


On the Origin of Neo-Sex Chromosomes in the Neotropical Dragonflies Rhionaeschna bonariensis and R. planaltica (Aeshnidae, Odonata)
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Chromosome Preparations
2.3. Fluorescent Banding
2.4. FISH with rDNA Probes
2.5. Genomic in Situ Hybridization (GISH)
2.6. Microscopy and Image Processing
3. Results
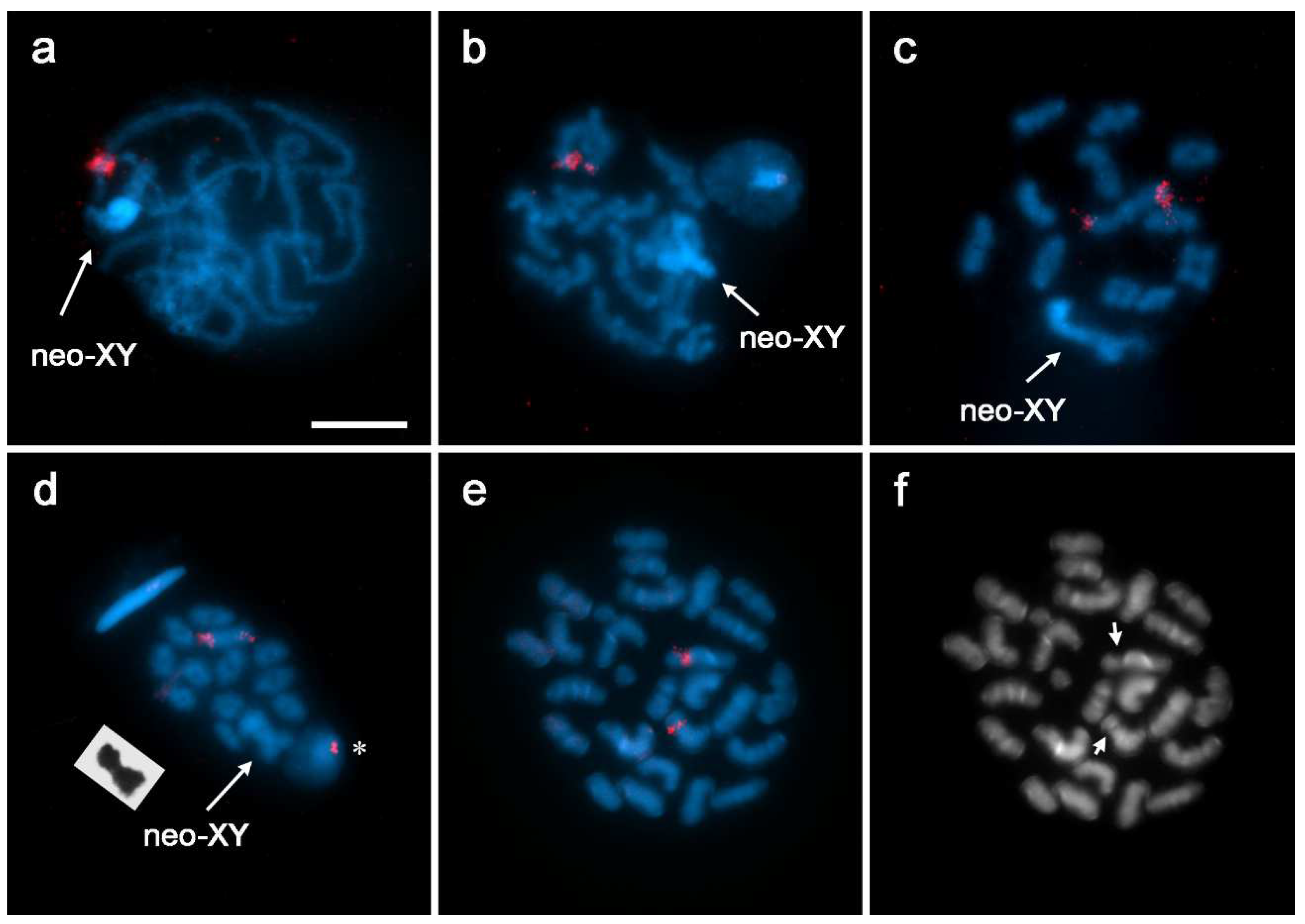
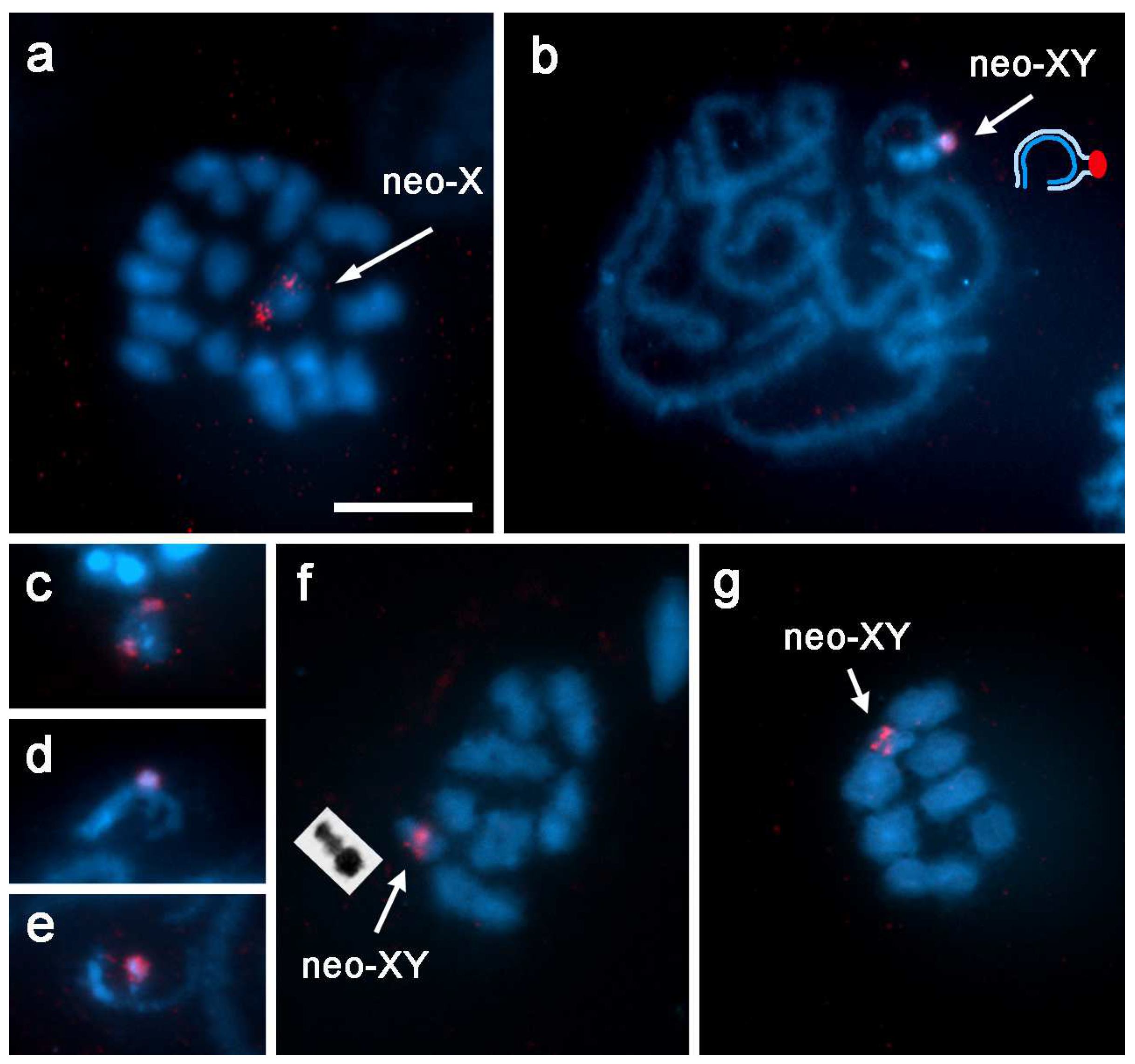
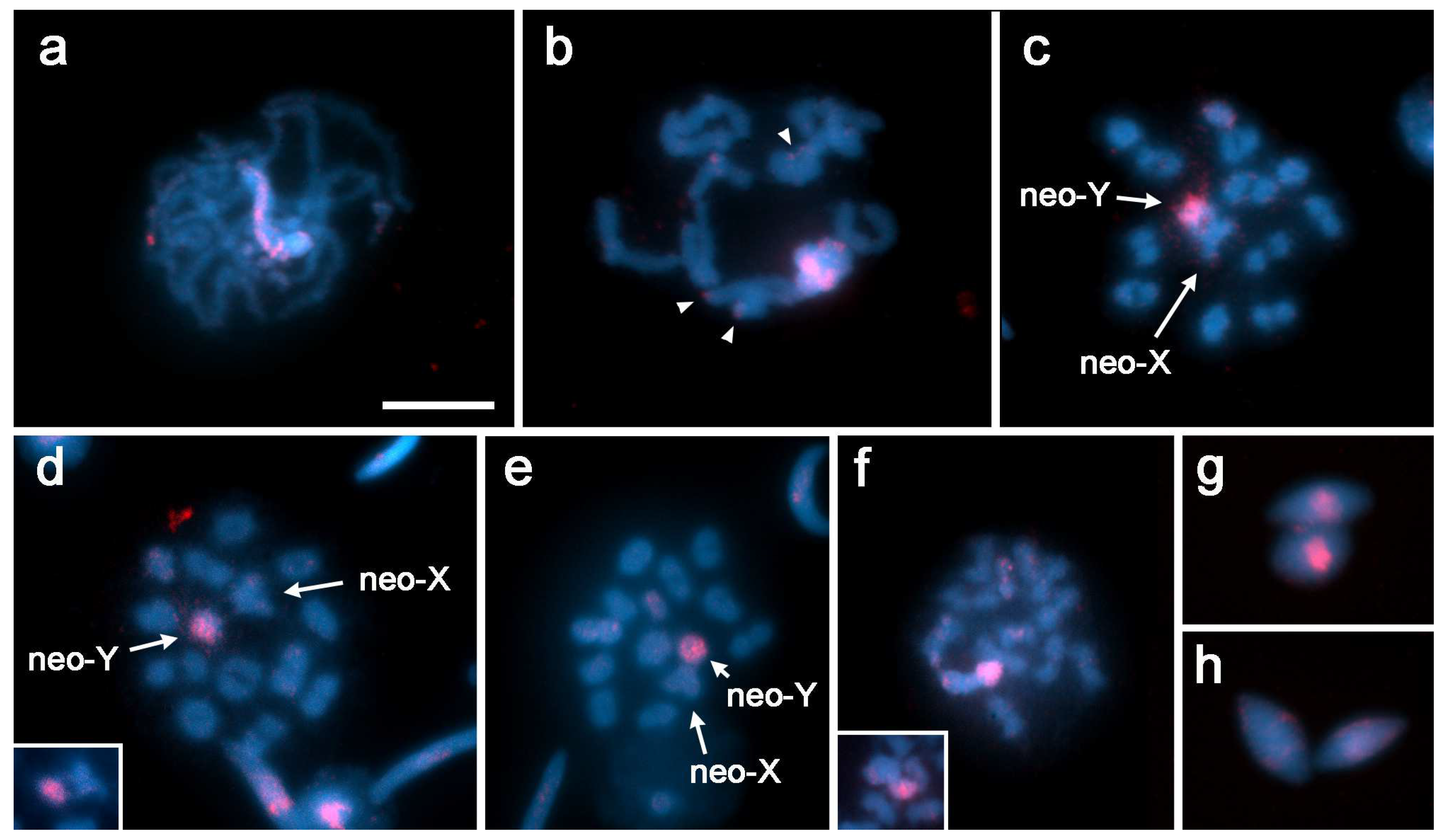
3.1. Localization of rDNA
3.2. GISH Characterization of Neo-XY Sex Chromosomes in Rhionaeschna Bonariensis
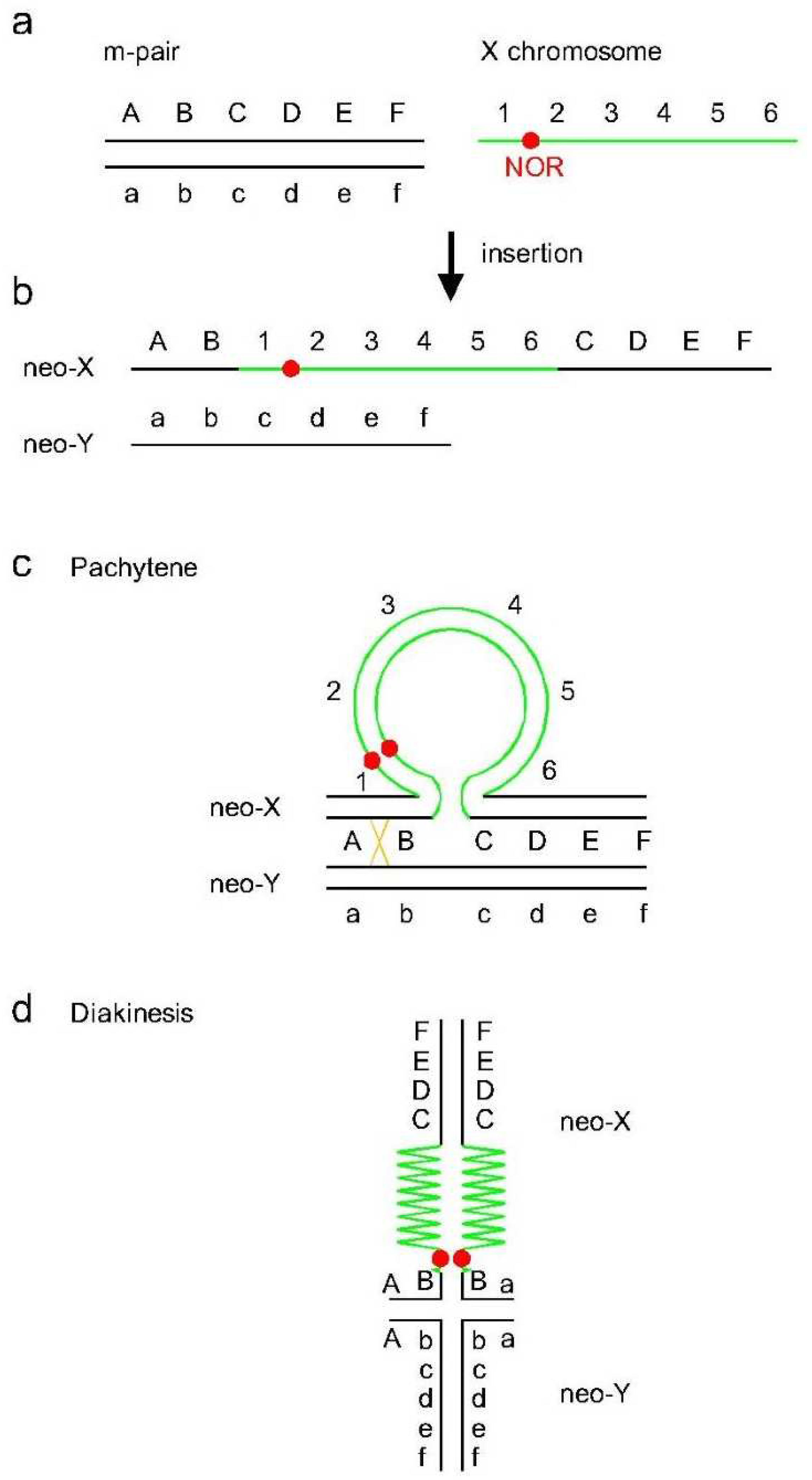
4. Discussion
4.1. Origin of the Neo-XY Sex Chromosomes in Rhionaeschna planaltica
4.2. Characteristics of the Neo-XY Sex Chromosomes in Rhionaeschna bonariensis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tennessen, K.J. The rate of species descriptions in Odonata. Entomol. News 1997, 108, 122–126. [Google Scholar]
- Paulson, D.; Schorr, M.; Deliry, C. World Odonata List. Available online: https://www2.pugetsound.edu/academics/academic-resources/slater-museum/biodiversity-resources/dragonflies/world-odonata-list2/ (accessed on 28 October 2022).
- Pessacq, P.; Muzón, J.; Neiss, U.G. Order Odonata. In Thorp and Covich’s Freshwater Invertebrates; Thorp, J.H., Covich, A.P., Eds.; Elsevier: London, UK, 2018; pp. 355–366. [Google Scholar]
- Lohmann, H. Das Phylogenetische System der Anisoptera (Odonata). Entomol. Z. 1996, 106, 209–266. [Google Scholar]
- Carle, F.L.; Kjer, K.M.; May, M.L. A molecular phylogeny and classification of Anisoptera (Odonata). Arthropod Syst. Phylogeny 2015, 73, 281–301. [Google Scholar]
- Oertli, B. The use of dragonflies in the assessment and monitoring of aquatic habitats. In Dragonflies and Damselflies: Model Organisms for Ecological and Evolutionary Research; Córdoba-Aguilar, A., Ed.; Oxford University Press: Oxford, UK, 2008; pp. 79–95. [Google Scholar]
- Lozano, F.; del Palacio, A.; Ramos, L.S.; Granato, L.; Drozd, A.; Muzón, J. Recovery of local dragonfly diversity following restoration of an artificial lake in an urban area near Buenos Aires. Basic Appl. Ecol. 2022, 58, 88–97. [Google Scholar] [CrossRef]
- Nokkala, S.; Laukkanen, A.; Nokkala, C. Mitotic and meiotic chromosomes in Somatochlora metallica (Corduliidae, Odonata). The absence of localized centromeres and inverted meiosis. Hereditas 2002, 136, 7–12. [Google Scholar] [CrossRef]
- Oksala, T. Zytologische Studien an Odonaten I. Chromosomenverhältnisse bei der Gattung Aeschna mit besonderer Berücksichtigung der Postreduktionellen Teilung der Bivalente; Annales Academiae Scientiarum Fennicae (A) IV Biologica; Suomalainen Tiedeakatemia: Helsinki, Finland, 1943; Volume IV, pp. 1–64. [Google Scholar]
- Kiauta, B. Sex chromosomes and sex determining mechanisms in Odonata, with a review of the cytological conditions in the family Gomphidae, and references to the karyotypic evolution in the order. Genetica 1969, 40, 127–157. [Google Scholar] [CrossRef]
- Tyagi, B.K. Cytotaxonomy of the genus Onychogomphus Selys (Odonata: Anisoptera, Gomphidae), with a special reference to the evolution of the sex-determining mechanism and reduced chromosome number in the family Gomphidae. In Proceedings of the First Indian Symposium of Odonatology; Mathavan, S., Ed.; Madurai Kamaraj University: Madurai, India, 1985; pp. 217–226. [Google Scholar]
- Mola, L.M. Post-reductional meiosis in Aeshna (Aeshnidae, Odonata). Hereditas 1995, 122, 47–55. [Google Scholar] [CrossRef]
- Mola, L.M. Meiotic studies in nine species of Erythrodiplax (Libellulidae, Odonata). Neo-XY sex chromosome system in E. media. Cytologia 1996, 61, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Mola, L. Cytogenetics of American Odonata. In Odonata: Biology of Dragonflies; Tyagi, B.K., Ed.; Scientific Publishers: Jodhpur, India, 2007; pp. 153–173. [Google Scholar]
- Mola, L.M.; Papeschi, A.G.; Taboada Carrillo, E. Cytogenetics of seven species of dragonflies. A novel sex chromosome determining system in Micrathyria ungulata. Hereditas 1999, 131, 147–153. [Google Scholar] [CrossRef]
- Mola, L.M.; Fourastié, M.F.; Agopian, S.S. High karyotypic variation in Orthemis Hagen, 1861 species, with insights about the neo-XY in Orthemis ambinigra Calvert, 1909 (Libellulidae, Odonata). Comp. Cytogenet. 2021, 15, 355–375. [Google Scholar] [CrossRef]
- Perepelov, E.A.; Bugrov, A.G.; Warchałowska-Śliwa, E. C-banded karyotypes of some dragonfly species from Russia. Folia Biol. 1998, 46, 137–142. [Google Scholar]
- Perepelov, E.; Bugrov, A.G. Constitutive heterochromatin in chromosomes of some Aeshnidae, with notes on the formation of the neo-XY/neo-XX mode of sex determination in Aeshna (Anisoptera). Odonatologica 2002, 31, 77–83. [Google Scholar]
- Mola, L.M.; Papeschi, A.G. Holokinetic chromosomes at a glance. BAG—J. Basic Appl. Genet. 2006, 17, 147–153. [Google Scholar]
- Papeschi, A.G.; Bressa, M.J. Evolutionary cytogenetics in Heteroptera. J. Biol. Res. 2006, 5, 3–21. [Google Scholar]
- Sahara, K.; Yoshido, A.; Traut, W. Sex chromosome evolution in moths and butterflies. Chromosome Res. 2012, 20, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Gaba, K. First report on a multiple sex chromosome system (X1X2X30) and population variations in the frequency of ring bivalents in Pyrrhocoridae (Hemiptera: Heteroptera). Eur. J. Entomol. 2015, 112, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Šíchová, J.; Ohno, M.; Dincă, V.; Watanabe, M.; Sahara, K.; Marec, F. Fissions, fusions, and translocations shaped the karyotype and multiple sex chromosome constitution of the northeast-Asian wood white butterfly, Leptidea amurensis. Biol. J. Linn. Soc. 2016, 118, 457–471. [Google Scholar] [CrossRef]
- Yoshido, A.; Šíchová, J.; Pospíšilová, K.; Nguyen, P.; Voleníková, A.; Šafář, J.; Provazník, J.; Vila, R.; Marec, F. Evolution of multiple sex-chromosomes associated with dynamic genome reshuffling in Leptidea wood-white butterflies. Heredity 2020, 125, 138–154. [Google Scholar] [CrossRef]
- Hejníčková, M.; Dalíková, M.; Potocký, P.; Tammaru, T.; Trehubenko, M.; Kubíčková, S.; Marec, F.; Zrzavá, M. Degenerated, undifferentiated, rearranged, lost: High variability of sex chromosomes in Geometridae (Lepidoptera) identified by sex chromatin. Cells 2021, 10, 2230. [Google Scholar] [CrossRef]
- Seshachar, B.R.; Bagga, S. Chromosome number and sex-determining mechanism in the dragonfly Hemianax ephippiger (Burmeister). Cytologia 1962, 27, 443–449. [Google Scholar] [CrossRef] [Green Version]
- Kiauta, B. Studies on the germ cell chromosome cytology of some cytotaxonomically interesting or hitherto not studied Odonata from the autonomous region Friuli-Venezia Giulia (Northern Italy). Att. Mus. Civ. Stor. Nat. Trieste 1971, 27, 65–127. [Google Scholar]
- Ferreira, A.; Kiauta, B.; Zaha, A. Male germ cell chromosomes of thirty-two Brazilian dragonflies. Odonatologica 1979, 8, 5–22. [Google Scholar]
- Kiauta, B. The status of the Japanese Crocothemis servilia (Drury) as revealed by karyotypic morphology (Anisoptera: Libellulidae). Odonatologica 1983, 12, 381–388. [Google Scholar]
- Agopian, S.; Mola, L.M. Intra and interspecific karyotype variability in five species of Libellulidae (Anisoptera, Odonata). Caryologia 1988, 41, 69–78. [Google Scholar] [CrossRef]
- Kiauta, B. Variation in size of the dragonfly m-chromosome, with considerations on its significance for the chorogeography and taxonomy of the order Odonata, and notes on the validity of the rule of Reinig. Genetica 1968, 39, 64–74. [Google Scholar] [CrossRef]
- Kuznetsova, V.G.; Golub, N.V. A Checklist of chromosome numbers and a review of karyotype variation in Odonata of the world. Comp. Cytogenet. 2020, 14, 501–540. [Google Scholar] [CrossRef]
- von Ellenrieder, N. A synopsis of the Neotropical species of “Aeshna” Fabricius: The genus Rhionaeschna Förster (Odonata: Aeshnidae). Tijdschr. Entomol. 2003, 146, 67–207. [Google Scholar] [CrossRef] [Green Version]
- Cumming, R.B. Cytogenetic Studies in the Order Odonata. Ph.D. Thesis, University of Texas, Austin, TX, USA, 1964. [Google Scholar]
- Kiauta, B. Notes on new or little known dragonfly karyotypes. III. Spermatocyte chromosomes of four Nearctic anisopterans: Aeshna californica Calvert (Aeshnidae), Cordulia shurtleffi Scudder (Corduliidae), Sympetrum internum Montgomery, and S. madidum (Hagen) (Libellulidae). Genen Phaenen 1973, 16, 7–12. [Google Scholar]
- Mola, L.M.; Papeschi, A.G. Karyotype evolution in Aeshna (Aeshnidae, Odonata). Hereditas 1994, 121, 185–189. [Google Scholar] [CrossRef]
- Cabral-de-Mello, D.C.; Marec, F. Universal fluorescence in situ hybridization (FISH) protocol for mapping repetitive DNAs in insects and other arthropods. Mol. Genet. Genom. 2021, 296, 513–526. [Google Scholar] [CrossRef]
- Kuznetsova, V.G.; Maryańska-Nadachowska, A.; Shapoval, N.A.; Anokhin, B.A.; Shapoval, A.P. Cytogenetic characterization of eight Odonata species originating from the Curonian Spit (the Baltic Sea, Russia) using C-banding and FISH with 18S rDNA and telomeric (TTAGG)n probes. Cytogenet. Genome Res. 2018, 153, 147–157. [Google Scholar] [CrossRef]
- Kuznetsova, V.; Maryańska-Nadachowska, N.; Anokhin, B.; Shapoval, N.; Shapoval, A. Chromosomal analysis of eight species of dragonflies (Anisoptera) and damselflies (Zygoptera) using conventional cytogenetics and fluorescence in situ hybridization: Insights into the karyotype evolution of the ancient insect order Odonata. J. Zool. Syst. Evol. Res. 2021, 59, 387–399. [Google Scholar] [CrossRef]
- Traut, W. Pachytene mapping in the female silkworm, Bombyx mori L. (Lepidoptera). Chromosoma 1976, 58, 275–284. [Google Scholar] [CrossRef]
- Rebagliati, P.J.; Papeschi, A.G.; Mola, L.M. Meiosis and fluorescent banding in Edessa meditabunda and E. rufomarginata (Heteroptera: Pentatomidae: Edessinae). Eur. J. Entomol. 2003, 100, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Whiting, M.F. Phylogeny of the holometabolous insect orders: Molecular evidence. Zool. Scr. 2002, 31, 3–15. [Google Scholar] [CrossRef]
- Fuková, I.; Nguyen, P.; Marec, F. Codling moth cytogenetics: Karyotype, chromosomal location of rDNA, and molecular differentiation of sex chromosomes. Genome 2005, 48, 1083–1092. [Google Scholar] [CrossRef]
- Sahara, K.; Marec, F.; Traut, W. TTAGG telomeric repeats in chromosomes of some insects and other arthropods. Chromosome Res. 1999, 7, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Adilardi, R.S.; Affilastro, A.A.O.; Martí, D.A.; Mola, L.M. Cytogenetic analysis on geographically distant parthenogenetic populations of Tityus trivittatus Kraepelin, 1898 (Scorpiones, Buthidae): Karyotype, constitutive heterochromatin and rDNA localization. Comp. Cytogenet. 2014, 8, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Camacho, J.P.M.; Cabrero, J.; López-León, M.D.; Cabral-de-Mello, D.C.; Ruiz-Ruano, F.J. Grasshoppers (Orthoptera). In Protocols for Cytogenetic Mapping of Arthropod Genomes; Sharakhov, I.V., Ed.; CRC Press: Boca Ratón, FL, USA, 2015; Volume 1, pp. 416–418. [Google Scholar]
- Traut, W.; Sahara, K.; Otto, T.D.; Marec, F. Molecular differentiation of sex chromosomes probed by comparative genomic hybridization. Chromosoma 1999, 108, 173–180. [Google Scholar] [CrossRef]
- Kiauta, B. Notes on new or little known dragonfly karyotypes, 2. Male germ cell chromosomes of four east Mediterranean species: Lestes barbarus (Fabricius), Calopteryx splendens amasina Bartenev (Zygoptera: Lestidae, Calopterygidae), Caliaeschna microstigma (Schneider) and Orthetrum taeniolatum (Schneider) (Anisoptera: Aeshnidae, Libellulidae). Genen Phaenen 1972, 15, 95–98. [Google Scholar]
- Kiauta, B.; Kiauta, M. The chromosome numbers of sixteen dragonfly species from the Arun Valley, Eastern Nepal. Not. Odonatol. 1982, 9, 143–146. [Google Scholar]
- Criniti, A.; Simonazzi, G.; Cassanelli, S.; Ferrari, M.; Bizzaro, D.; Manicardi, G.C. Distribution of heterochromatin and rDNA on the holocentric chromosomes of the aphids Dysaphis plantaginea and Melanaphis pyraria (Hemiptera: Aphididae). Eur. J. Entomol. 2009, 106, 153–157. [Google Scholar] [CrossRef] [Green Version]
- de Gennaro, D.; Rebagliati, P.J.; Mola, L.M. Fluorescent banding and meiotic behaviour in Erythrodiplax nigricans (Libellulidae) and Coryphaeschna perrensi (Aeschnidae) (Anisoptera, Odonata). Caryologia 2008, 61, 60–67. [Google Scholar] [CrossRef]
- Cattani, M.V.; Papeschi, A.G. Nucleolus organizing regions and semi-persistent nucleolus during meiosis in Spartocera fusca (Thunberg) (Coreidae, Heteroptera). Hereditas 2004, 140, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.P.M.; Belda, J.; Cabrero, J. Meiotic behaviour of the holocentric chromosomes of Nezara viridula (Insecta, Heteroptera) analysed by C-banding and silver impregnation. Can. J. Genet. Cytol. 1985, 27, 490–497. [Google Scholar] [CrossRef]
- Papeschi, A.G.; Mola, L.M.; Bressa, M.J.; Greizerstein, E.J.; Lía, V.; Poggio, L. Behaviour of ring bivalents in holokinetic systems: Alternative sites of spindle attachment in Pachylis argentinus and Nezara viridula (Heteroptera). Chromosome Res. 2003, 11, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Castillo, E.R.; Martí, D.A.; Bidau, C.J. Sex and neo-sex chromosomes in Orthoptera: A review. J. Orthoptera Res. 2010, 19, 213–231. [Google Scholar] [CrossRef] [Green Version]
- Castillo, E.R.D.; Taffarel, A.; Martí, D.A. The early evolutionary history of neo-sex chromosomes in Neotropical grasshoppers, Boliviacris noroestensis (Orthoptera: Acrididae: Melanoplinae). Eur. J. Entomol. 2014, 111, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Bidau, C.J.; Martí, D.A.; Castillo, E.R. Inexorable spread: Inexorable death? The fate of neo-XY chromosomes of grasshoppers. J. Genet. 2011, 90, 397–400. [Google Scholar] [CrossRef]
- Palacios-Gimenez, O.M.; Milani, D.; Lemos, B.; Castillo, E.R.; Martí, D.A.; Ramos, E.; Martins, C.; Cabral-De-Mello, D.C. Uncovering the evolutionary history of neo-XY sex chromosomes in the grasshopper Ronderosia bergii (Orthoptera, Melanoplinae) through satellite DNA analysis. BMC Evol. Biol. 2018, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Steinemann, M.; Steinemann, S. Enigma of Y chromosome degeneration: Neo-Y and neo-X chromosomes of Drosophila miranda a model for sex chromosome evolution. Genetica 1998, 102–103, 409–420. [Google Scholar] [CrossRef]
- Vítková, M.; Fuková, I.; Kubíčková, S.; Marec, F. Molecular divergence of the W chromosomes in pyralid moths (Lepidoptera). Chromosome Res. 2007, 15, 917–930. [Google Scholar] [CrossRef]
- Palacios-Gimenez, O.M.; Marti, D.A.; Cabral-de-Mello, D.C. Neo-sex chromosomes of Ronderosia bergi: Insight into the evolution of sex chromosomes in grasshoppers. Chromosoma 2015, 124, 353–365. [Google Scholar] [CrossRef]
- Jetybayev, I.Y.; Bugrov, A.G.; Ünal, M.; Buleu, O.G.; Rubtsov, N.B. Molecular cytogenetic analysis reveals the existence of two independent neo-XY sex chromosome systems in Anatolian Pamphagidae grasshoppers. BMC Evol. Biol. 2017, 17, 20. [Google Scholar] [CrossRef] [Green Version]
- Buleu, O.; Jetybayev, I.; Mofdi-Neyestanak, M.; Bugrov, A. Karyotypes diversity in some Iranian Pamphagidae grasshoppers (Orthoptera, Acridoidea, Pamphagidae): New insights on the evolution of the neo-XY sex chromosomes. Comp. Cytogenet. 2020, 14, 549–566. [Google Scholar] [CrossRef]
- Kaiser, V.B.; Bachtrog, D. Evolution of sex chromosomes in insects. Annu. Rev. Genet. 2010, 44, 91–112. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Ellison, C.E.; Kaiser, V.B.; Alekseyenko, A.A.; Gorchakov, A.A.; Bachtrog, D. The epigenome of evolving Drosophila neo-sex chromosomes: Dosage compensation and heterochromatin formation. PLoS Biol. 2013, 11, e1001711. [Google Scholar] [CrossRef] [Green Version]
- Palacios-Gimenez, O.M.; Dias, G.B.; de Lima, L.G.; Kuhn, G.C.E.S.; Ramos, É.; Martins, C.; Cabral-De-Mello, D.C. High-throughput analysis of the satellitome revealed enormous diversity of satellite DNAs in the neo-Y chromosome of the cricket Eneoptera surinamensis. Sci. Rep. 2017, 7, 6422. [Google Scholar] [CrossRef] [Green Version]
- Moses, M.J.; Poorman, P.A. Synapsis, synaptic adjustment and DNA synthesis in mouse oocytes. In Chromosomes Today; Bennett, M.D., Gropp, A., Wolf, U., Eds.; Springer: Dordrecht, The Netherlands, 1984; pp. 90–103. [Google Scholar] [CrossRef]
- Weith, A.; Traut, W. Synaptic adjustment, non-homologous pairing, and non-pairing of homologous segments in sex chromosome mutants of Ephestia kuehniella (Insecta, Lepidoptera). Chromosoma 1986, 94, 125–131. [Google Scholar] [CrossRef]
- Marec, F.; Traut, W. Sex chromosome pairing and sex chromatin bodies in W-Z translocation strains of Ephestia kuehniella (Lepidoptera). Genome 1994, 37, 426–435. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
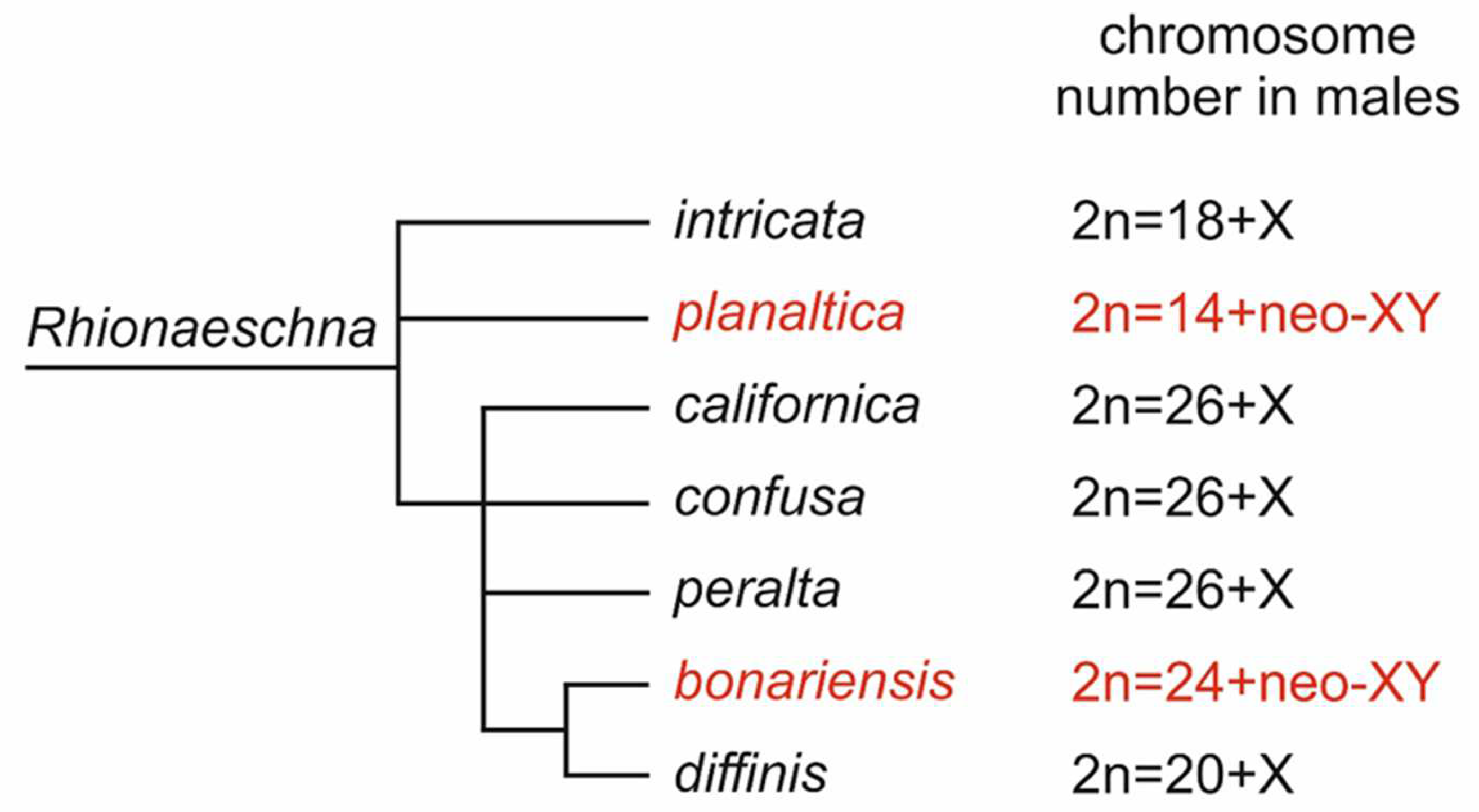
| Species | 2n | n | Locality | References |
|---|---|---|---|---|
| R. bonariensis (Rambur, 1842) | 26 | 12 + neo-XY | Argentina, Uruguay | [12] |
| R. californica (Calvert, 1895) | 27 | 13 + X0 | Canada | [35] |
| R. confusa (Rambur, 1842) | 27 | 13 + X0 | Argentina, Uruguay | [12] |
| R. diffinis (Rambur, 1842) | 21 | 10 + X0 | Bolivia | [34] |
| R. intricata (Martin, 1908) | 19 | 9 + X0 | Bolivia | [34] |
| R. peralta (Ris, 1918) | 27 | 13 + X0 | Bolivia | [34] |
| R. planaltica (Calvert, 1845) | 16 | 7 + neo-XY | Argentina | [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mola, L.M.; Vrbová, I.; Tosto, D.S.; Zrzavá, M.; Marec, F. On the Origin of Neo-Sex Chromosomes in the Neotropical Dragonflies Rhionaeschna bonariensis and R. planaltica (Aeshnidae, Odonata). Insects 2022, 13, 1159. https://doi.org/10.3390/insects13121159
Mola LM, Vrbová I, Tosto DS, Zrzavá M, Marec F. On the Origin of Neo-Sex Chromosomes in the Neotropical Dragonflies Rhionaeschna bonariensis and R. planaltica (Aeshnidae, Odonata). Insects. 2022; 13(12):1159. https://doi.org/10.3390/insects13121159
Chicago/Turabian StyleMola, Liliana M., Iva Vrbová, Daniela S. Tosto, Magda Zrzavá, and František Marec. 2022. "On the Origin of Neo-Sex Chromosomes in the Neotropical Dragonflies Rhionaeschna bonariensis and R. planaltica (Aeshnidae, Odonata)" Insects 13, no. 12: 1159. https://doi.org/10.3390/insects13121159
APA StyleMola, L. M., Vrbová, I., Tosto, D. S., Zrzavá, M., & Marec, F. (2022). On the Origin of Neo-Sex Chromosomes in the Neotropical Dragonflies Rhionaeschna bonariensis and R. planaltica (Aeshnidae, Odonata). Insects, 13(12), 1159. https://doi.org/10.3390/insects13121159