
Spatial Distribution of Pyrethroid Resistance and kdr Mutations in Aedes aegypti from La Guajira, Colombia


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
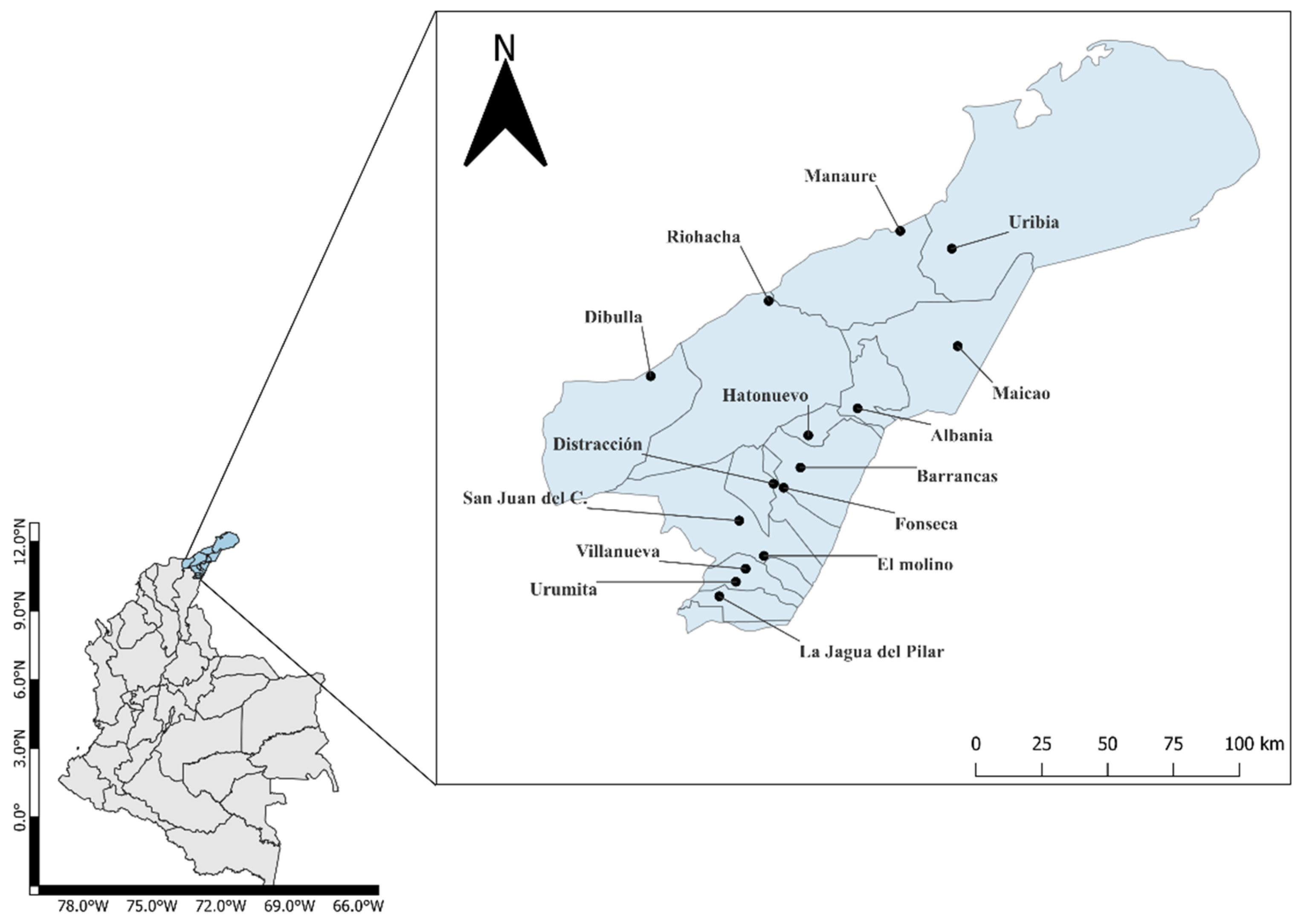
2.1. Study Area
2.2. Collection of Mosquitoes and Obtaining F1/F2
2.3. Bioassays
2.4. Identification of kdr Mutations
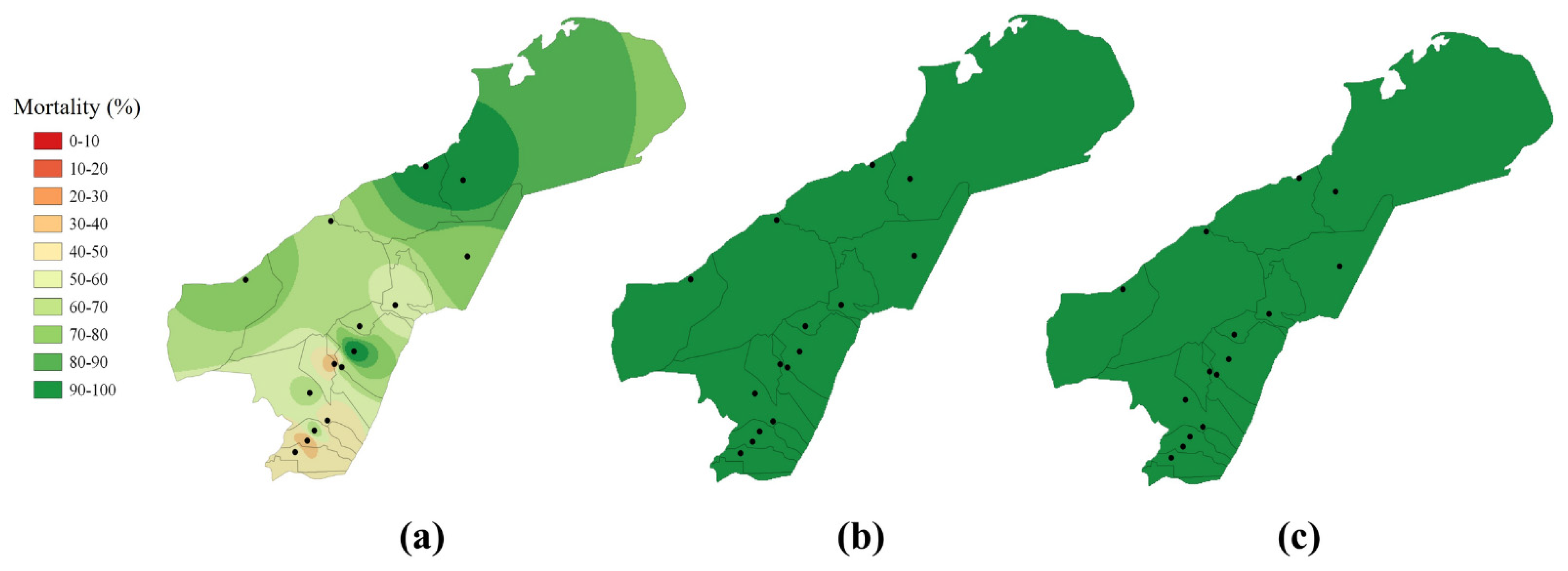
2.5. Spatial Distribution of Resistance to Pyrethroids
3. Results
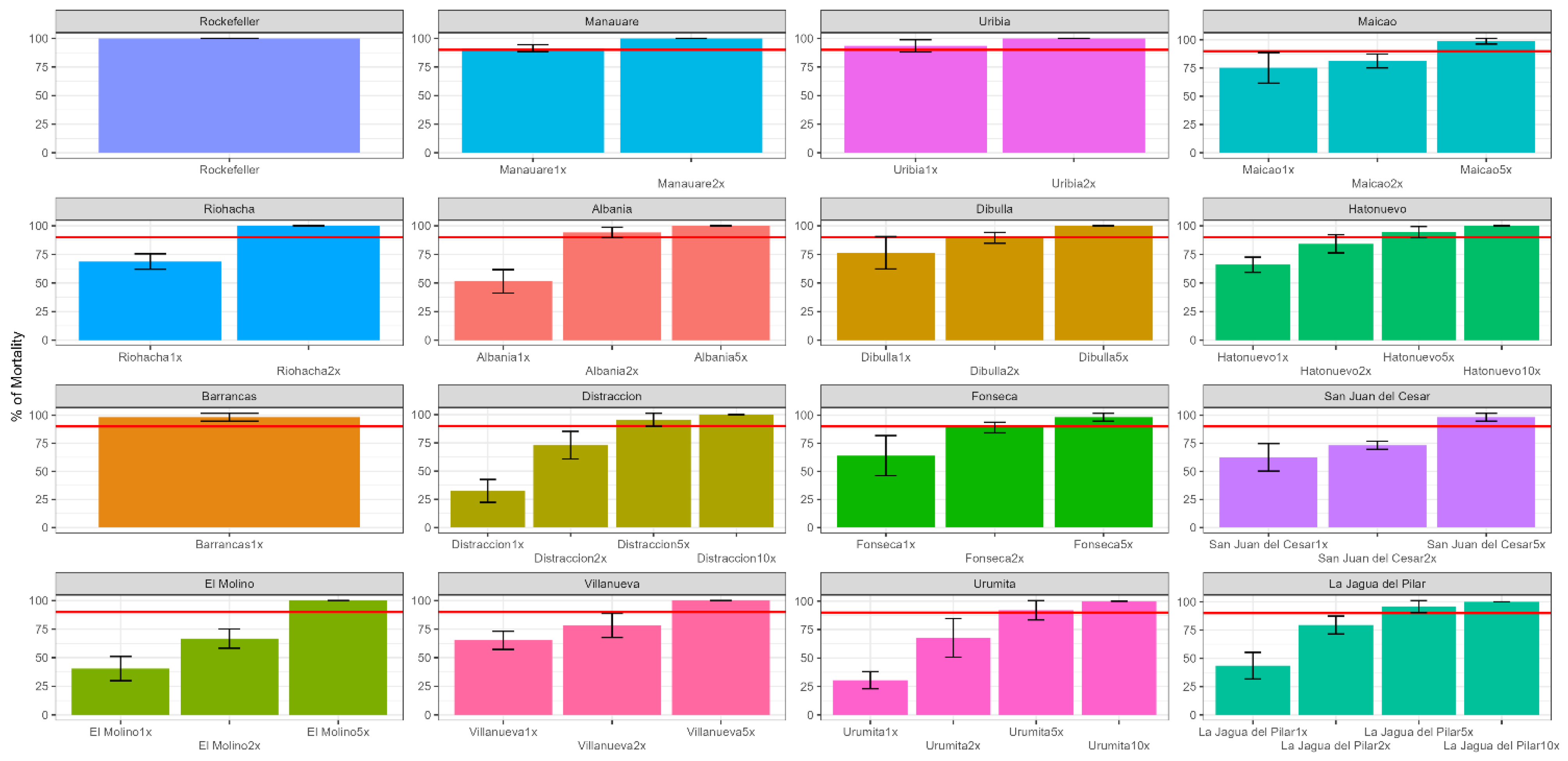
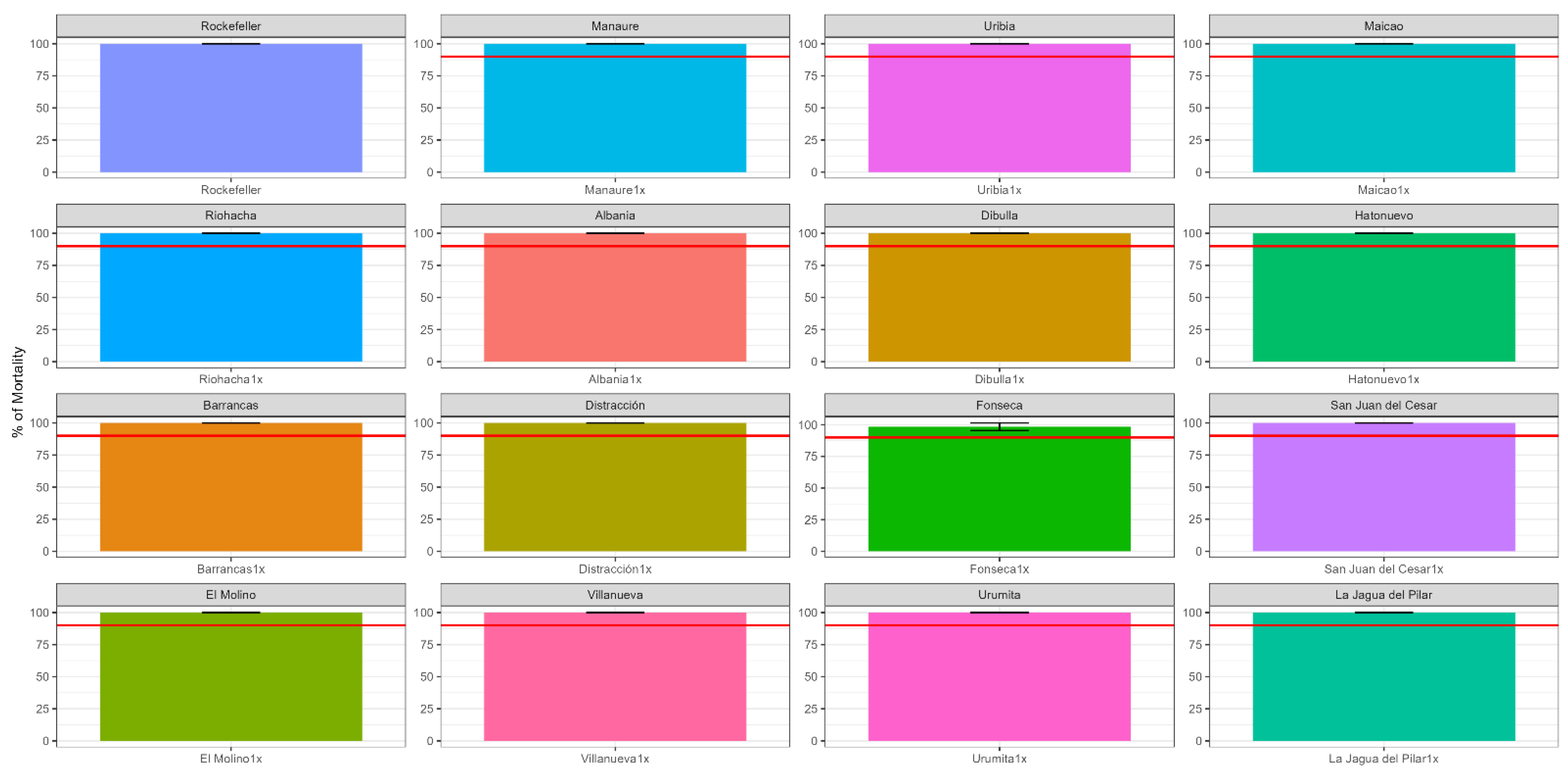
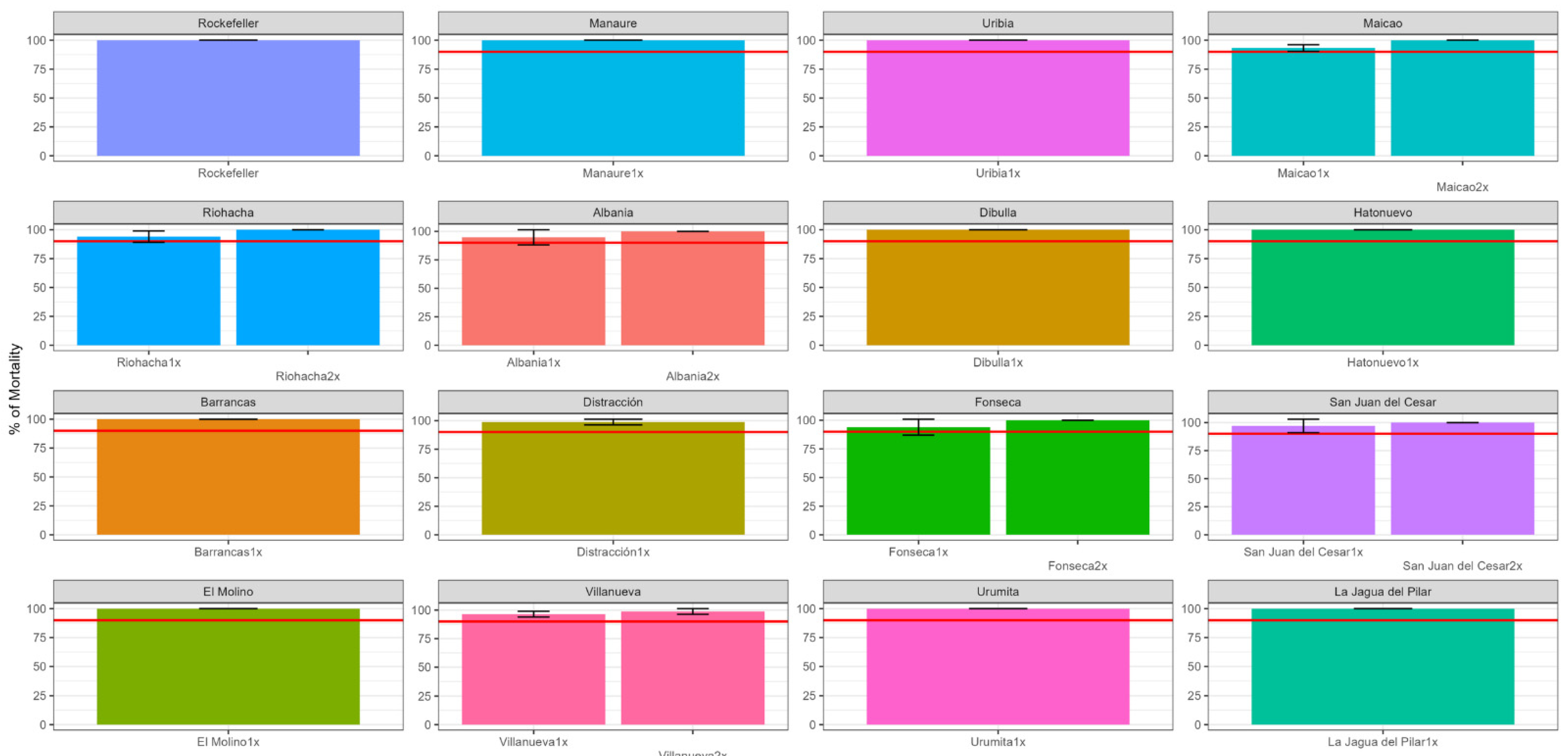
3.1. Frequency and Intensity of Pyrethroid Resistance
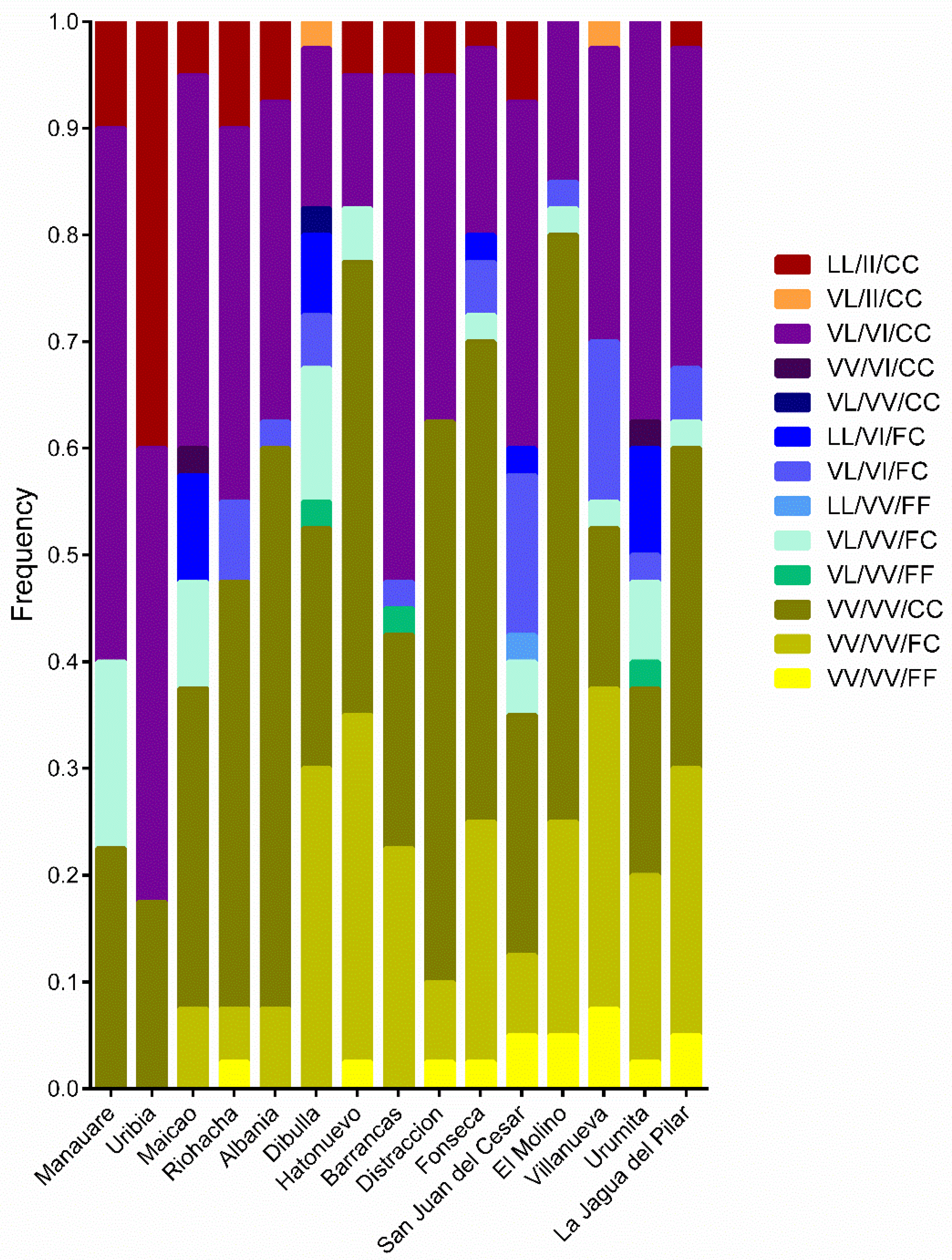
3.2. Allele and Genotype Frequencies of the kdr Mutations V410L, V1016I, and F1534C
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arredondo-García, J.L.; Méndez-Herrera, A.; Medina-Cortina, H. Arbovirus en Latinoamérica. Acta Pediatr. Mex. 2016, 37, 111–131. Available online: https://www.medigraphic.com/pdfs/actpedmex/apm-2016/apm162h.pdf (accessed on 25 July 2022). [CrossRef] [Green Version]
- Instituto Nacional de Salud (INS). Gestión para la Vigilancia Entomológica y Control de la Transmisión de Dengue. 2010. Available online: https://www.paho.org/col/dmdocuments/Entomologia_DENGUE.pdf (accessed on 25 July 2022).
- Organización Panamericana de la Salud (OPS). Actualización Epidemiológica Dengue y otras Arbovirosis. 2020. Available online: https://www.paho.org/es/documentos/actualizacion-epidemiologica-dengue-otras-arbovirosis-10-junio-2020 (accessed on 25 July 2022).
- Organización Panamericana de la Salud (OPS). Actualización Epidemiológica. 2021. Available online: https://www.paho.org/es/documentos/actualizacion-epidemiologica-arbovirosis-contexto-covid-19-2-julio-2021 (accessed on 25 July 2022).
- Organización Panamericana de la Salud (OPS). Actualización Epidemiológica Anual para Dengue, Chikungunya y Zika. 2022. Available online: https://ais.paho.org/ha_viz/arbo/pdf/OPS%20Arbo%20Boletin%202022.pdf (accessed on 18 October 2022).
- Instituto Nacional de Salud (INS). Informe del Evento Dengue, Colombia, Hasta el Décimo Tercer Periodo Epidemiológico. 2015. Available online: https://www.ins.gov.co/buscador-eventos/Informesdeevento/Dengue%202015.pdf (accessed on 23 July 2022).
- Instituto Nacional de Salud (INS). Informe del Evento Dengue, Colombia, Hasta el Décimo Tercer Periodo Epidemiológico. 2016. Available online: https://www.ins.gov.co/buscador-eventos/Informesdeevento/Dengue%202016.pdf (accessed on 23 July 2022).
- Instituto Nacional de Salud (INS). Informe del Evento Dengue, Colombia. 2017. Available online: https://www.ins.gov.co/buscador-eventos/Informesdeevento/DENGUE%202017.pdf (accessed on 23 July 2022).
- Instituto Nacional de Salud (INS). Informe del Evento Dengue, Colombia. 2018. Available online: https://www.ins.gov.co/buscador-eventos/Informesdeevento/DENGUE_2018.pdf (accessed on 23 July 2022).
- Instituto Nacional de Salud (INS). Informe del Evento Dengue, Colombia. 2019. Available online: https://www.ins.gov.co/buscador-eventos/Informesdeevento/DENGUE_2019.pdf (accessed on 23 July 2022).
- Secretaria de Salud de la Guajira, A. (Secretaria de Salud de la Guajira, Riohacha, La Guajira, Colombia); Incidencia Dengue para el Departamento de La Guajira (Colombia). 2020; (Unpublished information). [Google Scholar]
- Informe Quincenal Epidemiológico Nacional (IQEN). Chikungunya en Colombia, el Inicio de la Transmisión Autóctona. 2014; Volume 19, pp. 261–279. Available online: https://www.ins.gov.co/buscador-eventos/IQEN/IQEN%20vol%2019%202014%20num%2018.pdf (accessed on 20 July 2022).
- Instituto Nacional de Salud. Informe de Evento, Enfermedad por virus Zika y Chikungunya. 2019. Available online: https://www.ins.gov.co/buscador-eventos/Informesdeevento/chikungunya%20y%20enfermedad%20por%20virus%20Zika_2019.pdf (accessed on 23 July 2022).
- World Health Organization (WHO). Pesticides and Their Application for the Control of Vectors and Pests of Public Health Importance. 2006. Available online: https://apps.who.int/iris/bitstream/handle/10665/69223/WHO_CDS_NTD_WHOPES_GCDPP_2006.1_eng.pdf?sequence=1&isAllowed=y (accessed on 22 July 2022).
- Maestre-Serrano, R. Susceptibility status of Aedes aegypti to insecticides in Colombia. In Insecticides-Pest Engineering; Perveeb, F., Ed.; InTech Publishers: Rijeka, Croatia, 2012; pp. 163–200. [Google Scholar] [CrossRef]
- Maestre, R.; Gómez, D. Dengue: Epidemiología, políticas públicas y resistencia de vectores a insecticidas. Rev. Cienc. Biomed. 2013, 4, 302–317. [Google Scholar]
- Secretaria de Salud de la Guajira, B. (Secretaria de Salud de la Guajira, Riohacha, La Guajira, Colombia); Insecticidas usados en el departamento de La Guajira para el control del dengue. 2022; (Unpublished information). [Google Scholar]
- Du, Y.; Nomura, Y.; Satar, G.; Hu, Z.; Nauen, R.; He, S.Y.; Zhorov, B.S.; Dong, K. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc. Natl. Acad. Sci. USA 2013, 110, 11785–11790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterall, W.A. Voltage-gated sodium channels at 60: Structure, function and pathophysiology. J. Physiol. 2012, 590, 2577–2589. [Google Scholar] [CrossRef] [PubMed]
- Maestre, R.; Rey, G.; De Las Salas, J.; Vergara, C.; Santacoloma, L.; Goenaga, S.; Carrasquilla, M.C. Susceptibility status of Aedes aegypti to insecticides in Atlántico (Colombia). Rev. Colomb. Entomol. 2010, 36, 242–248. [Google Scholar] [CrossRef]
- Maestre-Serrano, R.; Gomez-Camargo, D.; Ponce-Garcia, G.; Flores, A.E. Susceptibility to insecticides and resistance mechanisms in Aedes aegypti from the Colombian Caribbean Region. Pestic. Biochem. Physiol. 2014, 116, 63–73. [Google Scholar] [CrossRef]
- Atencia, M.C.; Pérez, M.D.J.; Jaramillo, M.C.; Caldera, S.M.; Cochero, S.; Bejarano, E.E. Primer reporte de la mutación F1534C asociada con resistencia cruzada a DDT y piretroides en Aedes aegypti en Colombia. Biomedica 2016, 36, 432–437. [Google Scholar] [CrossRef] [Green Version]
- Granada, Y.; Mejía-Jaramillo, A.M.; Strode, C.; Triana-Chavez, O.A. Point Mutation V419L in the sodium channel gene from natural populations of Aedes aegypti is involved in resistance to λ-cyhalothrin in Colombia. Insects 2018, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Maestre-Serrano, R.; Pareja-Loaiza, P.; Gomez Camargo, D.; Ponce-García, G.; Flores, A.E. Co-occurrence of V1016I and F1534C mutations in the voltage-gated sodium channel and resistance to pyrethroids in Aedes aegypti (L.) from the Colombian Caribbean region. Pest Manag. Sci. 2019, 75, 1681–1688. [Google Scholar] [CrossRef] [Green Version]
- Pareja-Loaiza, P.X.; Santacoloma Varon, L.; Rey Vega, G.; Gómez-Camargo, D.; Maestre-Serrano, R.; Lenhart, A. Mechanisms associated with pyrethroid resistance in populations of Aedes aegypti (Diptera: Culicidae) from the Caribbean coast of Colombia. PLoS ONE 2020, 15, e0228695. [Google Scholar] [CrossRef] [PubMed]
- Brogdon, W.; Chan, A. Guideline for Evaluating Insecticide Resistance in Vectors Using the CDC Bottle Bioassay (with Inserts 1 in 2012 and 2 in 2014). 2010. Available online: http://www.cdc.gov/malaria/features/bioassay.html (accessed on 15 January 2022).
- Brogdon, W.G.; McAllister, J.C. Simplification of adult mosquito bioassays through use of time-mortality determinations in glass bottles. J. Am. Mosq. Control Assoc. 1998, 14, 159–164. [Google Scholar] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes, 2nd ed.; World Health Organization, 2016; Available online: https://apps.who.int/iris/handle/10665/250677 (accessed on 12 January 2022).
- Saavedra-Rodriguez, K.; Urdaneta-Marquez, L.; Rajatileka, S.; Moulton, M.; Flores, A.E.; Fernandez-Salas, I.; Bisset, J.; Rodriguez, M.; McCall, P.J.; Donnelly, M.J.; et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 2007, 16, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Somwang, P.; Prapanthadara, L.A. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop. Med. Int. Health 2011, 16, 501–509. [Google Scholar] [CrossRef]
- Haddi, K.; Tomé, H.V.V.; Du, Y.; Valbon, W.R.; Nomura, Y.; Martins, G.F.; Dong, K.; Oliveira, E.E. Detection of a new pyrethroid resistance mutation (V410L) in the sodium channel of Aedes aegypti: A potential challenge for mosquito control. Sci. Rep. 2017, 7, 46549. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.H.; Hung, M.C. Comparison of Spatial Interpolation Techniques Using Visualization and Quantitative Assessment. In Applications of Spatial Statistics; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. Systems of mating. II. The effects of inbreeding on the genetic composition of a population. Genetics 1921, 6, 124–143. [Google Scholar] [CrossRef]
- Agresti, A.; Coull, B.A. Approximate is better than “exact” for interval estimation of binominal proportions. Am. Stat. 1998, 52, 119–126. [Google Scholar]
- Santacoloma Varón, L.; Chaves Córdoba, B.; Brochero, H.L. Susceptibilidad de Aedes aegypti a DDT, deltametrina y lambdacialotrina en Colombia. Rev. Panam. Salud Pública 2010, 27, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-González, I.; Quiñones, M.L.; Lenhart, A.; Brogdon, W.G. Insecticide resistance status of Aedes aegypti (L.) from Colombia. Pest Manag. Sci. 2011, 67, 430–437. [Google Scholar] [CrossRef]
- Ocampo, C.B.; Salazar-Terreros, M.J.; Mina, N.J.; McAllister, J.; Brogdon, W. Insecticide resistance status of Aedes aegypti in 10 localities in Colombia. Acta Trop. 2011, 118, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ardila-Roldán, S.; Santacoloma, L.; Brochero, H. Estado de la sensibilidad a los insecticidas de uso en salud pública en poblaciones naturales de Aedes aegypti (Diptera: Culicidae) del departamento de Casanare, Colombia. Biomedica 2013, 33, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Padilla, J.C.; Lizarazo, F.E.; Murillo, O.L.; Mendigaña, F.A.; Pachón, E.; Vera, M.J. Epidemiología de las principales enfermedades transmitidas por vectores en Colombia, 1990–2016. Biomedica 2017, 37, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guajira 360°. Available online: https://guajira360.org/informe-economico-reactivacion-del-sector-agropecuario-de-la-guajira/ (accessed on 22 December 2022).
- Wagner Medina, M. Las Huellas Ambientales del oro Blanco: La Expansión Algodonera en el Valle del río Cesar. (1950–1980). Master’s Dissertation/Tesis, Universidad de los Andes, Colombia, July 2011. Available online: https://repositorio.uniandes.edu.co/bitstream/handle/1992/11452/u470720.pdf?sequence=1&isAllowed=y (accessed on 21 December 2022).
- Lu, G.Y.; Wong, D.W. An adaptive inverse-distance weighting spatial interpolation technique. Comput. Geosci. 2008, 34, 1044–1055. [Google Scholar] [CrossRef]
- Chang, C.; Shen, W.K.; Wang, T.T.; Lin, Y.H.; Hsu, E.L.; Dai, S.M. A novel amino acid substitution in a voltage-gated sodium channel is associated with knockdown resistance to permethrin in Aedes aegypti. Insect Biochem. Mol. Biol. 2009, 39, 272–278. [Google Scholar] [CrossRef]
- Srisawat, R.; Komalamisra, N.; Apiwathnasorn, C.; Paeporn, P.; Roytrakul, S.; Rongsriyam, Y.; Eshita, Y. Field-collected permethrin-resistant Aedes aegypti from central Thailand contain point mutations in the domain IIS6 of the sodium channel gene (KDR). Southeast Asian J. Trop. Med. Public Health 2012, 43, 1380–1386. [Google Scholar]
- Kushwah, R.B.; Dykes, C.L.; Kapoor, N.; Adak, T.; Singh, O.P. Pyrethroid-resistance and presence of two knockdown resistance (kdr) mutations, F1534C and a novel mutation T1520I, in Indian Aedes aegypti. PLoS Negl. Trop. Dis. 2015, 9, e3332. [Google Scholar] [CrossRef] [Green Version]
- Brengues, C.; Hawkes, N.J.; Chandre, F.; McCarroll, L.; Duchon, S.; Guillet, P.; Manguin, S.; Morgan, J.C.; Hemingway, J. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 2003, 17, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Collet, M.L.; Frizzo, C.; Orlandin, E.; Rona, L.D.P.; Nascimento, J.C.; Montano, M.A.E.; Müller, G.A.; Wagner, G. Frequency of the Val1016Ile mutation on the kdr gene in Aedes aegypti (Diptera: Culicidae) in south Brazil. GMR 2016, 15, gmr15048940. [Google Scholar] [CrossRef]
- Siller, Q.; Ponce, G.; Lozano, S.; Flores, A.E. Update on the frequency of Ile1016 mutation in voltage-gated sodium channel gene of Aedes aegypti in Mexico. J. Am. Mosq. Control Assoc. 2011, 27, 357–362. [Google Scholar] [CrossRef]
- Vera-Maloof, F.Z.; Saavedra-Rodriguez, K.; Elizondo-Quiroga, A.E.; Lozano-Fuentes, S.; Black, W.C., IV. Coevolution of the Ile1,016 and Cys1,534 Mutations in the Voltage Gated Sodium Channel Gene of Aedes aegypti in Mexico. PLoS Negl. Trop. Dis. 2015, 9, e0004263. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Segura, O.K.; Ontiveros-Zapata, K.A.; Lopez-Monroy, B.; Ponce-Garcia, G.; Gutierrez-Rodriguez, S.M.; Davila-Barboza, J.A.; Mora-Jasso, E.J.; Flores, A.E. Distribution and frequency of the kdr mutation V410L in natural populations of Aedes aegypti (L.) (Diptera: Culicidae) from Eastern and Southern Mexico. J. Med. Entomol. 2020, 57, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Obando, O.A.; Pietrobon, A.J.; Dalla Bona, A.C.; Navarro-Silva, M.A. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in Aedes aegypti populations from Jacarezinho (Brazil) after a dengue outbreak. Rev. Bras. Entomol. 2016, 60, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Melo Costa, M.; Campos, K.B.; Brito, L.P.; Roux, E.; Melo Rodovalho, C.; Bellinato, D.F.; Lima, J.B.P.; Martins, A.J. Kdr genotyping in Aedes aegypti from Brazil on a nation-wide scale from 2017 to 2018. Sci. Rep. 2020, 10, 13267. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.; Palomino, M.; Mendoza-Uribe, L.; Sinti, C.; Liebman, K.A.; Lenhart, A. Susceptibility to insecticides and resistance mechanisms in three populations of Aedes aegypti from Peru. Parasit. Vectors 2019, 12, 494. [Google Scholar] [CrossRef] [Green Version]
- Zardkoohi, A.; Castañeda, D.; Lol, J.C.; Castillo, C.; Lopez, F.; Marín Rodriguez, R.; Padilla, N. Co-occurrence of kdr mutations V1016I and F1534C and its association with phenotypic resistance to pyrethroids in Aedes aegypti (Diptera: Culicidae) populations from Costa Rica. J. Med. Entomol. 2020, 57, 830–836. [Google Scholar] [CrossRef]
- Ponce-García, G.; Del Río-Galvan, S.; Barrera, R.; Saavedra-Rodriguez, K.; Villanueva-Segura, K.; Felix, G.; Amador, M.; Flores, A.E. Knockdown Resistance Mutations in Aedes aegypti (Diptera: Culicidae) From Puerto Rico. J. Med. Entomol. 2016, 53, 1410–1414. [Google Scholar] [CrossRef]
- Alvarez, L.C.; Ponce-García, G.; Saavedra-Rodriguez, K.; Lopez, B.; Flores, A.E. Frequency of V1016I and F1534C mutations in the voltage-gated sodium channel gene in Aedes aegypti in Venezuela. Pest Manag. Sci. 2015, 71, 863–869. [Google Scholar] [CrossRef]
- Aguirre-Obando, O.A.; Dalla Bona, A.C.; Duque, L.J.E.; Navarro-Silva, M.A. Insecticide resistance and genetic variability in natural populations of Aedes (Stegomyia) aegypti (Diptera: Culicidae) from Colombia. Zoologia 2015, 32, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Aponte, A.; Penilla, R.P.; Rodríguez, A.D.; Ocampo, C.B. Mechanisms of pyrethroid resistance in Aedes (Stegomyia) aegypti from Colombia. Acta Trop. 2019, 191, 146–154. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | n | VV | VL | LL | Frequency (IC 95%) | FIS | x2 Hardy-Weinberg | p Value |
|---|---|---|---|---|---|---|---|---|
| Manaure | 40 | 9 | 27 | 4 | 0.44 (0.30–0.59) | −0.37 | 5.51 | 0.02 |
| Uribia | 40 | 7 | 17 | 16 | 0.61 (0.46–0.75) | 0.10 | 0.43 | 0.50 |
| Maicao | 40 | 16 | 18 | 6 | 0.38 (0.24–0.53) | 0.04 | 0.06 | 0.80 |
| Riohacha | 40 | 19 | 17 | 4 | 0.31 (0.20–0.47) | 0.01 | 0 | 0.94 |
| Albania | 40 | 24 | 13 | 3 | 0.24 (0.13–0.39) | 0.10 | 0.42 | 0.51 |
| Dibulla | 40 | 20 | 17 | 3 | 0.29 (0.17–0.44) | −0.03 | 0.05 | 0.81 |
| Hatonuevo | 40 | 31 | 7 | 2 | 0.14 (0.06–0.28) | 0.26 | 2.74 | 0.10 |
| Barrancas | 40 | 17 | 21 | 2 | 0.31 (0.20–0.47) | −0.22 | 1.96 | 0.16 |
| Distracción | 40 | 24 | 13 | 3 | 0.24 (0.13–0.39) | 0.10 | 0.42 | 0.51 |
| Fonseca | 40 | 28 | 10 | 2 | 0.18 (0.09–0.32) | 0.13 | 0.72 | 0.40 |
| San Juan del Cesar | 40 | 14 | 21 | 5 | 0.39 (0.25–0.54) | −0.10 | 0.44 | 0.50 |
| El Molino | 40 | 32 | 8 | 0 | 0.10 (0.04–0.24) | −0.11 | 0.49 | 0.50 |
| Villanueva | 40 | 21 | 19 | 0 | 0.24 (0.13–0.39) | −0.31 | 3.88 | 0.05 |
| Urumita | 40 | 16 | 20 | 4 | 0.35 (0.22–0.50) | −0.09 | 0.39 | 0.53 |
| La Jagua del Pilar | 40 | 24 | 15 | 1 | 0.21 (0.11–0.36) | −0.12 | 0.58 | 0.44 |
| Population | n | VV | VI | II | Frequency (IC 95%) | FIS | x2 Hardy-Weinberg | p Value |
|---|---|---|---|---|---|---|---|---|
| Manaure | 40 | 16 | 20 | 4 | 0.35 (0.22–0.50) | −0.1 | 0.39 | 0.53 |
| Uribia | 40 | 7 | 17 | 16 | 0.61 (0.46–0.75) | 0.1 | 0.44 | 0.51 |
| Maicao | 40 | 19 | 19 | 2 | 0.29 (0.17–0.44) | −0.16 | 1.02 | 0.31 |
| Riohacha | 40 | 19 | 17 | 4 | 0.31 (0.19–0.47) | 0.01 | 0 | 0.94 |
| Albania | 40 | 24 | 13 | 3 | 0.24 (0.13–0.39) | 0.1 | 0.42 | 0.52 |
| Dibulla | 40 | 28 | 11 | 1 | 0.16 (0.08–0.31) | 0 | 0 | 1 |
| Hatonuevo | 40 | 33 | 5 | 2 | 0.11 (0.05–0.25) | 0.37 | 5.59 | 0.01 |
| Barrancas | 40 | 18 | 20 | 2 | 0.30 (0.18–0.45) | −0.19 | 1.45 | 0.23 |
| Distracción | 40 | 25 | 13 | 2 | 0.21 (0.11–0.36) | 0.02 | 0.03 | 0.86 |
| Fonseca | 40 | 29 | 10 | 1 | 0.15 (0.07–0.29) | 0.02 | 0.02 | 0.9 |
| San Juan del Cesar | 40 | 17 | 20 | 3 | 0.33 (0.20–0.48) | −0.14 | 0.78 | 0.38 |
| El Molino | 40 | 33 | 7 | 0 | 0.09 (0.03–0.21) | −0.096 | 0.36 | 0.55 |
| Villanueva | 40 | 22 | 17 | 1 | 0.24 (0.13–0.39) | −0.17 | 1.2 | 0.27 |
| Urumita | 40 | 19 | 21 | 0 | 0.26 (0.15–0.42) | −0.35 | 5.06 | 0.02 |
| La Jagua del Pilar | 40 | 25 | 14 | 1 | 0.20 (0.10–0.35) | −0.09 | 0.35 | 0.55 |
| Population | n | FF | FC | CC | Frequency (IC95%) | FIS | x2 Hardy-Weinberg | p Value |
|---|---|---|---|---|---|---|---|---|
| Manaure | 40 | 0 | 7 | 33 | 0.91 (0.79–0.97) | −0.1 | 0.37 | 0.54 |
| Uribia | 40 | 0 | 0 | 40 | 1.00 (0.91–1.00) | - | - | - |
| Maicao | 40 | 0 | 11 | 29 | 0.86 (0.72–0.94) | −0.16 | 1.02 | 0.31 |
| Riohacha | 40 | 1 | 5 | 34 | 0.91 (0.79–0.97) | 0.22 | 2.05 | 0.15 |
| Albania | 40 | 0 | 4 | 36 | 0.95 (0.84–0.99) | −0.05 | 0.11 | 0.74 |
| Dibulla | 40 | 1 | 22 | 17 | 0.70 (0.55–0.82) | −0.37 | 5.4 | 0.02 |
| Hatonuevo | 40 | 1 | 15 | 24 | 0.79 (0.64–0.89) | −0.12 | 0.58 | 0.44 |
| Barrancas | 40 | 1 | 10 | 29 | 0.85 (0.71–0.93) | 0.02 | 0.02 | 0.9 |
| Distracción | 40 | 1 | 3 | 36 | 0.94 (0.82–0.98) | 0.36 | 5.18 | 0.02 |
| Fonseca | 40 | 1 | 13 | 26 | 0.81 (0.67–0.90) | −0.07 | 0.18 | 0.67 |
| San Juan del Cesar | 40 | 3 | 12 | 25 | 0.78 (0.63–0.88) | 0.14 | 0.78 | 0.38 |
| El Molino | 40 | 2 | 10 | 28 | 0.83 (0.69–0.91) | 0.13 | 0.72 | 0.40 |
| Villanueva | 40 | 3 | 19 | 18 | 0.69 (0.53–0.81) | −0.11 | 0.44 | 0.5 |
| Urumita | 40 | 2 | 15 | 23 | 0.76 (0.61–0.87) | −0.03 | 0.05 | 0.82 |
| La Jagua del Pilar | 40 | 2 | 13 | 25 | 0.79 (0.64–0.89) | 0.02 | 0.03 | 0.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maestre-Serrano, R.; Flórez-Rivadeneira, Z.; Castro-Camacho, J.M.; Soto-Arenilla, E.; Gómez-Camargo, D.; Pareja-Loaiza, P.; Ponce-Garcia, G.; Juache-Villagrana, A.E.; Flores, A.E. Spatial Distribution of Pyrethroid Resistance and kdr Mutations in Aedes aegypti from La Guajira, Colombia. Insects 2023, 14, 31. https://doi.org/10.3390/insects14010031
Maestre-Serrano R, Flórez-Rivadeneira Z, Castro-Camacho JM, Soto-Arenilla E, Gómez-Camargo D, Pareja-Loaiza P, Ponce-Garcia G, Juache-Villagrana AE, Flores AE. Spatial Distribution of Pyrethroid Resistance and kdr Mutations in Aedes aegypti from La Guajira, Colombia. Insects. 2023; 14(1):31. https://doi.org/10.3390/insects14010031
Chicago/Turabian StyleMaestre-Serrano, Ronald, Zulibeth Flórez-Rivadeneira, Juan M. Castro-Camacho, Eva Soto-Arenilla, Doris Gómez-Camargo, Paula Pareja-Loaiza, Gustavo Ponce-Garcia, Alan E. Juache-Villagrana, and Adriana E. Flores. 2023. "Spatial Distribution of Pyrethroid Resistance and kdr Mutations in Aedes aegypti from La Guajira, Colombia" Insects 14, no. 1: 31. https://doi.org/10.3390/insects14010031
APA StyleMaestre-Serrano, R., Flórez-Rivadeneira, Z., Castro-Camacho, J. M., Soto-Arenilla, E., Gómez-Camargo, D., Pareja-Loaiza, P., Ponce-Garcia, G., Juache-Villagrana, A. E., & Flores, A. E. (2023). Spatial Distribution of Pyrethroid Resistance and kdr Mutations in Aedes aegypti from La Guajira, Colombia. Insects, 14(1), 31. https://doi.org/10.3390/insects14010031