

Use of a Sprayable Sex Pheromone Formulation in Landscape-Level Control of Choristoneura fumiferana Populations

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
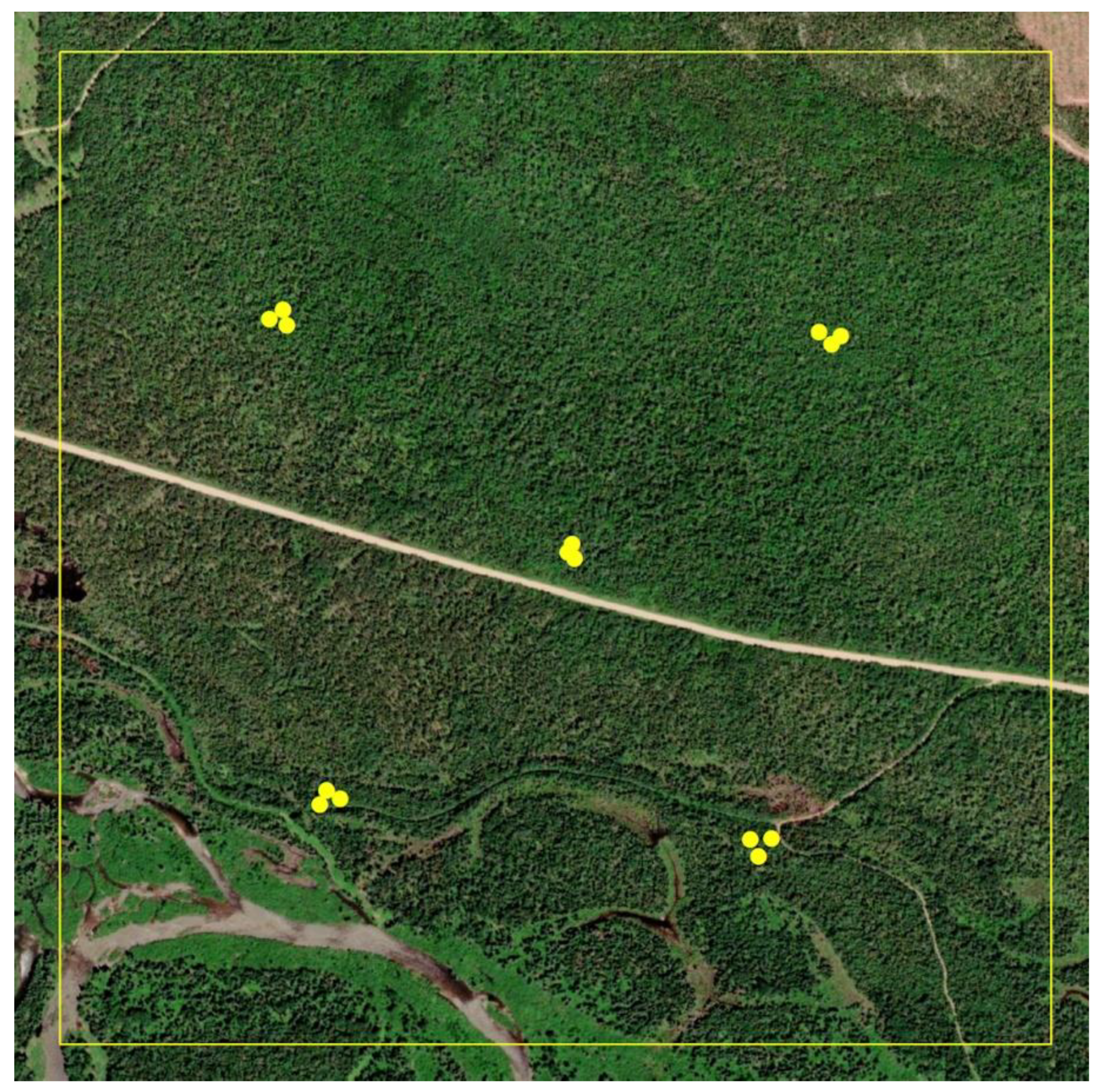
2.1. Sites
2.2. Population Measurements
2.3. Pheromone Monitoring
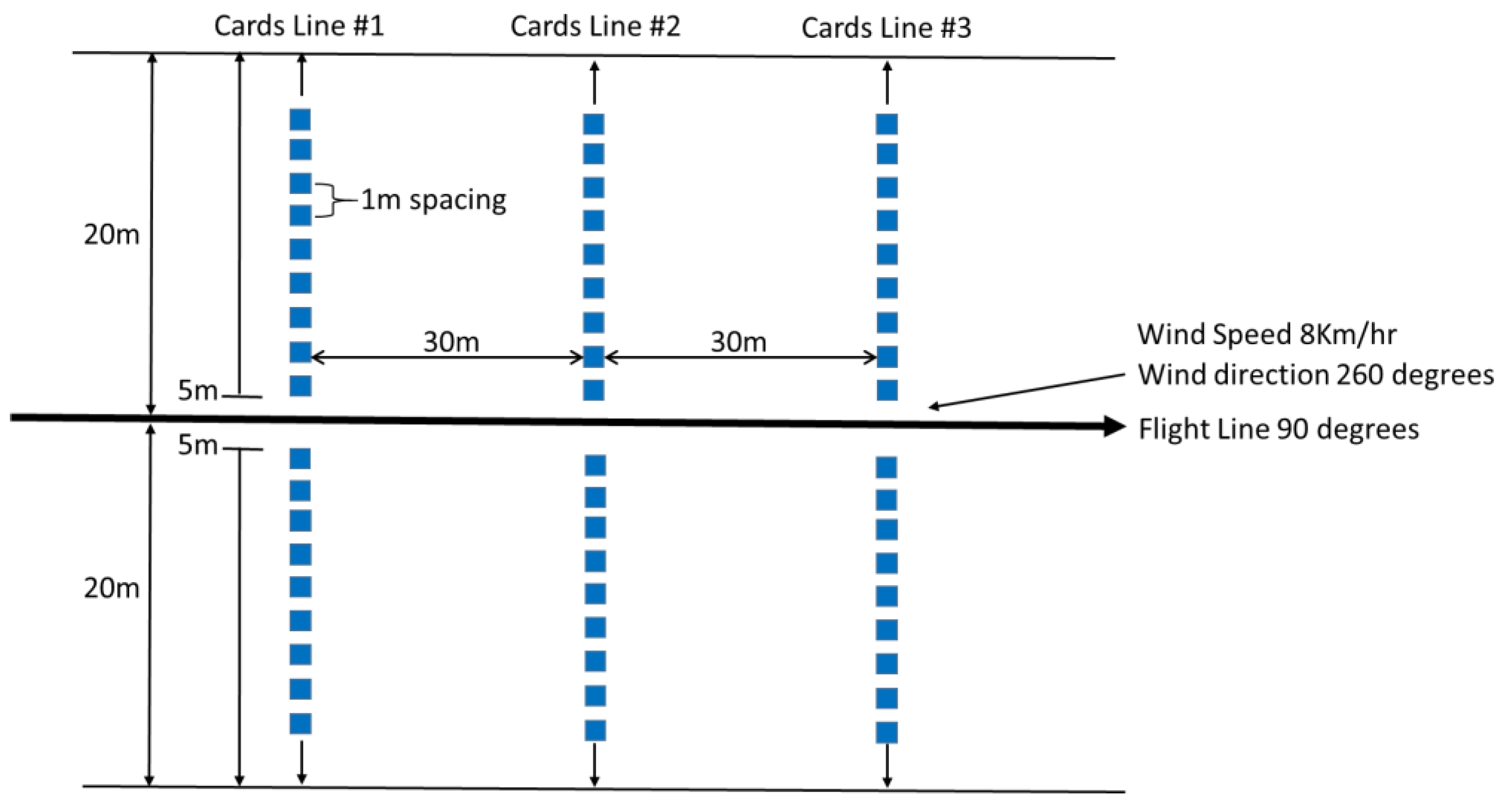
2.4. Pheromone Application
2.5. Statistical Analysis
3. Results
3.1. Spray Deposit
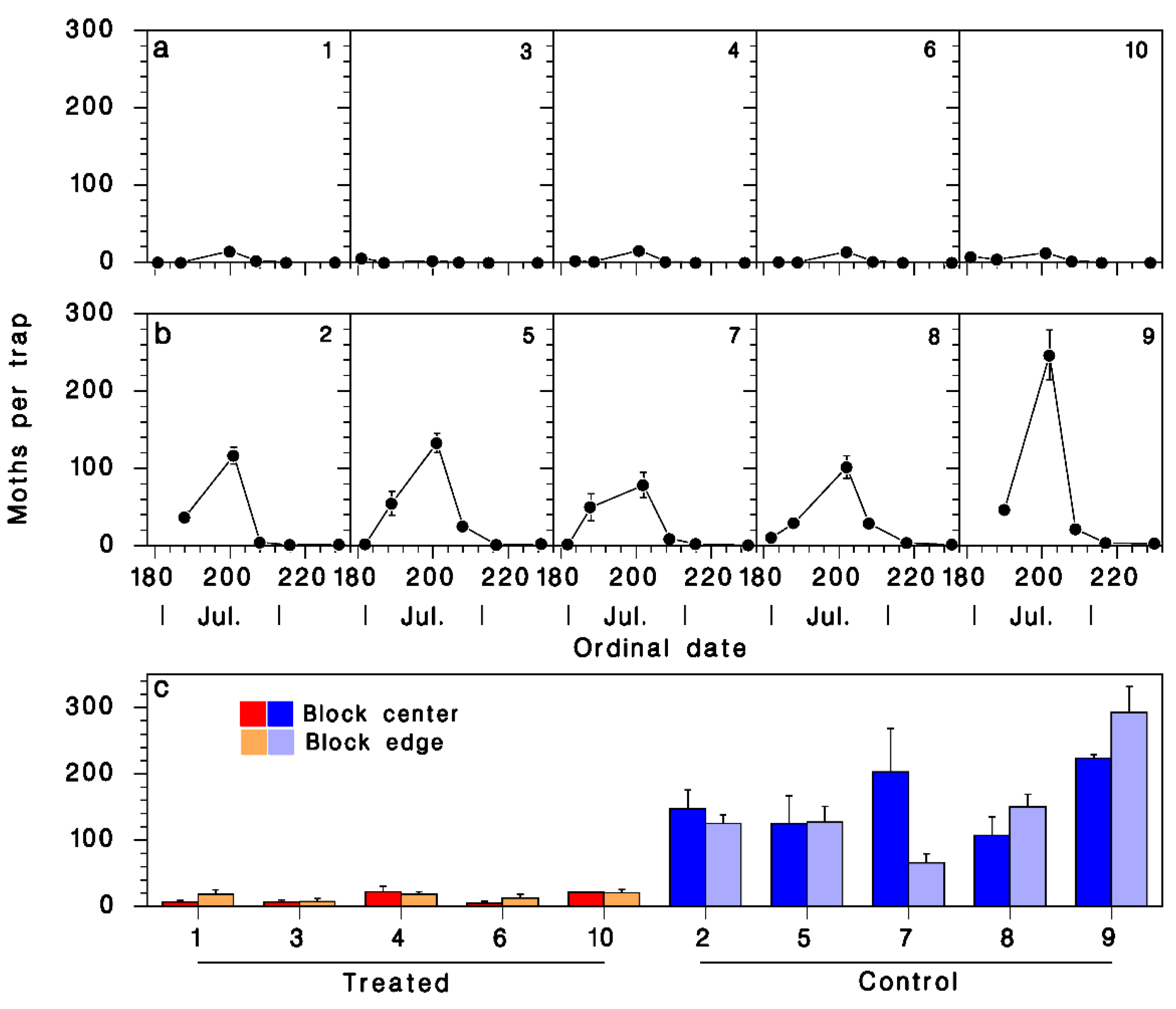
3.2. Captures in Pheromone Traps
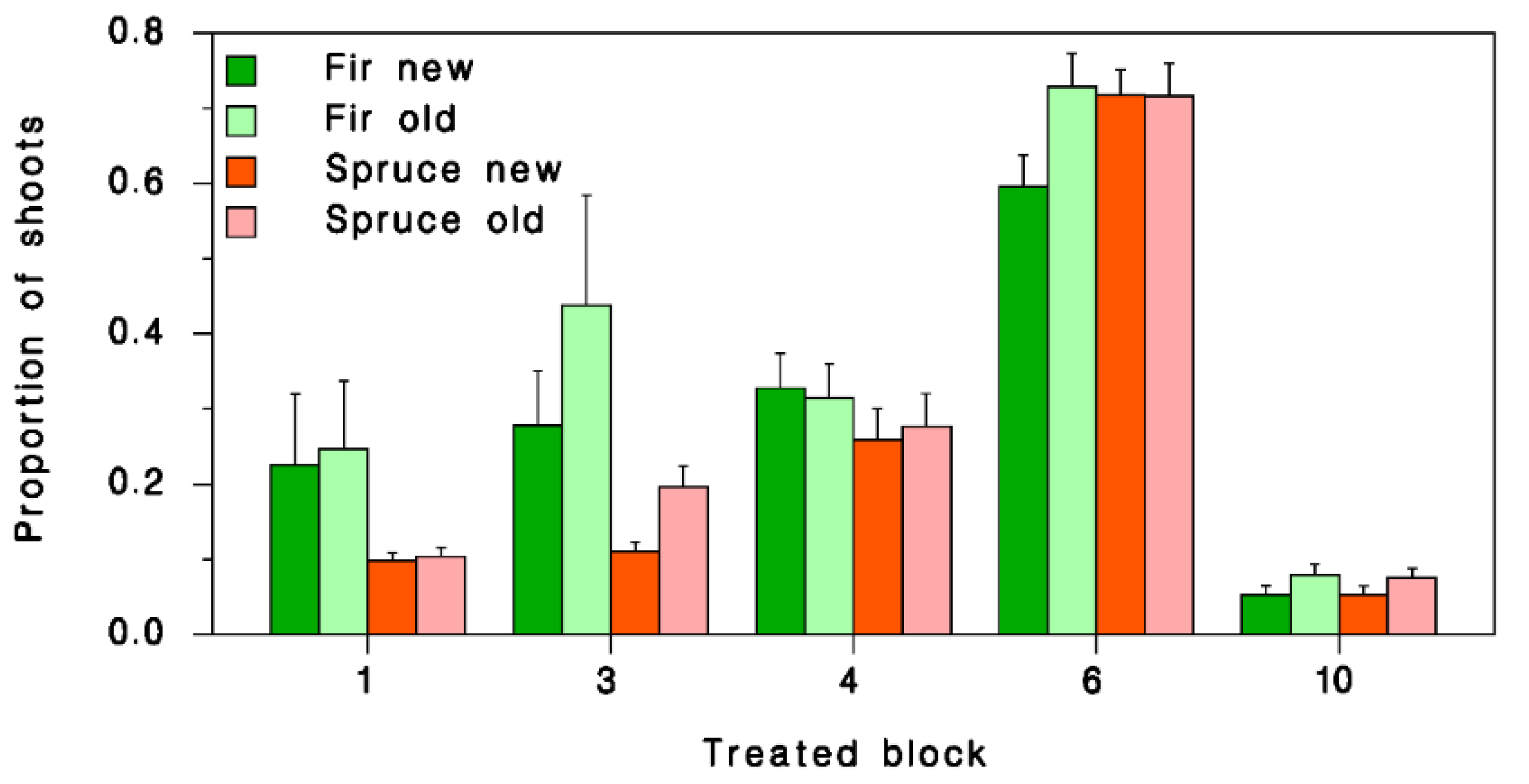
3.3. Pheromone Treatment Efficacy on SBW Population Performance
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greenfield, M.D. Moth sex pheromones: An evolutionary perspective. Fla. Entomol. 1981, 64, 4–17. [Google Scholar] [CrossRef]
- Cardé, R.T.; Haynes, K.F. Structure of the pheromone communication. In Advances in Insect Chemical Ecology; Cardé, R.T., Minks, J.G., Eds.; Cambridge University Press: Cambridge MA, USA, 2004; pp. 283–332. [Google Scholar]
- Byers, J.A. Pheromone component patterns of moth evolution revealed by computer analysis of the Pherolist. J. Anim. Ecol. 2006, 75, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, T.C.; Kuenen, L.P.S. Pheromone source location by flying moths: A supplementary non-anemotactic mechanism. Science 1982, 216, 424–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, J.S. Zigzagging and casting as a programmed response to wind-borne odour: A review. Physiol. Entomol. 1983, 8, 109–120. [Google Scholar] [CrossRef]
- Mafra-Neto, A.; Cardé, R.T. Fine-scale structure of pheromone plumes modulates upwind orientation of flying moths. Nature 1994, 369, 142–144. [Google Scholar] [CrossRef]
- Cork, A. Pheromones as management tools: Mass-trapping, and lure and kill. In Pheromone Communication in Moths: Evolution, Behavior, and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016; pp. 349–365. [Google Scholar]
- Evenden, M. Mating disruption of moth pests in integrated pest management. In Pheromone Communication in Moths: Evolution, Behavior, and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016; pp. 365–393. [Google Scholar]
- Suckling, D.M. Monitoring for surveillance and management. In Pheromone Communication in Moths: Evolution, Behavior, and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016; pp. 337–347. [Google Scholar]
- Foster, S.P.; Harris, M.O. Behavioral manipulation methods for insect pest-management. Annu. Rev. Entomol. 1997, 42, 123–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhainds, M.; Kettela, E.G.; Silk, P.J. CP Alexander review: Thirty-five years of pheromone-based mating disruption studies with Choristoneura fumiferana (Clemens) (Lepidoptera: Tortricidae). Can. Entomol. 2012, 144, 379–395. [Google Scholar] [CrossRef]
- Cardé, R.T.; Minks, A.K. Control of moth pests by mating disruption: Successes and constraints. Annu. Rev. Entomol. 1995, 40, 559–585. [Google Scholar] [CrossRef]
- Bartell, R.J. Mechanisms of communication disruption by pheromone in the control of Lepidoptera: A review. Physiol. Entomol. 1982, 7, 353–364. [Google Scholar] [CrossRef]
- Gut, L.J.; Stelinski, L.L.; Thomson, D.R.; Miller, J.R.; Koul, O.; Dhaliwal, G.S.; Cuperus, G.W. Behaviour-Modifying Chemicals: Prospects and Constraints in IPM. Integrated Pest Management Potential, Constraints and Challenges; CABI Publishing: Cambridge, MA, USA, 2004. [Google Scholar]
- Baker, T.C.; Heath, J.J. Pheromones: Function and use in insect control. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Pergamon Press: Berkeley, CA, USA; Elsevier: Amsterdam, The Netherlands, 2005; Volume 6, pp. 407–459. [Google Scholar]
- Jacobson, M. Insect Sex Attractants; John Wiley & Sons: New York, NY, USA; London, UK; Sydney, Australia, 1965. [Google Scholar]
- Gaston, L.K.; Shorey, H.H.; Saario, C.A. Insect population control by the use of sex pheromones to inhibit orientation between the sexes. Nature 1967, 213, 155–1155. [Google Scholar] [CrossRef]
- Gaston, L.K.; Kaae, R.S.; Shorey, H.H.; Sellers, D. Controlling the pink bollworm by disrupting sex pheromone communication between adult moths. Science 1977, 196, 904–905. [Google Scholar] [CrossRef] [PubMed]
- Critchley, B.R.; Campion, D.G.; McVeigh, L.J.; Hunter-Jones, P.; Hall, D.R.; Cork, A.R.; Nesbitt, B.F.; Marrs, G.J.; Jutsum, A.R.; Hosny, M.M.; et al. Control of pink boll worm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae), in Egypt by mating disruption using an aerially applied microencapsulated pheromone formulation. Bull. Entomol. Res. 1983, 73, 289–299. [Google Scholar] [CrossRef]
- Staten, R.T.; Flint, H.M.; Weddle, R.C.; Quintero, E.; Zarate, R.E.; Finnell, C.M.; Hernandes, M.; Yamamoto, A. Pink bollworm (Lepidoptera: Gelechiidae): Large-scale field trials with a high-rate gossyplure formulation. J. Econ. Entomol. 1987, 80, 1267–1271. [Google Scholar] [CrossRef]
- Cardé, R.T.; Baker, T.C.; Castrovillo, P.J. Disruption of sexual communication in Laspeyresia pomonella (codling moth), Grapholitha molesta (Oriental fruit moth) and G. prunivora (lesser appleworm) with hollow fiber attractant sources. Entomol. Exp. Appl. 1977, 22, 280–288. [Google Scholar] [CrossRef]
- Charlton, R.E.; Cardé, R.T. Comparing the effectiveness of sexual communication disruption in the oriental fruit moth (Grapholitha molesta) using different combinations and dosages of its pheromone blend. J. Chem. Ecol. 1981, 7, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Vickers, R.A.; Rothschild, G.H.L.; Jones, E.L. Control of the oriental fruit moth, Cydia molesta (Busek) (Lepidoptera: Tortricidae), at a district level by mating disruption with synthetic female pheromone. Bull. Entomol. Res. 1985, 75, 625–634. [Google Scholar] [CrossRef]
- Moffitt, H.R.; Westigard, P.H. Suppression of the codling moth (Lepidoptera: Tortricidae) population on pears in southern Oregon through mating disruption with sex pheromone. J. Econ. Entomol. 1984, 77, 1513–1519. [Google Scholar] [CrossRef]
- Jiménez, M.; Toscano, N.; Flaherty, D.; Ilic, P.; Zalom, F.; Kido, K. Controlling tomato pinworm by mating disruption. Calif. Agric. 1988, 42, 10–12. [Google Scholar]
- Henneberry, T.J.; Gillespie, J.M.; Bariola, L.A.; Flin, H.M.; Lingren, P.D.; Kydonieus, A.F. Gossyplure in laminated plastic formulations for mating disruption and pink bollworm control. J. Econ. Entomol. 1981, 74, 376–381. [Google Scholar] [CrossRef]
- Westigard, P.H.; Moffitt, H.R. Natural control of the pear psylla (Homoptera: Psyllidae): Impact of mating disruption with the sex pheromone for control of the codling moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 1984, 77, 1520–1523. [Google Scholar] [CrossRef]
- Dennehy, T.; Clark, L.; Kamas, J. Pheromonal control of the grape berry moth: An effective alternative to conventional insecticides. N.Y. Food Life Sci. Bull. 1991, 135, 1–6. [Google Scholar]
- Royama, T. Population dynamics of the spruce budworm Choristoneura fumiferana. Ecol. Monog. 1984, 54, 429–462. [Google Scholar] [CrossRef]
- Weatherston, J.; Roelofs, W.; Comeau, A.; Sanders, C.J. Studies of the physiologically active arthropod secretions: X. Sex pheromone of the eastern spruce budworm, Choristoneura fumiferana (Lepidoptera: Tortricidae). Can. Entomol. 1971, 103, 1741–1747. [Google Scholar] [CrossRef] [Green Version]
- Silk, P.J.; Tan, S.H.; Wiesner, C.J.; Ross, R.J.; Lonergan, G.C. Sex pheromone chemistry of the eastern spruce budworm, Choristoneura fumiferana. Environ. Entomol. 1980, 9, 640–644. [Google Scholar] [CrossRef]
- Silk, P.J.; Kuenen, L.P.S. Sex pheromones and behavioral biology of the coniferophagous Choristoneura. Annu. Rev. Entomol. 1988, 33, 83–101. [Google Scholar] [CrossRef]
- Kettela, E.; Silk, P. Development of a Pheromone Formulation for Use in Early Intervention Pest Management Strategies of the Spruce Budworm Choristoneura fumiferana (Clem.). Final Report, SERG-1 Project. 2005, Volume 614. Available online: https://documents.serginternational.org/english/index.html (accessed on 14 November 2022).
- Régnière, J.; Delisle, J.; Dupont, A.; Trudel, R. The impact of moth migration on apparent fecundity overwhelms mating disruption as a method to manage spruce budworm populations. Forests 2019, 10, 775. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, K.W.; Van der Pers, J.; Leonard, D.S.; Sellers, P.; Mastro, V.C.; Webb, R.E.; Reardon, R.C. Electroantennogram measurements of atmospheric pheromone concentration after aerial and ground application of gypsy moth mating disruptants. J. Appl. Entomol. 2007, 131, 146–152. [Google Scholar] [CrossRef]
- MacLean, D.; Amirault, P.; Amos-Binks, L.; Cerleton, D.; Hennigar, C.; Johns, R.; Régnière, J. Positive results of an early intervention strategy to suppress a spruce budworm outbreak after five years of trials. Forests 2019, 10, 448. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.A.; Kettela, E.G.; McDougall, G.A. A Sampling Technique for Overwintering Spruce Budworm and Its Applicability to Population Surveys. Canadian Forestry Service, Info. Rep. M-X-25. 1971, pp. 1–11. Available online: https://cfs.nrcan.gc.ca/publications/download-pdf/7648 (accessed on 14 November 2022).
- Régnière, J.; Cooke, B.J.; Béchard, A.; Dupont, A.; Therrien, P. Dynamics and management of rising outbreak spruce budworm populations. Forests 2019, 10, 748. [Google Scholar] [CrossRef] [Green Version]
- Régnière, J.; Nealis, V.G. Density dependence of egg recruitment and moth dispersal in spruce budworms. Forests 2019, 10, 706. [Google Scholar] [CrossRef] [Green Version]
- Fettes, J.J. Investigation of Sampling Techniques for Population Studies of the Spruce Budworm on Balsam Fir in Ontario; Annual Technical Report; Forest Insect Laboratory: Sault Ste. Marie, ON, Canada, 1950; pp. 163–401. Available online: https://www.yumpu.com/en/document/read/35161436/forest-insect-laboratory-sault-ste-marie-ontario-annual-nfis (accessed on 9 November 2022).
- Sanders, C.J. Evaluation of high-capacity, nonsaturating sex pheromone traps for monitoring population densities of spruce budworm (Lepidoptera: Tortricidae). Can. Entomol. 1986, 118, 611–619. [Google Scholar] [CrossRef]
- Jobin, L.J.; Coulombe, C.; Bernier-Cardou, M. Use of the Multi-Pher® Trap to Monitor Spruce Budworm Populations. Canadian Forest Service, Info. Rep. LAU-X-103E. 1993. Available online: https://scf.rncan.gc.ca/publications/telecharger-pdf/15525 (accessed on 14 November 2022).
- Régnière, J.; Delisle, J.; Pureswaran, D.; Trudel, R. Mate-finding Allee effect in spruce budworm population dynamics. Entomol. Exp. Appl. 2012, 146, 112–122. [Google Scholar] [CrossRef]
- Rhainds, M.; Kettela, E.G. Oviposition threshold for flight in an inter-reproductive migrant moth. J. Insect Behav. 2013, 26, 850–859. [Google Scholar] [CrossRef]
- Régnière, J.; Sanders, C.J. Optimal sample size for the estimation of spruce budworm (Lepidoptera: Tortricidae) populations on balsam fir and white spruce. Can. Entomol. 1983, 115, 1621–1626. [Google Scholar] [CrossRef] [Green Version]
- Witzgall, P.; Stelinski, L.; Gut, L.; Thomson, D. Codling moth management and chemical ecology. Annu. Rev. Entomol. 2008, 53, 503–522. [Google Scholar] [CrossRef] [Green Version]
- Morton, B.; Hood, C.S.; Trefrey, D.; Leonard, D.E.; Knipling, E.F.; Klassen, W.; Stevens, L.J. Large field trial with microencapsulated sex pheromone to prevent mating of the gypsy moth. J. Econ. Entomol. 1974, 67, 659–664. [Google Scholar] [CrossRef]
- Onufrieva, K.S.; Hickman, A.D.; Leonard, D.S.; Tobin, P.C. Relationship between efficacy of mating disruption and gypsy moth density. Int. J. Pest Manag. 2019, 65, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Régnière, J.; Delisle, J.; Sturtevant, B.R.; Garcia, M.; Saint-Amant, R. Modeling migratory flight in the spruce budworm: Temperature constraints. Forests 2019, 10, 802. [Google Scholar] [CrossRef] [Green Version]
- Rhainds, M.; Lavigne, D.; Boulanger, Y.; DeMerchant, I.; Delisle, J.; Motty, J.; Rideout, T.; Labrecque, A. I know it when I see it: Incidence and intensity of immigration in spruce budworm. Agric. For. Entomol. 2022, 24, 152–166. [Google Scholar] [CrossRef]
- Brunner, J.; Welter, S.; Calkins, C.; Hilton, R.; Beers, E.; Dunley, J.; Unruh, T.; Knight, A.; Van Steenwyk, R.; Van Buskirk, P. Mating disruption of codling moth: A perspective from the Western United States. IOBC Wprs. Bull. 2002, 25, 11–20. Available online: https://www.researchgate.net/publication/242131238_Mating_disruption_of_codling_moth_A_perspective_from_the_Western_United_States (accessed on 14 November 2022).
- Sanders, C.J.; Lucuik, G.S. Mating behavior of spruce budworm moths, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). Can. Entomol. 1992, 124, 273–286. [Google Scholar] [CrossRef]
- Silk, P.J.; Eveleigh, E.; Roscoe, L.; Burgess, K.; Weatherby, S.; Leclair, G.; Mayo, P.; Brophy, M. Unsaturated cuticular hydrocarbons enhance responses to sex pheromone in spruce budworm, Choristoneura fumiferana. J. Chem. Ecol. 2017, 43, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Ioriatti, C.; Bagnoli, B.; Lucchi, A.; Veronelli, V. Vine moths control by mating disruption in Italy: Results and future prospects. Redia 2004, 87, 117–128. Available online: http://hdl.handle.net/10449/17868 (accessed on 14 November 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Paired Comparisons | Odds Ratio | 95% Confidence Interval | ||
|---|---|---|---|---|
| Lower | Upper | |||
| Blocks | ||||
| 3 | 1 | 0.9769 | 0.8863 | 1.0769 |
| 4 | 1 | 4.4844 | 4.1533 | 4.8420 |
| 6 | 1 | 15.863 | 14.433 | 17.436 |
| 10 | 1 | 0.4973 | 0.4484 | 0.5515 |
| 4 | 3 | 4.5903 | 4.1950 | 5.0228 |
| 6 | 3 | 16.238 | 14.612 | 18.045 |
| 10 | 3 | 0.5090 | 0.4542 | 0.5705 |
| 6 | 4 | 3.5375 | 3.2533 | 3.8465 |
| 10 | 4 | 0.1109 | 0.1008 | 0.1220 |
| 10 | 6 | 0.0313 | 0.0281 | 0.0350 |
| Host species | ||||
| Spruce | Fir | 0.8566 | 0.7942 | 0.9238 |
| Shoot age | ||||
| Old | New | 1.1857 | 1.1204 | 1.2548 |
| Source | DF | Adj SS | Adj MS | F | p > F |
|---|---|---|---|---|---|
| Treatment | 1 | 363.1 | 363.1 | 128.70 | 0.000 |
| Trap position (Center/Edge) | 1 | 0.705 | 0.705 | 0.25 | 0.621 |
| Block (Treatment) | 8 | 33.57 | 4.196 | 1.49 | 0.203 |
| Treatment × Position | 1 | 0.308 | 0.308 | 0.11 | 0.743 |
| Block (Treatment) × Position | 8 | 16.45 | 2.056 | 0.73 | 0.665 |
| Error | 30 | 84.65 | 2.822 | ||
| Total | 49 | 723.7 |
| Source | DF | Adj SS | Adj MS | F | p > F |
|---|---|---|---|---|---|
| log(E) regression | 3 | 1.18910 | 0.39637 | 7.75 | 0.017 |
| Treatment | 1 | 0.05678 | 0.05678 | 1.11 | 0.332 |
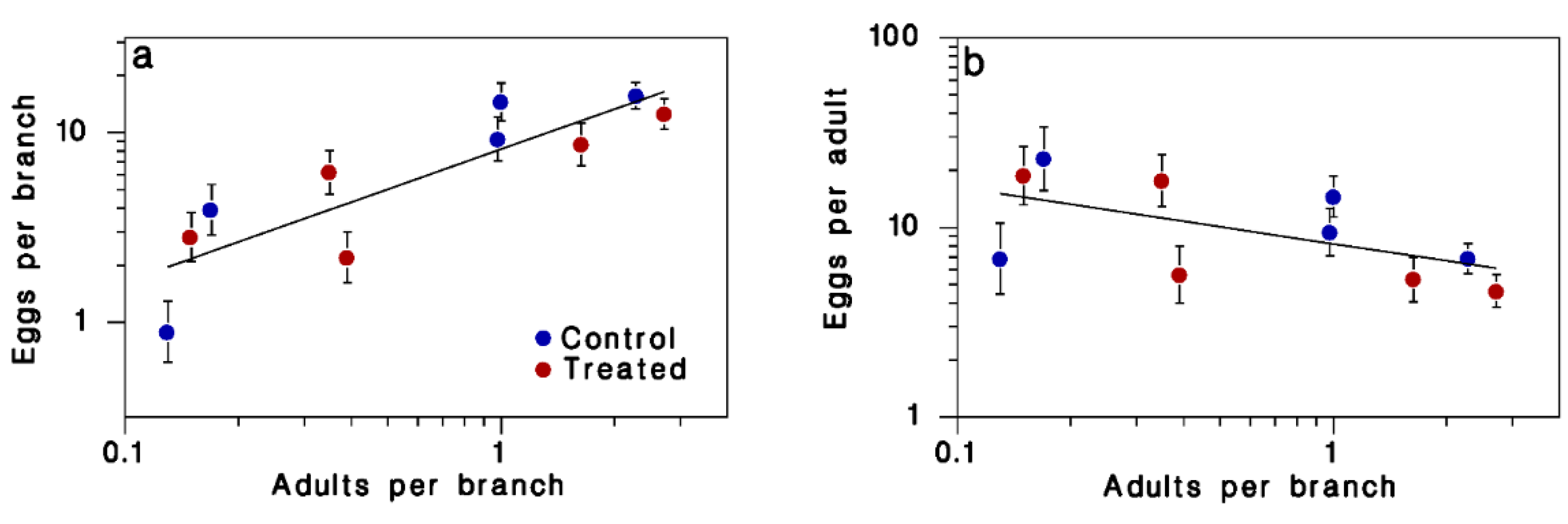
| log(A) | 1 | 0.89038 | 0.89038 | 17.42 | 0.006 |
| log(A) × Treatment | 1 | 0.06872 | 0.06872 | 1.34 | 0.290 |
| Error | 6 | 0.30669 | 0.05111 | ||
| Total | 9 | 1.49579 | |||
| log(E/A) regression | 3 | 0.28694 | 0.09565 | 1.87 | 0.235 |
| Treatment | 1 | 0.05678 | 0.05678 | 1.11 | 0.332 |
| log(A) | 1 | 0.01832 | 0.01832 | 0.36 | 0.571 |
| log(A) × Treatment | 1 | 0.06872 | 0.06872 | 1.34 | 0.290 |
| Error | 6 | 0.30669 | 0.05111 | ||
| Total | 9 | 0.59363 |
| Source | DF | Adj SS | Adj MS | F | p > F |
|---|---|---|---|---|---|
| log(L2f) regression | 3 | 2.85312 | 0.95104 | 5.45 | 0.038 |
| Treatment | 1 | 0.01688 | 0.01688 | 0.10 | 0.766 |
| log(L2s) | 1 | 1.93502 | 1.93502 | 11.09 | 0.016 |
| log(L2s) × Treatment | 1 | 0.00776 | 0.00776 | 0.04 | 0.840 |
| Error | 6 | 1.04665 | 0.17444 | ||
| Total | 9 | 3.89976 | |||
| log (L2f/L2s) regression | 3 | 0.55897 | 0.186322 | 1.07 | 0.430 |
| Treatment | 1 | 0.01688 | 0.016882 | 0.10 | 0.766 |
| log(L2s) | 1 | 0.38808 | 0.388079 | 2.22 | 0.186 |
| log(L2s) × Treatment | 1 | 0.00776 | 0.007757 | 0.04 | 0.840 |
| Error | 6 | 1.04665 | 0.174442 | ||
| Total | 9 | 1.60562 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roscoe, L.E.; MacKinnon, W.; Régnière, J.; Forbes, G.; Brophy, M.; Lamb, R. Use of a Sprayable Sex Pheromone Formulation in Landscape-Level Control of Choristoneura fumiferana Populations. Insects 2022, 13, 1175. https://doi.org/10.3390/insects13121175
Roscoe LE, MacKinnon W, Régnière J, Forbes G, Brophy M, Lamb R. Use of a Sprayable Sex Pheromone Formulation in Landscape-Level Control of Choristoneura fumiferana Populations. Insects. 2022; 13(12):1175. https://doi.org/10.3390/insects13121175
Chicago/Turabian StyleRoscoe, Lucas E., Wayne MacKinnon, Jacques Régnière, Glen Forbes, Matt Brophy, and Rosanna Lamb. 2022. "Use of a Sprayable Sex Pheromone Formulation in Landscape-Level Control of Choristoneura fumiferana Populations" Insects 13, no. 12: 1175. https://doi.org/10.3390/insects13121175
APA StyleRoscoe, L. E., MacKinnon, W., Régnière, J., Forbes, G., Brophy, M., & Lamb, R. (2022). Use of a Sprayable Sex Pheromone Formulation in Landscape-Level Control of Choristoneura fumiferana Populations. Insects, 13(12), 1175. https://doi.org/10.3390/insects13121175