

Rickettsia Infection Benefits Its Whitefly Hosts by Manipulating Their Nutrition and Defense

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants
2.2. Insects
2.3. Entomopathogenic Fungus and Insecticides
2.4. Effect of Rickettsia on Development and Reproduction of Bemisia tabaci
2.5. Effect of Rickettsia on the Nutritional Changes of Bemisia Tabaci
2.6. Effect of Rickettsia on the Defense of Bemisia tabaci against Akanthomyces attenuatus
2.7. Effect of Rickettsia on the Defense of Bemisia Tabaci against the Parasitoid
2.8. Effect of Rickettsia on the Resistance of Bemisia tabaci to Insecticides
2.9. Statistical Analyses
3. Results
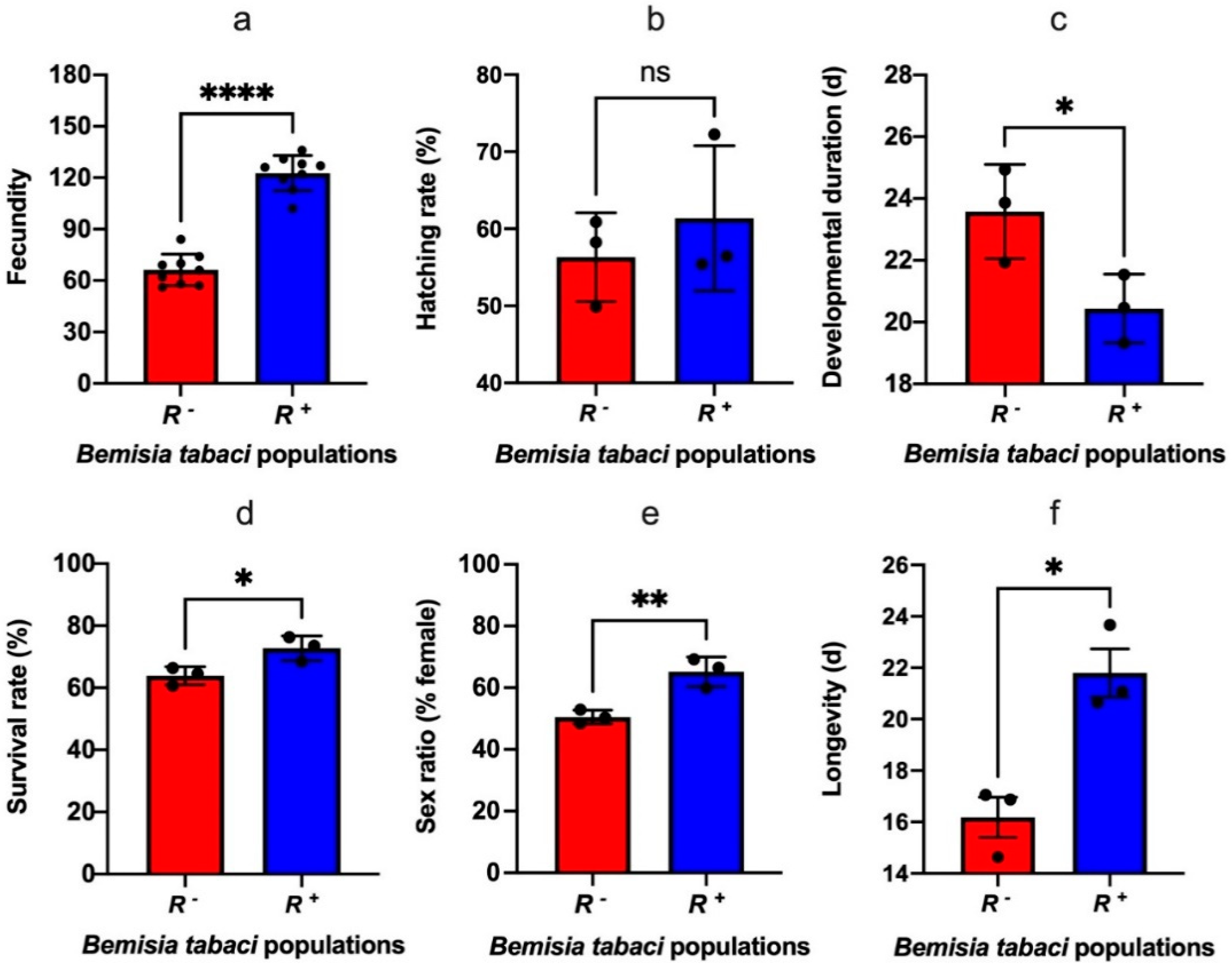
3.1. Effect of Rickettsia on Development and Reproduction of Bemisia tabaci
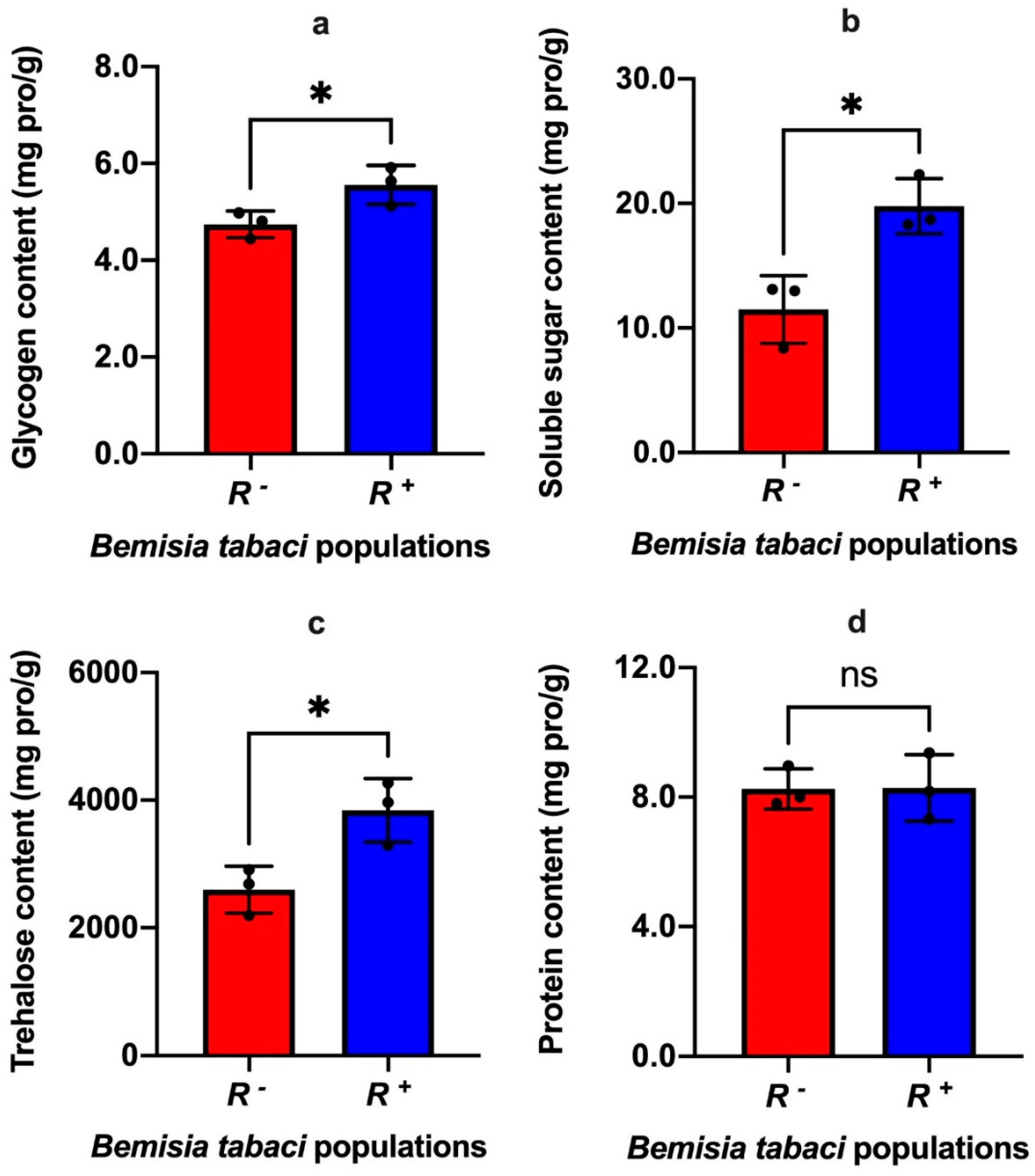
3.2. Effect of Rickettsia on the Nutritional Components of Bemisia tabaci
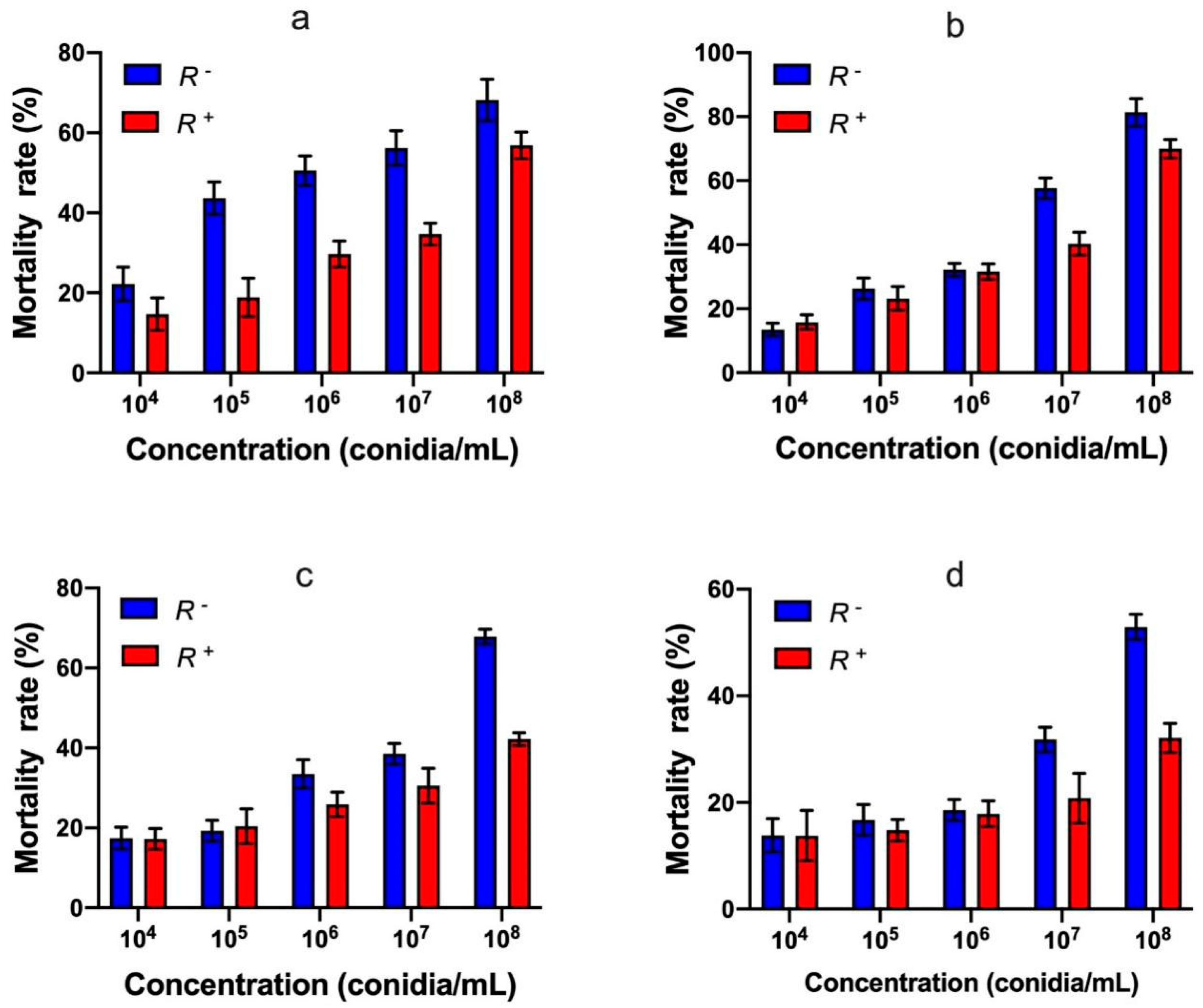
3.3. Effect of Rickettsia Persistence on Bemisia tabaci Defense against Akanthomyces attenuatus
3.4. Effect of Rickettsia Infection on Parasitism Rate of Encarsia formosa
3.5. Effect of Rickettsia Infection on Insecticide Resistance of Bemisia tabaci
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Gündüz, E.A.; Douglas, A.E. Symbiotic bacteria enable insect to use a nutritionally inadequate diet. Proc. R. Soc. B 2009, 276, 987–991. [Google Scholar] [CrossRef] [Green Version]
- Enders, L.S.; Miller, N.J. Stress-induced changes in abundance differ among obligate and facultative endosymbionts of the soybean aphid. Ecol. Evol. 2016, 6, 818–829. [Google Scholar] [CrossRef]
- Zhang, B.; Leonard, S.P.; Li, Y.; Moran, N.A. Obligate bacterial endosymbionts limit thermal tolerance of insect host species. Proc. Natl. Acad. Sci. USA 2019, 116, 24712–24718. [Google Scholar] [CrossRef]
- Lv, N.; Peng, J.; Chen, X.-Y.; Guo, C.-F.; Sang, W.; Wang, X.-M.; Ahmed, M.Z.; Xu, Y.-Y.; Qiu, B.-L. Antagonistic interaction between male-killing and cytoplasmic incompatibility induced by Cardinium and Wolbachia in the whitefly, Bemisia tabaci. Insect Sci. 2021, 28, 330–346. [Google Scholar] [CrossRef]
- Hornett, E.A.; Engelstädter, J.; Hurst, G.D.D. Hidden cytoplasmic incompatibility alters the dynamics of male-killer/host interactions. J. Evol. Biol. 2010, 23, 479–487. [Google Scholar] [CrossRef]
- Cass, B.N.; Himler, A.G.; Bondy, E.C.; Bergen, J.E.; Fung, S.K.; Kelly, S.E.; Hunter, M.S. Conditional fitness benefits of the Rickettsia bacterial symbiont in an insect pest. Oecologia 2016, 180, 169–179. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Driscoll, T.P.; Verhoeve, V.I.; Rahman, M.S.; Macaluso, K.R.; Azad, A.F. A tangled web: Origins of reproductive parasitism. Genome Biol. Evol. 2018, 10, 2292–2309. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Pan, L.-L.; Liu, S.-S. Further insight into reproductive incompatibility between putative cryptic species of the Bemisia tabaci whitefly complex. Insect Sci. 2016, 23, 215–224. [Google Scholar] [CrossRef]
- Khatun, M.F.; Jahan, S.M.H.; Lee, S.; Lee, K.Y. Genetic diversity and geographic distribution of the Bemisia tabaci species complex in Bangladesh. Acta Trop. 2018, 187, 28–36. [Google Scholar] [CrossRef]
- Shi, X.B.; Chen, G.; Pan, H.P.; Xie, W.; Wu, Q.J.; Wang, S.L.; Liu, Y.; Zhou, X.G.; Zhang, Y.J. Plants pre-infested with viruliferous MED/Q cryptic species promotes subsequent Bemisia tabaci infestation. Front. Microbiol. 2018, 9, 1404. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.; Wan, F.H.; Zhang, Y.J.; Brown, J.K. Change in the biotype composition of Bemisia tabaci in Shandong province of China from 2005 to 2008. Environ. Entomol. 2010, 39, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Xue, X.; Ahmed, M.Z.; Ren, S.X.; Du, Y.Z.; Wu, J.H.; Cuthbertson, A.G.S.; Qiu, B.L. Host plants and natural enemies of Bemisia tabaci (Hemiptera: Aleyrodidae) in China. Insect Sci. 2011, 18, 101–120. [Google Scholar] [CrossRef]
- Perring, T.M.; Stansly, P.A.; Liu, T.X.; Smith, H.A.; Andreason, S.A. Chapter 4—Whiteflies: Biology, ecology, and management. In Sustainable Management of Arthropod Pests of Tomato; Elsevier Inc.: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2018; pp. 73–110. [Google Scholar]
- Nirgianaki, A.; Banks, G.K.; Frohlich, D.R.; Veneti, Z.; Braig, H.R.; Miller, T.A.; Bedford, I.D.; Markham, P.G.; Savakis, C.; Bourtzis, K. Wolbachia infections of the whitefly Bemisia tabaci. Curr. Microbiol. 2003, 47, 93–101. [Google Scholar] [PubMed]
- Weeks, A.R.; Velten, R.; Stouthamer, R. Incidence of a new sex–ratio–distorting endosymbiotic bacterium among arthropods. Proc. R. Soc. B 2003, 270, 1857–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zchori-Fein, E.; Perlman, S.J. Distribution of the bacterial symbiont Cardinium in arthropods. Mol. Ecol. 2004, 13, 2009–2016. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Ghanim, M.; Chiel, E.; Gerling, D.; Portnoy, V.; Steinberg, S.; Tzuri, G.; Horowitz, A.R.; Belausov, E.; Mozes-Daube, N.; et al. Identification and localization of a Rickettsia sp. in Bemisia tabaci (Homoptera: Aleyrodidae). Appl. Environ. Microb. 2006, 72, 3646–3652. [Google Scholar] [CrossRef] [Green Version]
- Bing, X.L.; Yang, J.; Zchori-Fein, E.; Wang, X.W.; Liu, S.S. Characterization of a newly discovered symbiont of the whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Appl. Environ. Microb. 2013, 79, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Oliver, K.M.; Pan, H.; Jiao, X.; Liu, B.; Xie, W.; Wang, S.; Wu, Q.; Xu, B.; White, J.A. Facultative symbiont Hamiltonella confers benefits to Bemisia tabaci (Hemiptera: Aleyrodidae), an invasive agricultural pest worldwide. Environ. Entomol. 2013, 42, 1265–1271. [Google Scholar] [CrossRef]
- Chiel, E.; Gottlieb, Y.; Zchori-Fein, E.; Mozes, D.N.; Katzir, N.; Inbar, M.; Ghanim, M. Biotype-dependent secondary symbiont communities in sympatric populations of Bemisia tabaci. Bull. Entomol. Res. 2007, 97, 407–413. [Google Scholar] [CrossRef]
- Himler, A.G.; Adachi, H.T.; Bergen, J.E.; Kozuch, A.; Kelly, S.E.; Tabashnik, B.E.; Chiel, E.; Duckworth, V.E.; Dennehy, T.J.; Zchori-Fein, E. Rapid spread of a bacterial symbiont in an invasive whitefly is driven by fitness benefits and female bias. Science 2011, 332, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, J.J.; Driscoll, T.P.; Verhoeve, V.I.; Utsuki, T.; Husseneder, C.; Chouljenko, V.N.; Azad, A.F.; Macaluso, K.R. Genomic diversification in strains of Rickettsia felis isolated from different arthropods. Genome Biol. Evol. 2014, 7, 35–56. [Google Scholar] [CrossRef]
- Shi, P.Q.; Wang, L.; Liu, Y.; An, X.; Chen, X.S.; Ahmed, M.Z.; Qiu, B.L.; Sang, W. Infection dynamics of endosymbionts reveal three novel localization patterns of Rickettsia during the development of whitefly Bemisia tabaci. FEMS Microbiol. Ecol. 2018, 94, fiy165. [Google Scholar] [CrossRef] [PubMed]
- Lawson, E.T.; Mousseau, T.A.; Klaper, R.; Hunter, M.D.; Werren, J.H. Rickettsia associated with male-killing in a buprestid beetle. Heredity 2001, 86, 497–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, L.A.; Werren, J.H.; Aebi, A.; Stone, G.N.; Jiggins, F.M. Evolution and diversity of Rickettsia bacteria. BMC Biol. 2009, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Nugnes, F.; Gebiola, M.; Monti, M.M.; Gualtieri, L.; Giorgini, M.; Wang, J.; Bernardo, U. Genetic diversity of the invasive gall wasp Leptocybe invasa (Hymenoptera: Eulophidae) and of its Rickettsia endosymbiont, and associated sex-ratio differences. PLoS ONE 2015, 10, e0124660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brumin, M.; Kontsedalov, S.; Ghanim, M. Rickettsia influences thermotolerance in the whitefly Bemisia tabaci B biotype. Insect Sci. 2011, 18, 57–66. [Google Scholar] [CrossRef]
- Chiel, E.; Inbar, M.; Mozes-Daube, N.; White, J.A.; Hunter, M.S.; Zchori-Fein, E. Assessments of fitness effects by the facultative symbiont Rickettsia in the sweetpotato whitefly (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2009, 102, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Xie, W.; Wang, S.; Wu, Q.; Liu, B.; Fang, Y.; Xu, B.; Zhang, Y. The endosymbiont Hamiltonella increases the growth rate of its host Bemisia tabaci during periods of nutritional stress. PLoS ONE 2014, 9, e89002. [Google Scholar] [CrossRef] [Green Version]
- Mahadav, A.; Gerling, D.; Gottlieb, Y.; Czosnek, H.; Ghanim, M. Parasitization by the wasp Eretmocerus mundus induces transcription of genes related to immune response and symbiotic bacteria proliferation in the whitefly Bemisia tabaci. BMC Genom. 2008, 9, 342. [Google Scholar] [CrossRef]
- Hendry, T.A.; Hunter, M.S.; Baltrus, D.A. The facultative symbiont Rickettsia protects an invasive whitefly against entomopathogenic Pseudomonas syringae strains. Appl. Environ. Microb. 2014, 80, 7161–7168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.D.; Guo, H.F. Importance of endosymbionts Wolbachia and Rickettsia in insect resistance development. Curr. Opin. Insect Sci. 2019, 33, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Dangelo, R.A.C.; Michereff-Filho, M.; Inoue-Nagata, A.K.; da Silva, P.S.; Chediak, M.; Guedes, R.N.C. Area-wide insecticide resistance and endosymbiont incidence in the whitefly Bemisia tabaci MEAM1 (B biotype): A Neotropical context. Ecotoxicology 2021, 30, 1056–1070. [Google Scholar] [CrossRef] [PubMed]
- Kanakala, S.; Ghanim, M. Global genetic diversity and geographical distribution of Bemisia tabaci and its bacterial endosymbionts. PLoS ONE 2019, 14, e0213946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Fan, Z.Y.; An, X.; Shi, P.Q.; Ahmed, M.Z.; Qiu, B.L. A single-pair method to screen Rickettsia-infected and uninfected whitefly Bemisia tabaci populations. J. Microbiol. Methods 2020, 168, 105797. [Google Scholar] [CrossRef] [PubMed]
- Caspi-Fluger, A.; Inbar, M.; Mozes-Daube, N.; Katzir, N.; Portnoy, V.; Belausov, E.; Hunter, M.S.; Zchori-Fein, E. Horizontal transmission of the insect symbiont Rickettsia is plant-mediated. Proc. R. Soc. B 2012, 279, 1791–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.M.; Fan, Z.Y.; O’Neill Rothenberg, D.; Peng, J.; Hafeez, M.; Chen, X.Y.; Pan, H.P.; Wu, J.H.; Qiu, B.L. Phototoxicity of ultraviolet-A against the whitefly Bemisia tabaci and its compatibility with an entomopathogenic fungus and whitefly parasitoid. Oxid. Med. Cell. Longev. 2021, 2021, 2060288. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Halhoul, M.N.; Kleinberg, I. Differential determination of glucose and fructose, and glucose- and fructose-yielding substances with anthrone. Anal. Biochem. 1972, 50, 337–343. [Google Scholar] [CrossRef]
- Ferreira, J.C.; Paschoalin, V.M.F.; Panek, A.D.; Trugo, L.C. Comparison of three different methods for trehalose determination in yeast extracts. Food Chem. 1997, 60, 251–254. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Northing, P.; Luo, W.; Cannon, R.J.C.; Walters, K.F.A. Leaf dipping as an environmental screening measure to test chemical efficacy against Bemisia tabaci on poinsettia plants. Int. J. Environ. Sci. Technol. 2009, 6, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelstädter, J.; Telschow, A. Cytoplasmic incompatibility and host population structure. Heredity 2009, 103, 196–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, P.Q.; Chen, X.Y.; Chen, X.S.; Lv, N.; Liu, Y.; Qiu, B.L. Rickettsia increases its infection and spread in whitefly populations by manipulating the defense patterns of the host plant. FEMS Microbiol. Ecol. 2021, 97, fiab032. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.A.; Clarke, H.K.; Turner, B.D.; Braig, H.R.; Perotti, B.M.A.; Clarke, H.K.; Turner, B.D.; Braig, H.R. Rickettsia as obligate and mycetomic. FASEB J. 2006, 20, 2372–2374. [Google Scholar] [CrossRef] [PubMed]
- Davison, H.R.; Pilgrim, J.; Wybouw, N.; Parker, J.; Pirro, S.; Hunter-Barnett, S.; Campbell, P.M.; Blow, F.; Darby, A.C.; Hurst, G.D.D.; et al. Genomic diversity across the Rickettsia and ‘Candidatus Megaira’ genera and proposal of genus status for the Torix group. Nat. Commun. 2022, 13, 2630. [Google Scholar] [CrossRef] [PubMed]
- Bockoven, A.A.; Bondy, E.C.; Flores, M.J.; Kelly, S.E.; Ravenscraft, A.M.; Hunter, M.S. What goes up might come down: The spectacular spread of an endosymbiont is followed by its decline a decade later. Microb. Ecol. 2020, 79, 482–494. [Google Scholar] [CrossRef]
- Giorgini, M.; Bernardo, U.; Monti, M.M.; Nappo, A.G.; Gebiola, M. Rickettsia symbionts cause parthenogenetic reproduction in the parasitoid wasp Pnigalio soemius (Hymenoptera: Eulophidae). Appl. Environ. Microb. 2010, 76, 2589–2599. [Google Scholar] [CrossRef] [Green Version]
- Cordaux, R.; Bouchon, D.; Grève, P. The impact of endosymbionts on the evolution of host sex-determination mechanisms. Trends Genet. 2011, 27, 332–341. [Google Scholar] [CrossRef]
- Hagimori, T.; Abe, Y.; Miura, K. The first finding of a Rickettsia bacterium associated with parthenogenesis induction among insects. Curr. Microbiol. 2006, 52, 97–101. [Google Scholar] [CrossRef]
- Adachi-Hagimori, T.; Miura, K. Limited mating ability of a wasp strain with Rickettsia-induced thelytoky. Ann. Entomol. Soc. Am. 2020, 113, 355–358. [Google Scholar] [CrossRef]
- Zhou, X.; Ling, X.; Guo, H.; Zhu-Salzman, K.; Ge, F.; Sun, Y. Serratia symbiotica enhances fatty acid metabolism of pea aphid to promote host development. Int. J. Mol. Sci. 2021, 22, 5951. [Google Scholar] [CrossRef] [PubMed]
- Fraga, A.; Ribeiro, L.; Lobato, M.; Santos, V.; Silva, J.R.; Gomes, H.; da Cunha Moraes, J.L.; de Souza Menezes, J.; de Oliveira, C.J.L.; Campos, E. Glycogen and glucose metabolism are essential for early embryonic development of the red flour beetle Tribolium castaneum. PLoS ONE 2013, 8, e65125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansford, R.G.; Johnson, R.N. The nature and control of the tricarboxylate cycle in beetle flight muscle. Biochem. J. 1975, 148, 389–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Sun, Q.; Gou, Y.; Zhang, K.; Zhang, Q.; Zhou, J.-J.; Liu, C. Long-term effect of elevated CO2 on the development and nutrition contents of the pea aphid (Acyrthosiphon pisum). Front. Physiol. 2021, 12, 688220. [Google Scholar] [CrossRef] [PubMed]
- Shukla, E.; Thorat, L.J.; Nath, B.B.; Gaikwad, S.M. Insect trehalase: Physiological significance and potential applications. Glycobiology 2015, 25, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Environ. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, Z.; Niu, H.; Guo, H. Win by quantity: A striking Rickettsia-bias symbiont community revealed by seasonal tracking in the whitefly Bemisia tabaci. Microb. Ecol. 2021, 81, 523–534. [Google Scholar] [CrossRef]
- Lv, N.; Wang, L.; Sang, W.; Liu, C.Z.; Qiu, B.L. Effects of endosymbiont disruption on the nutritional dynamics of the pea aphid Acyrthosiphon pisum. Insects 2018, 9, 161. [Google Scholar] [CrossRef]
- Ghanim, M.; Kontsedalov, S. Susceptibility to insecticides in the Q biotype of Bemisia tabaci is correlated with bacterial symbiont densities. Pest Manag. Sci. 2009, 65, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, D.Y.; Goryacheva, I.I.; Andrianov, B.V.; Reznik, N.L.; Lazebny, O.E.; Kulikov, A.M. The endosymbiotic bacterium Wolbachia enhances the nonspecific resistance to insect pathogens and alters behavior of Drosophila melanogaster. Russ. J. Genet. 2007, 43, 1066–1069. [Google Scholar] [CrossRef]
- Oliver, K.M.; Noge, K.; Huang, E.M.; Campos, J.M.; Becerra, J.X.; Hunter, M.S. Parasitic wasp responses to symbiont-based defense in aphids. BMC Biol. 2012, 10, 11. [Google Scholar] [CrossRef]
- Kontsedalov, S.; Zchori-Fein, E.; Chiel, E.; Gottlieb, Y.; Inbar, M.; Ghanim, M. The presence of Rickettsia is associated with increased susceptibility of Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides. Pest Manag. Sci. 2008, 64, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.P.; Chu, D.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, B.Y.; Zhang, Y.J. Relative amount of symbionts in insect hosts changes with host-plant adaptation and insecticide resistance. Environ. Entomol. 2013, 42, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xu, J.; Sun, T.; Ali, S. Synthesis of Cordyceps fumosorosea-biochar nanoparticles and their effects on growth and survival of Bemisia tabaci (Gennadius). Front. Microbiol. 2021, 12, 630220. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, C.L.; Ferrari, J.; Godfray, H.C.J. Aphid protected from pathogen by endosymbiont. Science 2005, 310, 1781. [Google Scholar] [CrossRef]
- Guay, J.-F.; Boudreault, S.; Michaud, D.; Cloutier, C. Impact of environmental stress on aphid clonal resistance to parasitoids: Role of Hamiltonella defensa bacterial symbiosis in association with a new facultative symbiont of the pea aphid. J. Insect Physiol. 2009, 55, 919–926. [Google Scholar] [CrossRef]
- Łukasik, P.; van Asch, M.; Guo, H.; Ferrari, J.; Charles, J. Godfray, H. Unrelated facultative endosymbionts protect aphids against a fungal pathogen. Ecol. Lett. 2013, 16, 214–218. [Google Scholar] [CrossRef]
- Vorburger, C.; Gehrer, L.; Rodriguez, P. A strain of the bacterial symbiont Regiella insecticola protects aphids against parasitoids. Biol. Lett. 2010, 6, 109–111. [Google Scholar] [CrossRef]
- Luo, C.; Luo, K.; Meng, L.; Wan, B.; Zhao, H.; Hu, Z. Ecological impact of a secondary bacterial symbiont on the clones of Sitobion avenae (Fabricius) (Hemiptera: Aphididae). Sci. Rep. 2017, 7, 40754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łukasik, P.; Dawid, M.A.; Ferrari, J.; Godfray, H.C.J. The diversity and fitness effects of infection with facultative endosymbionts in the grain aphid, Sitobion avenae. Oecologia 2013, 173, 985–996. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instar | Rickettsia +/− | LC50 (95% CI Conidia/mL) | Regression Virulence Model | χ2 | p |
|---|---|---|---|---|---|
| 1st | R− | 1.52 × 106 (5.77 × 105 − 4.30 × 106) | Y = 0.273x − 1.687 | 4.18 | 0.24 |
| R+ | 6.18 × 107 (1.93 × 107 − 3.87 × 108) | Y = 0.297x − 2.317 | 2.64 | 0.45 | |
| 2nd | R− | 3.19 × 106 (1.80 × 106 − 6.0 × 106) | Y = 0.488x − 3.172 | 4.80 | 0.19 |
| R+ | 1.28 × 107 (5.62 × 106 − 3.80 × 107) | Y = 0.357x − 2.54 | 6.19 | 0.10 | |
| 3rd | R− | 1.56 × 107 (2.37 × 106 − 1.29 × 109) | Y = 0.350x − 2.52 | 6.60 | 0.09 |
| R+ | 2.23 × 109 (1.65 × 108 − 1.36 × 1012) | Y = 0.188x − 1.754 | 0.50 | 0.92 | |
| 4th | R− | 1.82 × 108 (4.65 × 107 − 1.79 × 109) | Y = 0.294x − 2.426 | 5.42 | 0.14 |
| R+ | 4.93 × 1011 (3.41 × 109 − 8.07 × 1019) | Y = 0.151x − 1.768 | 1.404 | 0.70 |
| Pesticide | Instar | Rickettsia +/− | LC50 (95% CI) mg/L | Regression Virulence Model | χ2 | p |
|---|---|---|---|---|---|---|
| Imidacloprid | Adult | R− | 88.28 (65.14 − 132.22) | Y = 0.87x − 1.70 | 2.22 | 0.70 |
| R+ | 106.32 (83.28 − 144.95) | Y = 1.20x − 2.43 | 1.82 | 0.77 | ||
| 2nd nymph | R− | 34.89 (25.20 − 53.33) | Y = 0.76x − 1.17 | 3.38 | 0.50 | |
| R+ | 44.28 (31.79 − 69.72) | Y = 0.79x − 1.30 | 2.05 | 0.73 | ||
| Spirotetramat | Adult | R− | 97.97 (75.97 − 132.55) | Y = 1.15x − 2.28 | 2.03 | 0.73 |
| R+ | 120.14 (57.00 − 1336) | Y = 0.71x − 1.47 | 0.83 | 0.66 | ||
| 2nd nymph | R− | 13.24 (10.68 − 16.19) | Y = 1.24x − 1.39 | 2.69 | 0.61 | |
| R+ | 24.83 (19.46 − 32.56) | Y = 1.00x − 1.39 | 0.55 | 0.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Z.-Y.; Liu, Y.; He, Z.-Q.; Wen, Q.; Chen, X.-Y.; Khan, M.M.; Osman, M.; Mandour, N.S.; Qiu, B.-L. Rickettsia Infection Benefits Its Whitefly Hosts by Manipulating Their Nutrition and Defense. Insects 2022, 13, 1161. https://doi.org/10.3390/insects13121161
Fan Z-Y, Liu Y, He Z-Q, Wen Q, Chen X-Y, Khan MM, Osman M, Mandour NS, Qiu B-L. Rickettsia Infection Benefits Its Whitefly Hosts by Manipulating Their Nutrition and Defense. Insects. 2022; 13(12):1161. https://doi.org/10.3390/insects13121161
Chicago/Turabian StyleFan, Ze-Yun, Yuan Liu, Zi-Qi He, Qin Wen, Xin-Yi Chen, Muhammad Musa Khan, Mohamed Osman, Nasser Said Mandour, and Bao-Li Qiu. 2022. "Rickettsia Infection Benefits Its Whitefly Hosts by Manipulating Their Nutrition and Defense" Insects 13, no. 12: 1161. https://doi.org/10.3390/insects13121161
APA StyleFan, Z. -Y., Liu, Y., He, Z. -Q., Wen, Q., Chen, X. -Y., Khan, M. M., Osman, M., Mandour, N. S., & Qiu, B. -L. (2022). Rickettsia Infection Benefits Its Whitefly Hosts by Manipulating Their Nutrition and Defense. Insects, 13(12), 1161. https://doi.org/10.3390/insects13121161