygus lineolaris (P. de Beauvois) (Hemiptera:Miridae). Int. J. Insect Morphol. Embryol. 1980, 9, 59–66. [Google Scholar] [CrossRef]
- Catalá, S. Sensilla associated with the rostrum of eight species of Triatominae. J. Morphol. 1996, 228, 195–201. [Google Scholar] [CrossRef]
- Singh, R.; Singh, K.; Prakash, S.; Mendki, M.; Rao, K. Sensory organs on the body parts of the bed-bug Cimex hemipterus fabricius (Hemiptera: Cimicidae) and the anatomy of its central nervous system. Int. J. Insect Morphol. Embryol. 1996, 25, 183–204. [Google Scholar] [CrossRef]
- Sinitsina, E.E.; Chaika, S.Y. Morphology of sensory organs on rostrum apex of terrestrial bugs (Hemiptera). Zool. J. 1997, 76, 294–303. [Google Scholar]
- Romani, R.; Salerno, G.; Frati, F.; Conti, E.; Isidoro, N.; Bin, F. Oviposition behaviour in Lygus rugulipennis: A morpho-functional study. Entomol. Exp. Appl. 2005, 115, 17–25. [Google Scholar] [CrossRef]
- Brożek, J.; Chłond, D. Morphology, arrangement and classification of sensilla on the apical segment of labium in Peiratinae (Hemiptera: Heteroptera: Reduviidae). Zootaxa 2010, 2476, 39–52. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Dai, W. Fine Morphology of the Mouthparts in Cheilocapsus nigrescens (Hemiptera: Heteroptera: Miridae) Reflects Adaptation for Phytophagous Habits. Insects 2019, 10, 143. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.; Dai, W. Sensory armature and stylets of the mouthparts of Stephanitis nashi (Hemiptera: Cimicomorpha: Tingidae), their morphology and function. Micron 2020, 132, 102840. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Wang, W.; Brożek, J.; Dai, W. Unique Fine Morphology of Mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) Adapted to Millipede Feeding. Insects 2020, 11, 386. [Google Scholar] [CrossRef]
- Peregrine, D. Fine structure of sensilla basiconica on the labium of the cotton stainer, Dysdercus fasciatus (Signoret) (Heteroptera: Pyrrhocoridae). Int. J. Insect Morphol. Embryol. 1972, 1, 241–251. [Google Scholar] [CrossRef]
- Schoonhoven, L.; Henstra, S. Morphology of Some Rostrum Receptors in Dysdercus spp. Neth. J. Zool. 1971, 22, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Gaffal, K. Terminal sensilla on the labium of Dysdercus intermedius distant (Heteroptera: Pyrrhocoridae). Int. J. Insect Morphol. Embryol. 1981, 10, 1–6. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. Morphology and distribution of antennal sense organs and diversity of mouthpart structures in Odontopus nigricornis (Stall) and Nezara viridula L. (Hemiptera). Int. J. Insect Morphol. Embryol. 1995, 24, 119–132. [Google Scholar] [CrossRef]
- Usha Rani, P.; Madhavendra, S.S. External morphology of antennal and rostral sensillae in four hemipteran insects and their possible role in host plant selection. Int. J. Trop. Insect. Sci. 2005, 25, 198–207. [Google Scholar] [CrossRef]
- Sinitsina, E.E.; Krutov, V.V. Antennal and labial sense organs in the bug Podizus maculiventris (Hemiptera, Pentatomidae). Zool. Zhurnal 1996, 75, 1172–1173. [Google Scholar]
- Anderson, W.G.; Heng-Moss, T.M.; Baxendale, F.P.; Baird, L.M.; Sarath, G.; Higley, L. Chinch Bug (Hemiptera: Blissidae) Mouthpart Morphology, Probing Frequencies, and Locations on Resistant and Susceptible Germplasm. J. Econ. Entomol. 2006, 99, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Baker, G.T.; Chen, X.; Ma, P.W. Labial tip sensilla of Blissus leucopterus leucopterus (Hemiptera: Blissidae): Ultrastructure and behavior. Insect Sci. 2008, 15, 271–275. [Google Scholar] [CrossRef]
- Rani, P.U. Sensillary morphology on the rostral apex and their possible role in prey location behaviour of the carnivorous stinkbug, Eocanthecona furcellata (Wolff) (Heteroptera: Pentatomidae). Acta Zool. 2009, 90, 246–253. [Google Scholar] [CrossRef]
- Parveen, S.; Ahmad, A.; Brożek, J.; Ramamurthy, V.V. Morphological diversity of the labial sensilla of phytophagous and predatory Pentatomidae (Hemiptera: Heteroptera), with reference to their possible functions. Zootaxa 2015, 4039, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Ventura, M.U.; Montalván, R.; Panizzi, A.R. Feeding preferences and related types of behaviour of Neomegalotomus parvus. Entomol. Exp. Appl. 2000, 97, 309–315. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. Fine structure of mouthparts and feeding performance of Pyrrhocoris sibiricus Kuschakevich with remarks on the specialisation of sensilla and stylets for seed feeding. PLoS ONE 2017, 12, e01772. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Dai, W. How Does the Intricate Mouthpart Apparatus Coordinate for Feeding in the Hemimetabolous Insect Pest Erthesina fullo? Insects 2020, 11, 503. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Brożek, J.; Dai, W. Morphological Disparity of the Mouthparts in Polyphagous Species of Largidae (Heteroptera: Pentatomomorpha: Pyrrhocoroidea) Reveals Feeding Specialization. Insects 2020, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Brożek, J.; Dai, W. Comparative Morphology of the Mouthparts in Three Predatory Stink Bugs (Heteroptera: Asopinae) Reveals Feeding Specialization of Stylets and Sensilla. Insects 2020, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, D.A.; Polat, I.; Gözüpek, H.; Kıyak, S.; Suludere, Z.A. Scanning electron microscope study of the sensilla on antenna and mouthparts in Eurygaster testudinaria (Geoffroy, 1785) (Hemiptera, Heteroptera, Scutelleridae). J. Het. Turk. 2021, 3, 14–30. [Google Scholar]
- Taszakowski, A.; Kaszyca-Taszakowska, N. Teratological cases of the antennae in the family Aradidae (Hemiptera: Heteroptera). Sci. Rep. 2020, 10, 1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altner, H.; Prillinger, L. Ultrastructure of Invertebrate Chemo-, Thermo-, and Hygroreceptors and Its Functional Significance. Int. Rev. Cytol. 1980, 67, 69–139. [Google Scholar] [CrossRef]
- Shields, V.D. High resolution ultrastructural investigation of insect sensory organs using field emission scanning electron microscopy. In Microscopy: Science, Technology, Applications and Education; Méndez-Vilas, A., Díaz, J., Eds.; Formatex Research Center: Badajoz, Spain, 2010; pp. 321–328. [Google Scholar]
- Keil, T.A. Functional morphology of insect mechanoreceptors. Microsc. Res. Tech. 1997, 39, 506–531. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Ultrastructure and Function of Insect Chemosensilla. Annu. Rev. Entomol. 1980, 25, 27–47. [Google Scholar] [CrossRef]
- Slifer, E.H. The Structure of Arthropod Chemoreceptors. Annu. Rev. Entomol. 1970, 15, 121–142. [Google Scholar] [CrossRef]
- Li, Z.; Wei, Y.; Sun, L.; An, X.; Dhiloo, K.H.; Wang, Q.; Xiao, Y.; Khashaveh, A.; Gu, S.; Zhang, Y. Mouthparts enriched odorant binding protein AfasOBP11 plays a role in the gustatory perception of Adelphocoris fasciaticollis. J. Insect Physiol. 2019, 117, 103915. [Google Scholar] [CrossRef]
- Weirauch, C. Cladistic analysis of Reduviidae (Heteroptera: Cimicomorpha) based on morphological characters. Syst. Entomol. 2008, 33, 229–274. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aporous Sensilla | ||||
|---|---|---|---|---|
| Type | Localisation | Shape | Cuticle Surface | Proposed Function |
| A1 | SI, SIII | cupola | – | mechanoreception |
| A2 | SI, SII, SIV | hair-like | grooved | mechanoreception |
| A3 | SII, SIV | cone-like | smooth | mechanoreception (proprioreception) |
| A4 | SII | cone-like | grooved | mechanoreception |
| A5 | SI, SII, SIII | hair-like | smooth | mechanoreception |
| A6 | SII, SIII, SIV | cone-like | smooth | mechanoreception |
| A7 | SIV (lt) | shallow depression | – | mechanoreception |
| A8 | SIV (lt) | hair-like | finely grooved | mechanoreception |
| A9 | SIV (lt) | hair-like | smooth | mechanoreception |
| Porous Sensilla | ||||
| Type | Localisation | Shape | Cuticle | Proposed Function |
| P1 | SIV (lt) | cone-like | smooth | chemoreception (gustation) |
| P2 | SIV (lt) | cone-like | smooth | chemoreception (gustation) |
| P3 | SIV (lt) | cupola | – | chemoreception |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taszakowski, A.; Masłowski, A.; Brożek, J. Labial Sensory Organs of Two Leptoglossus Species (Hemiptera: Coreidae): Their Morphology and Supposed Function. Insects 2023, 14, 30. https://doi.org/10.3390/insects14010030
Taszakowski A, Masłowski A, Brożek J. Labial Sensory Organs of Two Leptoglossus Species (Hemiptera: Coreidae): Their Morphology and Supposed Function. Insects. 2023; 14(1):30. https://doi.org/10.3390/insects14010030
Chicago/Turabian StyleTaszakowski, Artur, Adrian Masłowski, and Jolanta Brożek. 2023. "Labial Sensory Organs of Two Leptoglossus Species (Hemiptera: Coreidae): Their Morphology and Supposed Function" Insects 14, no. 1: 30. https://doi.org/10.3390/insects14010030
APA StyleTaszakowski, A., Masłowski, A., & Brożek, J. (2023). Labial Sensory Organs of Two Leptoglossus Species (Hemiptera: Coreidae): Their Morphology and Supposed Function. Insects, 14(1), 30. https://doi.org/10.3390/insects14010030