


Comparative Studies of Perianal Structures in Myrmecophilous Aphids (Hemiptera, Aphididae)




Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Taxon Sampling
2.2. Material
2.3. Scanning Electron Microscopy
2.4. Light Microscopy
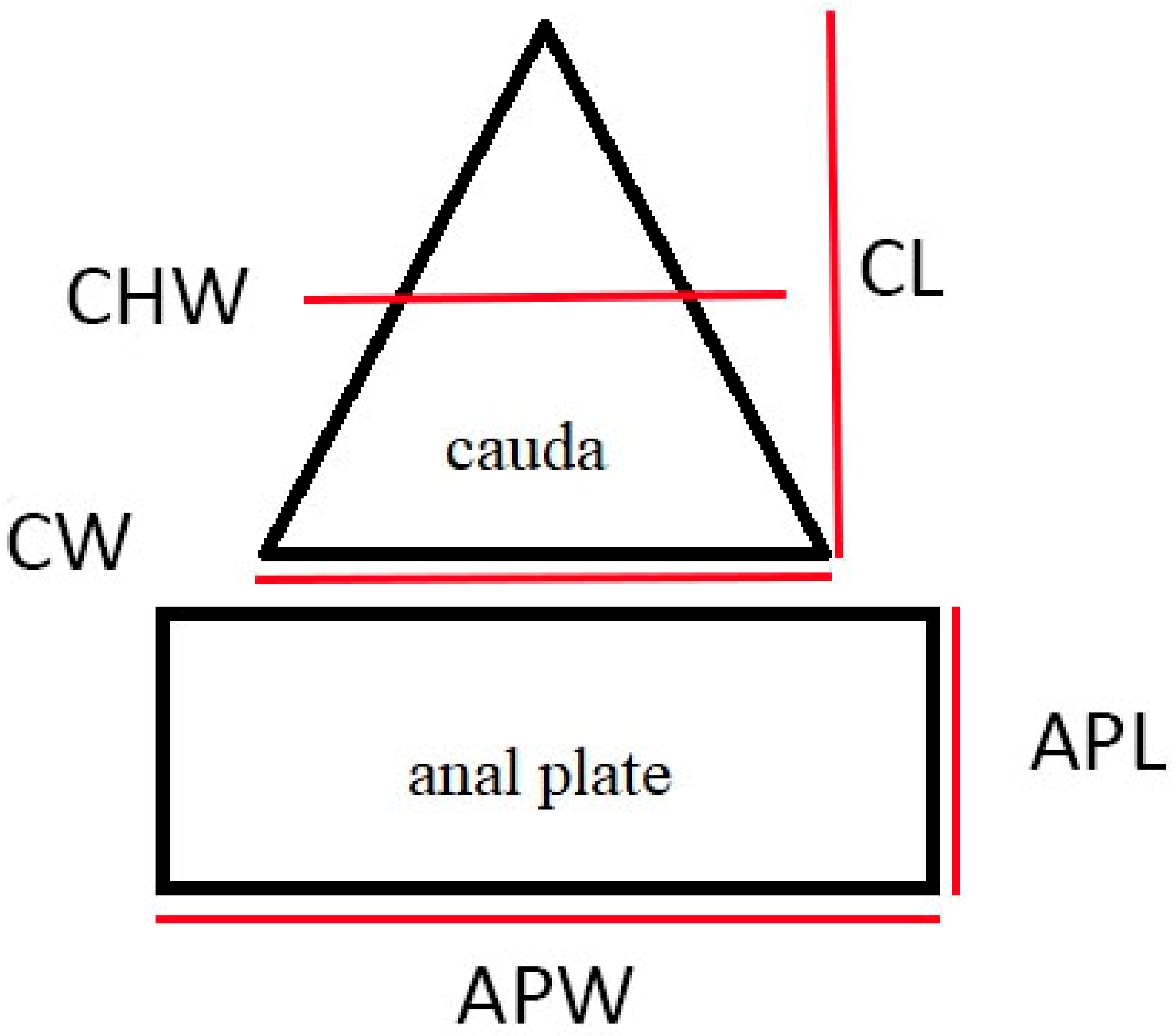
2.5. Morphological Measurements
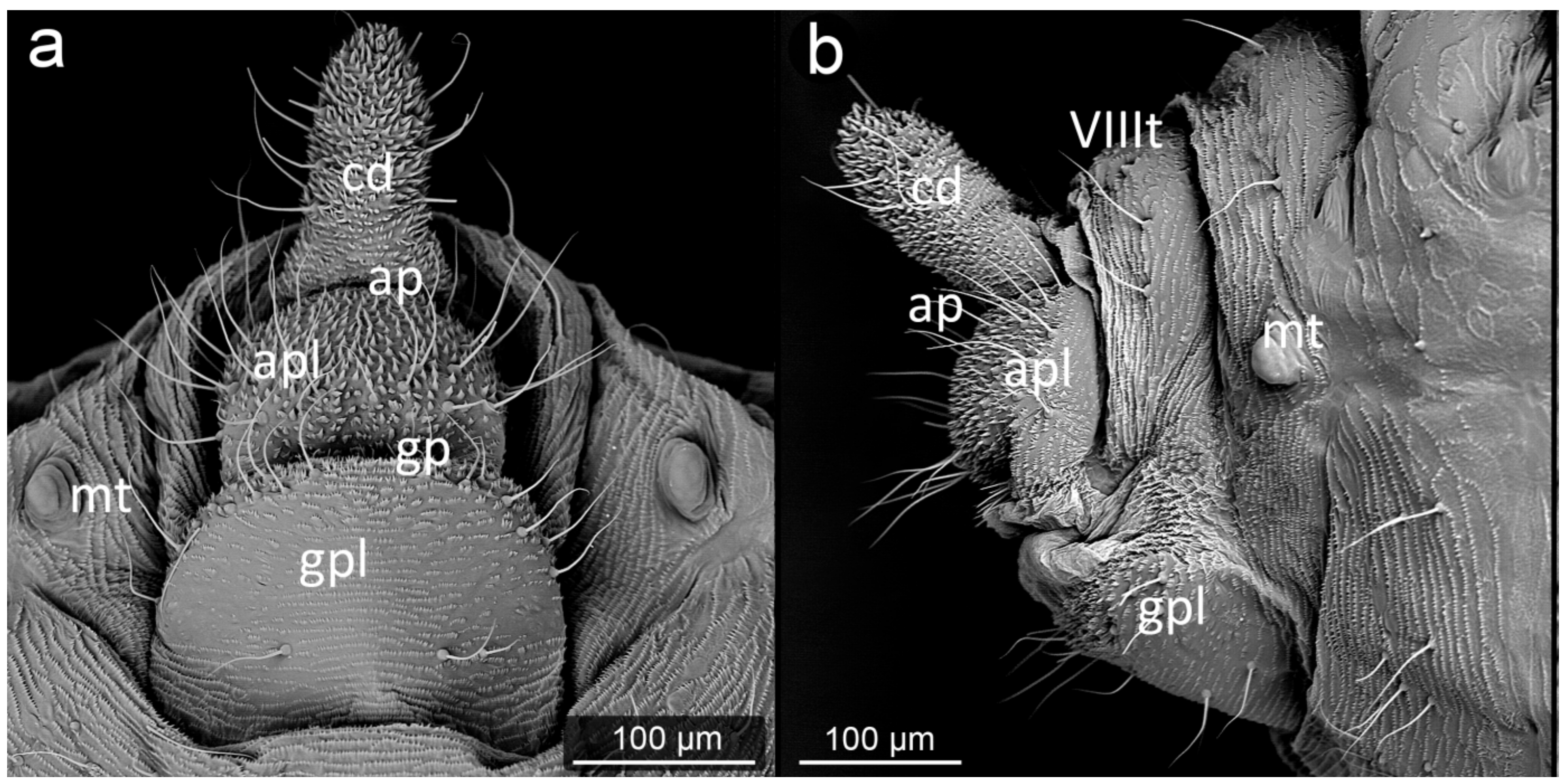
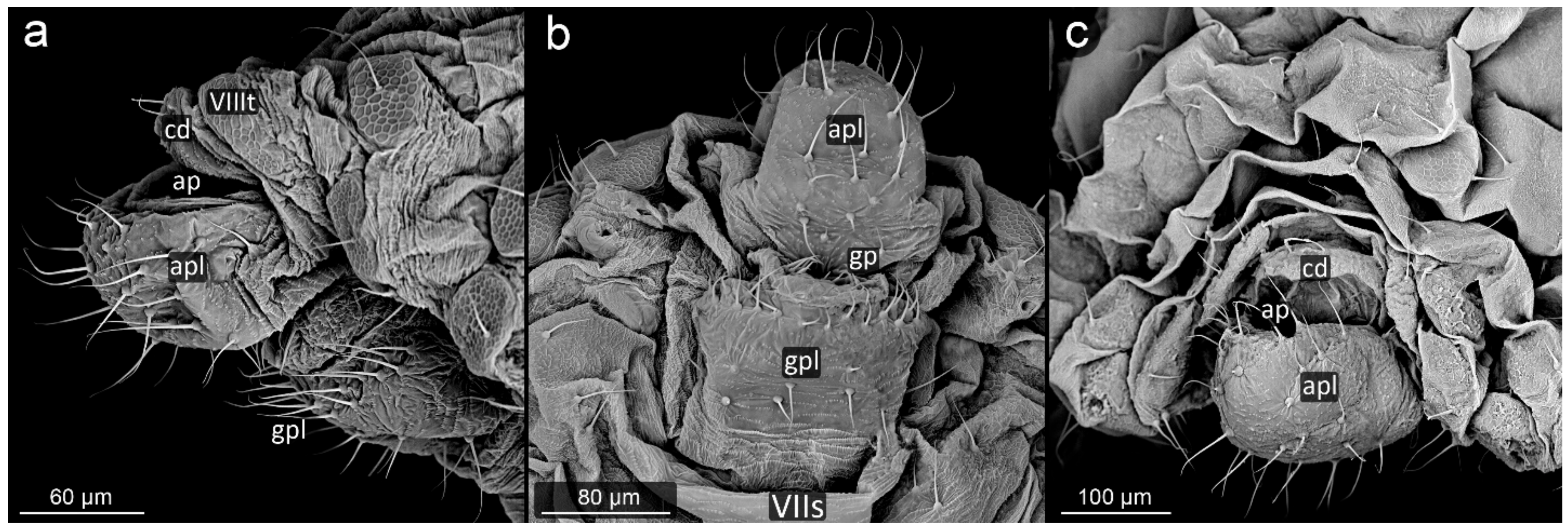
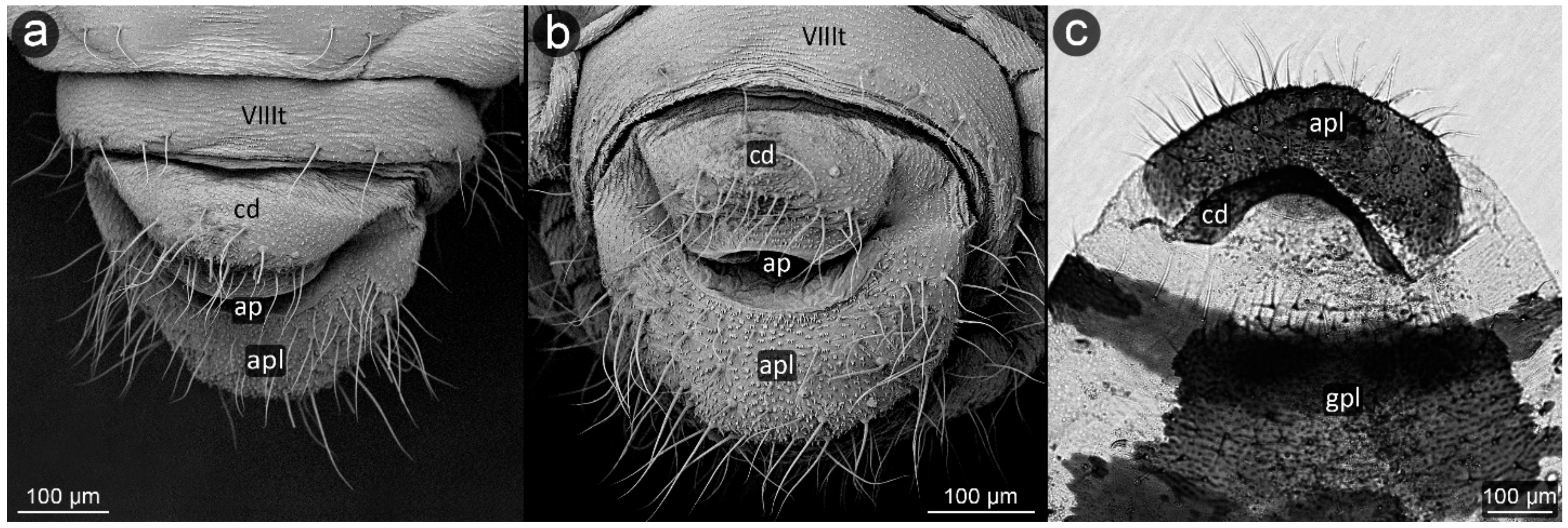
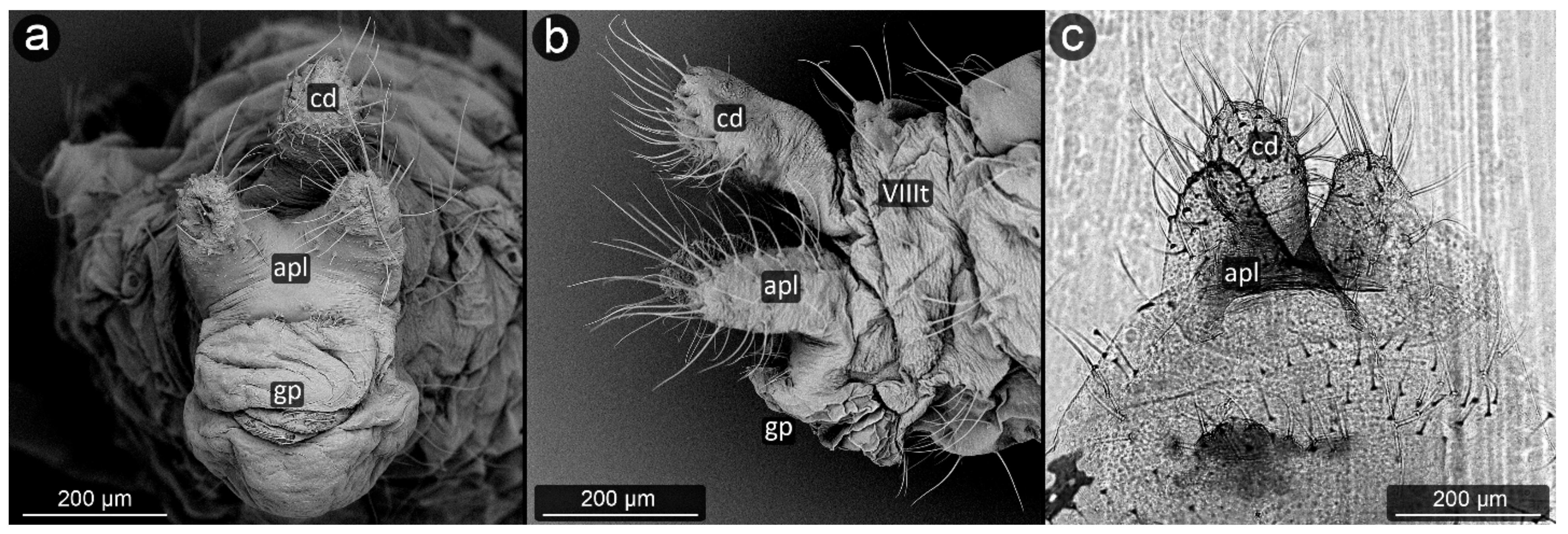
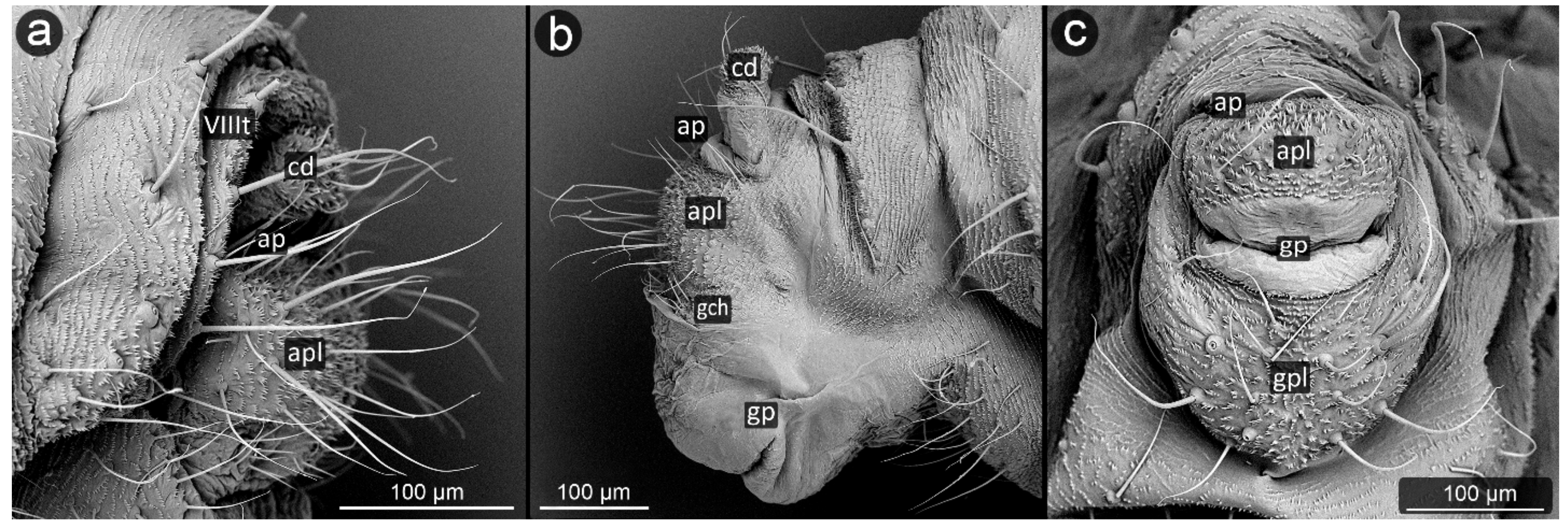
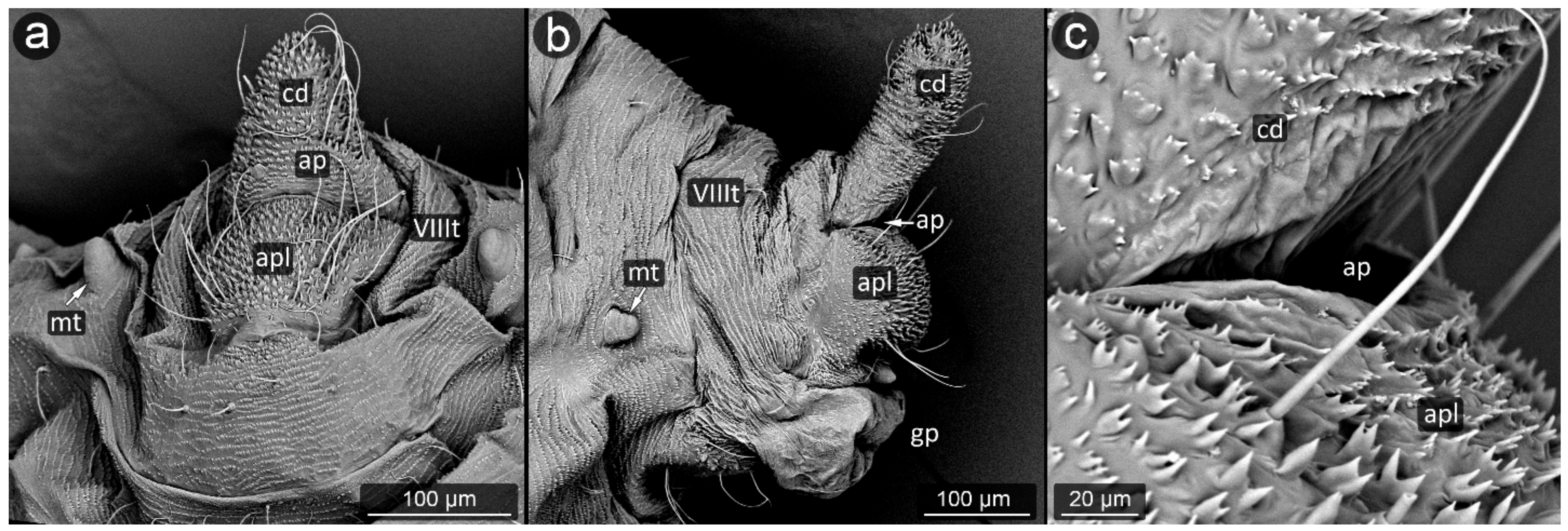
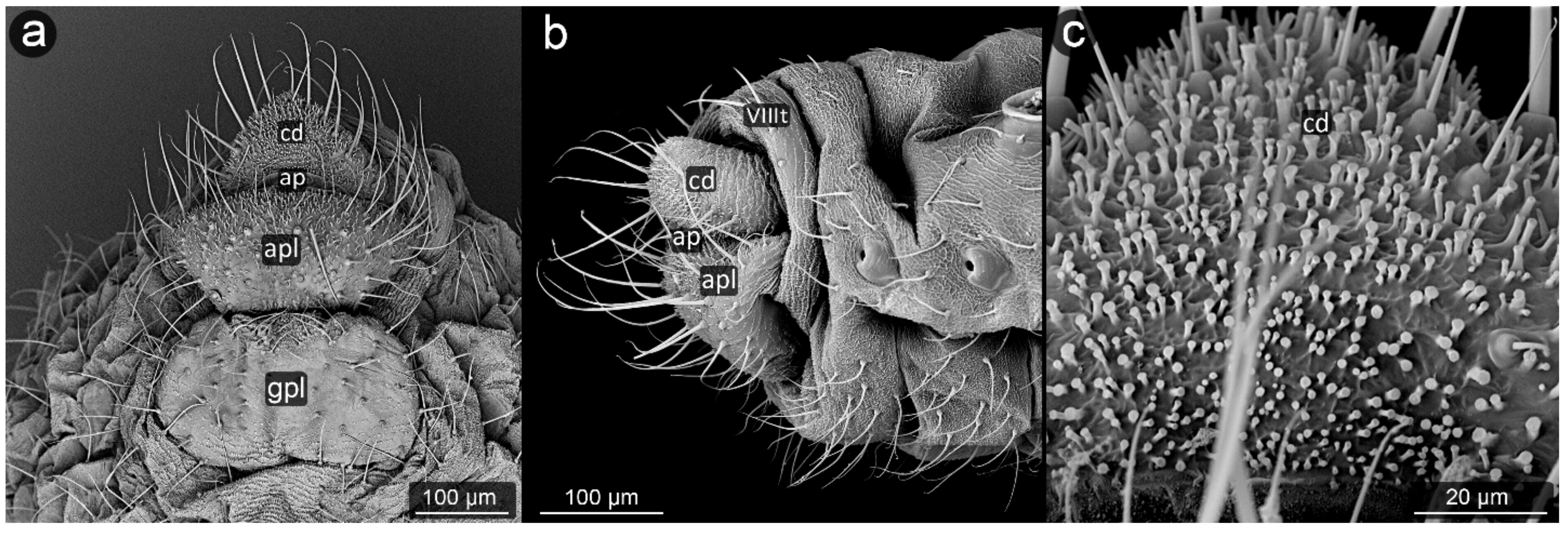
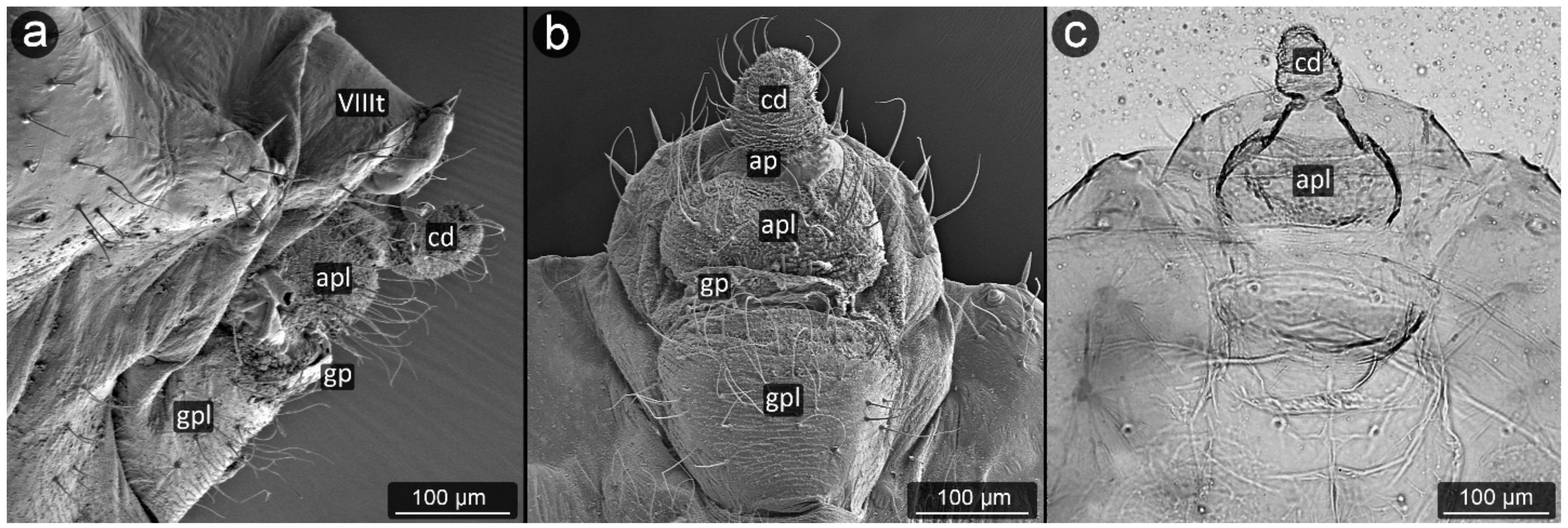
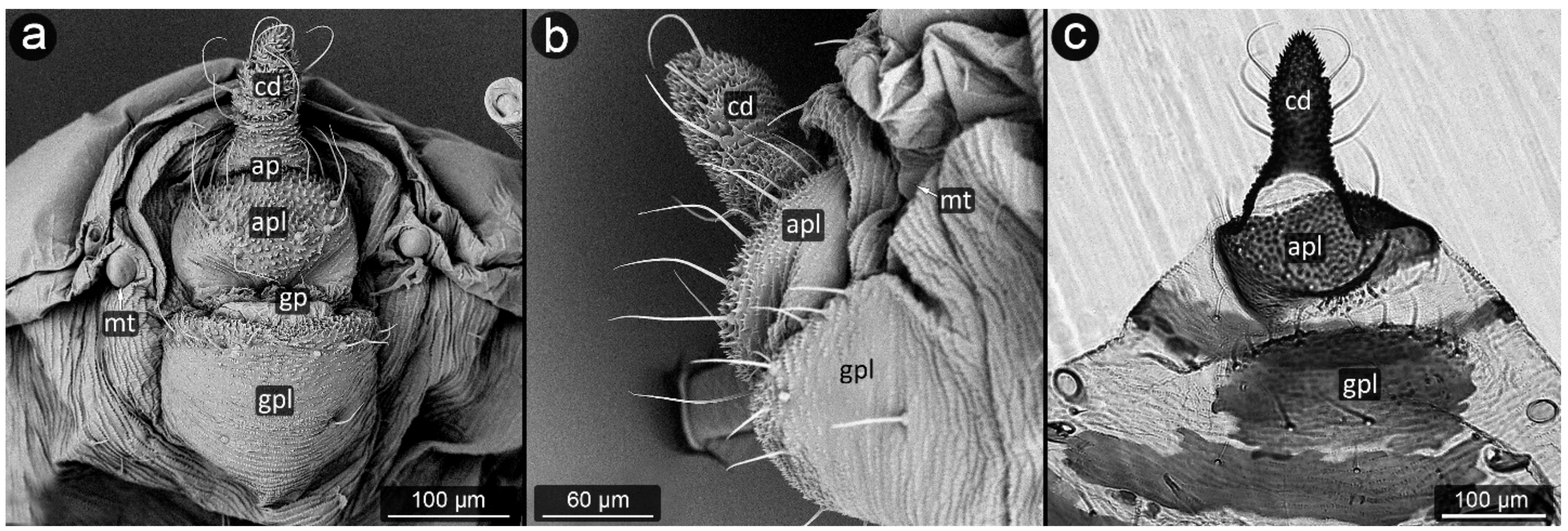
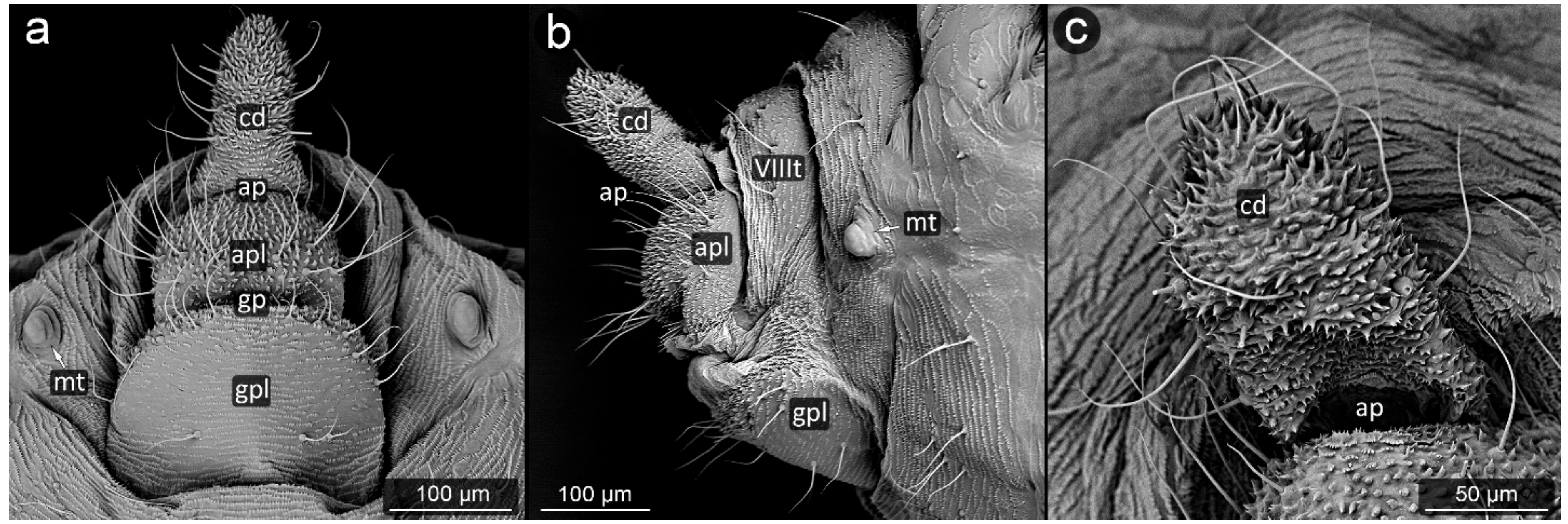
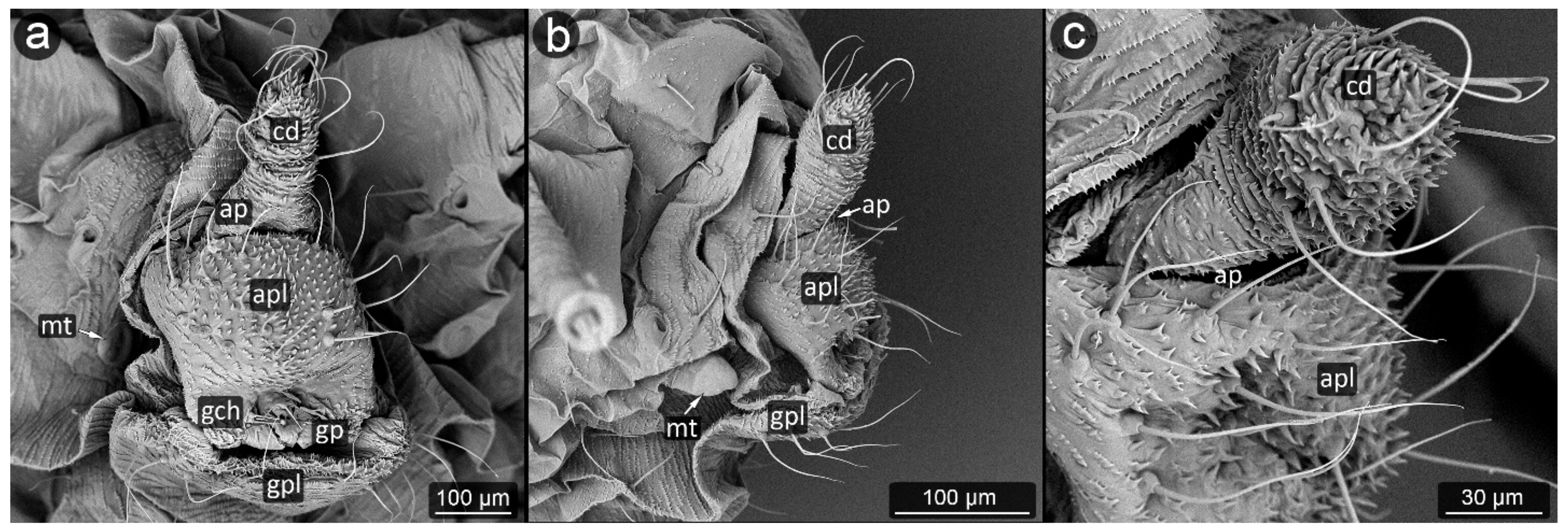
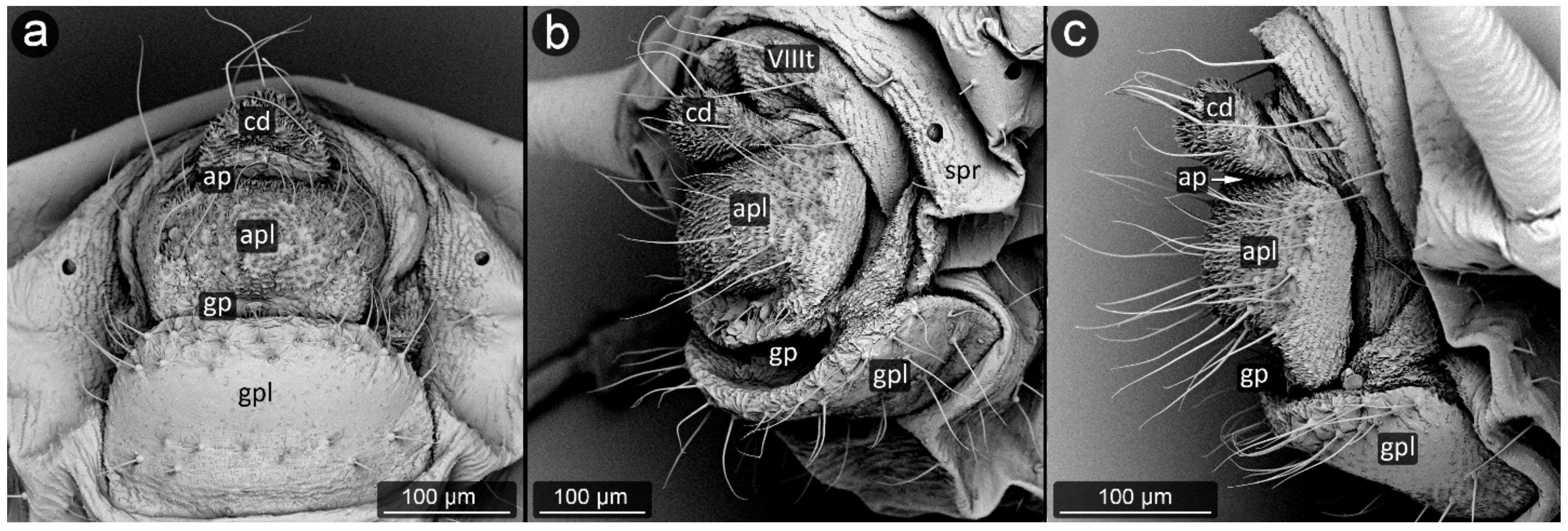
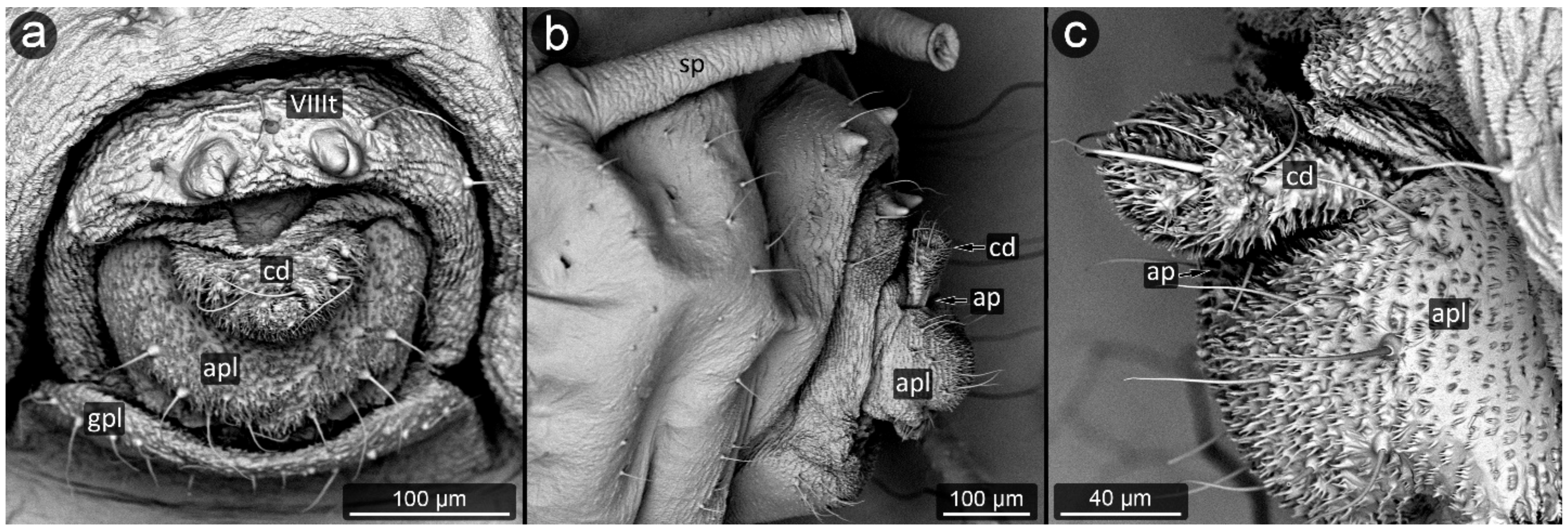
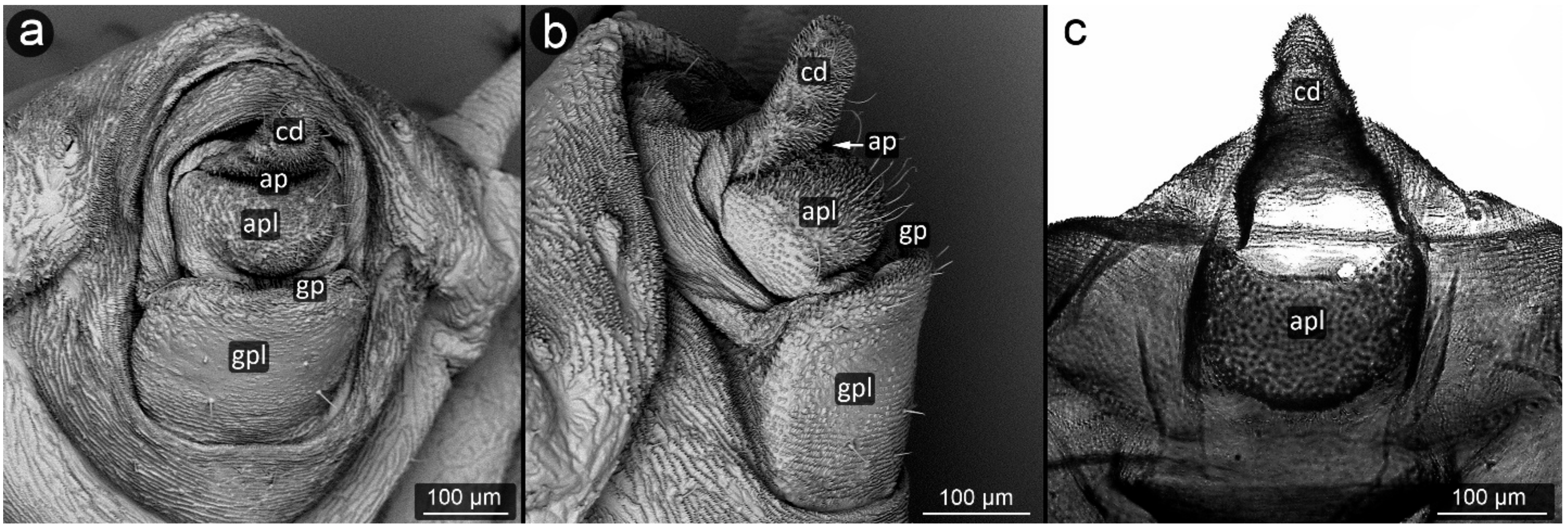
3. Results
3.1. Obligatorily Myrmecophilous Aphids
3.2. Facultatively Myrmecophilous Aphids
4. Discussion
4.1. Cauda and Anal Plate
4.2. Cuticle Surface and Setae of the Perianal Area
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stadler, B.; Dixon, A.F.G. Ecology and evolution of aphid–ant interactions. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 345–372. [Google Scholar] [CrossRef]
- Bronstein, J.L. Our current understanding of mutualism. Q. Rev. Biol. 1994, 69, 31–51. [Google Scholar] [CrossRef]
- Bronstein, J.L. The exploitation of mutualism. Ecol. Lett. 2001, 4, 277–287. [Google Scholar] [CrossRef]
- Thompson, J.N. The Coevolutionary Process; University of Chicago Press: Chicago, IL, USA, 1994; p. 376. [Google Scholar]
- Stadler, B.; Dixon, A.F.G. Mutualism. Ants and Their Insect Partners; Cambridge University Press: New York, NY, USA, 2008; p. 219. [Google Scholar]
- Banks, C.J. Effects of the ant Lasius niger (L.) on insects preying on small populations of Aphis fabae Scop. on bean plants. Ann. Appl. Entomol. 1962, 50, 669–679. [Google Scholar] [CrossRef]
- Nault, L.R.; Montgomery, M.E.; Bowers, W.S. Ant-aphid association: Role of aphid alarm pheromone. Science 1976, 192, 1349–1351. [Google Scholar] [CrossRef]
- Dixon, A.F.G.; Agarwala, B.K. Ladybird-induced life-history changes in aphids. Proc. R. Soc. B Boil. Sci. 1999, 266, 1549–1553. [Google Scholar] [CrossRef]
- Yao, I.; Shibao, H.; Akimoto, S. Costs and benefits of ant attendance to the drepanosiphid aphid Tuberculatus quercicola. Oikos 2000, 89, 3–10. [Google Scholar] [CrossRef]
- Katayama, N.; Suzuki, N. Cost and benefit of ant attendance for Aphis craccivora (Hemiptera: Aphididae) with reference to aphid colony size. Can. Entomol. 2002, 134, 241–249. [Google Scholar] [CrossRef]
- Matsuura, K.; Yashiro, T. Aphid egg protection by ants: A novel aspect of the mutualism between the tree-feeding aphid Stomaphis hirukawai and its attendant ant Lasius productus. Naturwissenschaften 2006, 93, 506–510. [Google Scholar] [CrossRef]
- Nielsen, C.; Agrawal, A.A.; Hajek, A.E. Ants defend aphids against lethal disease. Biol. Lett. 2010, 6, 205–208. [Google Scholar] [CrossRef]
- Bilska, A.; Francikowski, J.; Wyglenda, A.; Masłowski, A.; Kaszyca, N.; Depa, Ł. Aphids playing possum—Defensive or mutualistic response? J. Insect Behav. 2018, 31, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Miñarro, M.; Fernández-Mata, G.; Medina, P. Role of ants in structuring the aphid community on apple. Ecol. Entomol. 2010, 35, 206–215. [Google Scholar] [CrossRef]
- Nixon, G.E.J. The Association of Ants with Aphids and Coccids; Commonwealth Institute of Entomology: London, UK, 1951; pp. 1–36. [Google Scholar]
- Way, M.J. Mutualism between ants and honeydew-producing Homoptera. Annu. Rev. Entomol. 1963, 8, 307–344. [Google Scholar] [CrossRef]
- Yao, I.; Akimoto, S.I. Flexibility in the composition and concentration of amino acids in honeydew of the drepanosiphid aphid Tuberculatus quercicola. Ecol. Entomol. 2002, 27, 745–752. [Google Scholar] [CrossRef]
- Douglas, A.E. The nutritional physiology of aphids. Adv. Insect Physiol. 2003, 31, 73–140. [Google Scholar]
- Woodring, J.; Wiedemann, R.; Fischer, M.K.; Hoffmann, K.H.; Völkl, W. Honeydew amino acids in relation to sugars and their role in the establishment of ant-attendance hierarchy in eight species of aphids feeding on tansy (Tanacetum vulgare). Physiol. Entomol. 2004, 29, 311–319. [Google Scholar] [CrossRef]
- Fischer, M.K.; Völkl, W.; Hoffmann, K.H. Honeydew production and honeydew sugar composition of polyphagous black bean aphid, Aphis fabae (Hemiptera: Aphididae) on various host plants and implications for ant-attendance. Eur. J. Entomol. 2005, 102, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Mondor, E.B.; Rosenheim, J.A.; Addicott, J.F. Mutualist-induced transgenerational polyphenisms in cotton aphid populations. Funct. Ecol. 2008, 22, 157–162. [Google Scholar] [CrossRef]
- Detrain, C.; Verheggen, F.J.; Diez, L.; Wathelet, B.; Haubruge, E. Aphid–ant mutualism: How honeydew sugars influence the behaviour of ant scouts. Physiol. Entomol. 2010, 35, 168–174. [Google Scholar] [CrossRef]
- Fischer, M.K.; Shingleton, A.W. Host plant and ants influence the honeydew sugar composition of aphids. Funct. Ecol. 2001, 15, 544–550. [Google Scholar] [CrossRef]
- Bristow, C.M. Why are so few aphids ant-tended? In Ant–Plant Interactions; Huxley, C.R., Cutler, D.F., Eds.; Oxford University Press: Oxford, UK, 1991; pp. 104–119. [Google Scholar]
- Stadler, B. The relative importance of host plants, natural enemies and ants in the evolution of life-history characters in aphids. In Vertical Food Web Interactions; Dettner, K., Bauer, G., Völkl, W., Eds.; Springer: Berlin, Germany, 1997; pp. 241–256. [Google Scholar]
- Vantaux, A.; Van den Ende, W.; Billen, J.; Wenseleers, T. Large interclone differences in melezitose secretion in the facultatively ant-tended black bean aphid Aphis fabae. J. Insect Physiol. 2011, 57, 1614–1621. [Google Scholar] [CrossRef]
- Depa, Ł.; Kaszyca-Taszakowska, N.; Taszakowski, A.; Kanturski, M. Ant-induced evolutionary patterns in aphids. Biol. Rev. 2020, 95, 1574–1589. [Google Scholar] [CrossRef]
- Lorenz, H.; Scheurer, S. Biology and generation-order of Stomaphis quercus (Lachnidae) living on Betula pendula near Berlin, Germany. In Aphids in Natural and Managed Ecosystems; Nieto Nafria, J.M., Dixon, A.F.G., Eds.; Secretariado de publicaciones, Universidad de León: León, Spain, 1998; pp. 243–250. [Google Scholar]
- Stadler, B.; Kindlmann, P.; Ŝmilauer, P.; Fiedler, K. A comparative analysis of morphological and ecological characters of European aphids and lycaenids in relationship to ant attendance. Oecologia 2003, 135, 422–430. [Google Scholar] [CrossRef]
- Kloft, W. Die Trophobiose zwischen Waldameisen und Pflanzenläusen mit Untersuchungen über die Wechselwirkungen zwischen Pflanzenläusen und Pflanzengeweben. Entomophaga 1960, 5, 43–54. [Google Scholar] [CrossRef]
- Kanturski, M.; Karcz, J.; Kaszyca, N.; Depa, Ł. Perianal structures in myrmecophilous subterranean aphids (Insecta: Hemiptera: Aphididae)—Comparative morphology of trophobiotic organ with its first description in Lachninae. Arthropod Struct. Dev. 2017, 46, 496–507. [Google Scholar] [CrossRef]
- Zwölfer, H. Zur Systematik, Biologie und Oekologie unterirdisch lebender Aphiden (Homoptera; Aphidoidea). I. Z. Angew. Entomol. 1958, 40, 182–221. [Google Scholar] [CrossRef]
- Zwölfer, H. Zur Systematik, Biologie und Oekologie unterirdisch lebender Aphiden (Homoptera; Aphidoidea). II. Z. Angew. Entomol. 1958, 40, 528–575. [Google Scholar] [CrossRef]
- Zwölfer, H. Zur Systematik, Biologie und Oekologie unterirdisch lebender Aphiden (Homoptera; Aphidoidea). III. Z. Angew. Entomol. 1958, 42, 129–172. [Google Scholar] [CrossRef]
- Zwölfer, H. Zur Systematik, Biologie und Oekologie unterirdisch lebender Aphiden (Homoptera; Aphidoidea). IV. Z. Angew. Entomol. 1958, 43, 1–43. [Google Scholar] [CrossRef]
- Kaszyca-Taszakowska, N.; Yamamoto, T.; Depa, Ł. Trophobiotic organ and perianal structures in aphid genus Stomaphis (Walker). Micron 2020, 138, 102930. [Google Scholar] [CrossRef] [PubMed]
- Heie, O.E. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark. I. In Fauna Entomologica Scandinavica; Scandinavian Science Press: Klampenborg, Denmark, 1980; Volume 9, pp. 1–236. [Google Scholar]
- Heie, O.E. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark. II. In Fauna Entomologica Scandinavica; Scandinavian Science Press: Klampenborg, Denmark, 1982; Volume 11, pp. 1–169. [Google Scholar]
- Heie, O.E. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark. III. In Fauna Entomologica Scandinavica; Scandinavian Science Press: Klampenborg, Denmark, 1986; Volume 17, pp. 1–314. [Google Scholar]
- Heie, O.E. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark. IV. In Fauna Entomologica Scandinavica; Scandinavian Science Press: Klampenborg, Denmark, 1992; Volume 25, pp. 1–189. [Google Scholar]
- Heie, O.E. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark. V. In Fauna Entomologica Scandinavica; Scandinavian Science Press: Klampenborg, Denmark, 1994; Volume 28, pp. 1–242. [Google Scholar]
- Heie, O.E. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark. VI. In Fauna Entomologica Scandinavica; Scandinavian Science Press: Klampenborg, Denmark, 1995; Volume 31, pp. 1–222. [Google Scholar]
- Blackman, R.L.; Eastop, V. Aphidsonworldsplants.info. 2022. Available online: http://www.aphidsonworldsplants.info (accessed on 20 June 2022).
- Depa, Ł.; Wojciechowski, W. Aphids (Hemiptera; Aphidinea) of Garb Tarnogórski and their trophobiotic relations with ants. Ann. Up. Siles. Mus. Entomol. 2009, 18, 106. [Google Scholar]
- Kanturski, M.; Wieczorek, K. Metody i preparowania mszyc (Hemiptera, Aphidoidea) w badaniach faunistycznych, taksonomicznych i molekularnych. Młodzi Nauk. Dla Pol. Nauk. 2012, 8, 137–143. [Google Scholar]
- Wojciechowski, W. A Monograph of the Palearctic Pterocommatinae (Aphididae, Aphidinea, Hemiptera); Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2003; p. 112. [Google Scholar]
- Wojciechowski, W.; Depa, Ł.; Halgoš, J.; Matečný, I.; Lukáš, J.; Kanturski, M. Aphids of Slovakia. Distributional Catalogue, Checklist, Keys and List of Host Plants; Comenius University in Bratislava: Bratislava, Slovakia, 2016; p. 346. [Google Scholar]
- Favret, C. Aphid Species File. Version 5.0/5.0. Available online: http://Aphid.SpeciesFile.org (accessed on 20 June 2021).
- Mróz, E.; Trela, J.; Depa, Ł. Taxonomic analysis of Lachnus pallipes/longirostris-roboris complex (Hemiptera, Aphididae, Lachninae), with the redescription of sexual morphs and new synonymy. Zool. Anz. 2015, 254, 51–61. [Google Scholar] [CrossRef]
- Kanturski, M.; Karcz, J.; Wieczorek, K. Morphology of European species of the aphid genus Eulachnus (Hemiptera: Aphididae: Lachninae)—A SEM comparative and integration study. Micron 2015, 76, 23–36. [Google Scholar] [CrossRef]
- Szelegiewicz, H. Klucze do Oznaczania Owadów Polski. XVII, Homoptera, 5a, Mszyce–Aphidodea, 1, Lachnidae; Polskie Towarzystwo Entomologiczne, PWN: Warszawa, Poland, 1978; p. 107. (In Polish) [Google Scholar]
- Miyazaki, M. Morphology of aphids. In Aphids. Their Biology, Natural Enemies and Control; Harrewijn, P., Minks, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; Volume 2A, pp. 1–25. [Google Scholar]
- Watanabe, S.; Murakami, Y.; Hasegawa, E. Effects of attending ant species on the fate of colonies of an aphid Macrosiphoniella yomogicola (Matsumura) (Homoptera: Aphididae), in an ant-aphid symbiosis. Entomol. News 2019, 128, 325–332. [Google Scholar] [CrossRef]
- Wool, D.; Manheim, O.; Inbar, M. Return flight of sexuparae of galling aphids to their primary host trees: Implications for differential herbivory and gall (Aphidoidea: Pemphigidae: Fordinae) abundance. Ann. Entomol. Soc. Am. 1997, 90, 341–350. [Google Scholar] [CrossRef]
- Shingleton, A.W.; Stern, D.L. Molecular phylogenetic evidence for multiple gains and losses of ant mutualism within the aphid genus Chaitophorus. Mol. Phylogenet. Evol. 2003, 26, 26–35. [Google Scholar] [CrossRef]
- Kondo, T.; Williams, M.L. Redescription of the myrmecophilous soft scale insect: Aztecalecanium colimae (Cockerell), new genus and new combination (Hemiptera: Coccoidea: Coccidae). Tip Rev. Espec. Cienc. Quím-Biol. 2004, 7, 5–9. [Google Scholar]
- Ammar, E.-D.; Alessandro, R.; Shatters, R.G.; Hall, D.G. Behavioral, ultrastructural and chemical studies on the honeydew and waxy secretions by nymphs and adults of the Asian citrus psyllid Diaphorina citri (Hemiptera: Psyllidae). PLoS ONE 2013, 8, e64938. [Google Scholar] [CrossRef] [Green Version]
- Heie, O.E. Morphological structures and adaptations. In Aphids. Their Biology, Natural Enemies and Control; Harrewijn, P., Minks, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; Volume 2A, pp. 393–400. [Google Scholar]
- Neinhuis, C.; Barthlott, W. Characterization and distribution of water-repellent, self-cleaning plant surfaces. Ann. Bot. 1997, 79, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Darmanin, T.; Guittard, F. Superhydrophobic and superoleophobic properties in nature. Mater. Today 2015, 18, 273–285. [Google Scholar] [CrossRef]































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Species | om | fm | om | fm | om | fm | om | fm | |
|---|---|---|---|---|---|---|---|---|---|---|
| obligatorily myrmecophilous (om) | facultatively myrmecophilous (fm) | apl/cl | apl/apw | cl/cw | cw/chw | |||||
| Thelaxinae | Glyphina betulae (Linnaeus, 1758) | 0.43 | 0.26 | 1.27 | 1.89 | |||||
| Thelaxes dryophila (Schrank, 1801) | 0.63 | 0.41 | 1.08 | 0.56 | ||||||
| Eriosomatinae | Prociphilus bumeliae (Schrank, 1801) | 4.23 | 0.67 | 0.54 | 0.80 | |||||
| Prociphilus fraxini (Fabricius, 1777) | 2.37 | 1.19 | 0.78 | 0.73 | ||||||
| Calaphidinae | Symydobius oblongus (von Heyden, 1837) | 2.36 | 1.74 | 0.53 | 0.81 | |||||
| Panaphis juglandis (Goeze, 1778) | 1.09 | 1.39 | 2.31 | 0.52 | ||||||
| Chaitophorinae | Chaitophorus nassonowi Mordvilko, 1894 | 2.51 | 2.37 | 1.38 | 0.52 | |||||
| Chaitophorus populeti (Panzer, 1801) | 1.73 | 2.43 | 1.91 | 0.41 | ||||||
| Aphidinae | Aphis acetosae Linnaeus, 1761 | Aphis craccivora Koch, 1854 | 0.67 | 1.28 | 0.61 | 2.01 | 2.44 | 1.93 | 0.58 | 0.75 |
| Aphis jacobaeae Schrank, 1801 | Aphis fabae Scopoli, 1763 | 0.77 | 1.16 | 0.97 | 1.86 | 2.41 | 1.42 | 0.56 | 0.63 | |
| Aphis pomi De Geer, 1773 | Aphis hederae Kaltenbach, 1843 | 0.98 | 1.18 | 1.79 | 1.82 | 2.69 | 1.68 | 0.63 | 0.58 | |
| Aphis sedi Kaltenbach, 1843 | Rhopalosiphum padi (Linnaeus, 1758) | 0.59 | 0.98 | 0.57 | 0.64 | 2.38 | 1.14 | 0.77 | 0.54 | |
| Anuraphis catonii Hille Ris Lambers, 1935 | Brachycaudus cardui (Linnaeus, 1758) | 2.63 | 1.74 | 1.72 | 0.92 | 0.73 | 0.74 | 0.69 | 0.73 | |
| Brachycaudus tragopogonis (Kaltenbach, 1843) | 1.08 | 0.42 | 0.81 | 0.80 | ||||||
| Metopeurum fuscoviride Stroyan, 1950 | Dysaphis anthrisci Börner, 1950 | 0.96 | 2.18 | 0.92 | 2.31 | 1.46 | 0.94 | 0.51 | 0.80 | |
| Dysaphis plantaginea (Passerini, 1860) | 2.01 | 2.52 | 0.97 | 0.70 | ||||||
| Semiaphis dauci (Fabricius, 1775) | Dysaphis sorbi (Kaltenbach, 1843) | 0.62 | 1.55 | 0.52 | 1.48 | 1.96 | 0.87 | 0.62 | 0.60 | |
| Myzus cerasi (Fabricius, 1775) | 1.15 | 1.81 | 1.39 | 0.56 | ||||||
| Pterocomma konoi Hori, 1939 | Pterocomma rufipes (Hartig, 1841) | 2.42 | 2.53 | 2.11 | 2.73 | 0.83 | 0.57 | 0.77 | 0.70 | |
| Lachninae | Cinara pini (Linnaeus, 1758) | 1.76 | 1.63 | 0.61 | 0.68 | |||||
| Lachnus pallipes (Hartig, 1841) | 3.00 | 2.13 | 0.61 | 0.82 | ||||||
| mean for all samples: | 1.43 | 1.86 | 1.29 | 1.66 | 1.49 | 1.08 | 0.71 | 0.68 | ||
| mean for Aphidinae | 1.19 | 1.58 | 1.07 | 1.81 | 1.75 | 1.17 | 0.66 | 0.66 | ||
| x > 1—anal plate longer than cauda | x > 1—cauda longer than its base | |||||||||
| x > 1—anal plate longer than wide | x < 1—cauda is narrowing towards apex | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaszyca-Taszakowska, N.; Kanturski, M.; Depa, Ł. Comparative Studies of Perianal Structures in Myrmecophilous Aphids (Hemiptera, Aphididae). Insects 2022, 13, 1160. https://doi.org/10.3390/insects13121160
Kaszyca-Taszakowska N, Kanturski M, Depa Ł. Comparative Studies of Perianal Structures in Myrmecophilous Aphids (Hemiptera, Aphididae). Insects. 2022; 13(12):1160. https://doi.org/10.3390/insects13121160
Chicago/Turabian StyleKaszyca-Taszakowska, Natalia, Mariusz Kanturski, and Łukasz Depa. 2022. "Comparative Studies of Perianal Structures in Myrmecophilous Aphids (Hemiptera, Aphididae)" Insects 13, no. 12: 1160. https://doi.org/10.3390/insects13121160
APA StyleKaszyca-Taszakowska, N., Kanturski, M., & Depa, Ł. (2022). Comparative Studies of Perianal Structures in Myrmecophilous Aphids (Hemiptera, Aphididae). Insects, 13(12), 1160. https://doi.org/10.3390/insects13121160