

Fermented or Floral? Developing a Generalized Food Bait Lure to Monitor Cutworm and Armyworm Moths (Lepidoptera: Noctuidae) in Field Crops



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Lures
2.3. Monitoring and Moth Identification
2.4. Experiment 1—Efficacy of AAMB Lures to Monitor Cutworm and Armyworm Moths
2.5. Experiment 2—Attractiveness of AAMB Lures at Different Release Rates
2.6. Experiment 3—Attractiveness of AAMB Lure Released from Different Devices
2.7. Experiment 4—Augmentation of AAMB Lures with Additional Food-Based Semiochemicals
2.8. Experiment 5—Electrophysiological Response of Redbacked Cutworm Moths to Food Bait Volatiles
2.9. Statistical Analyses
3. Results
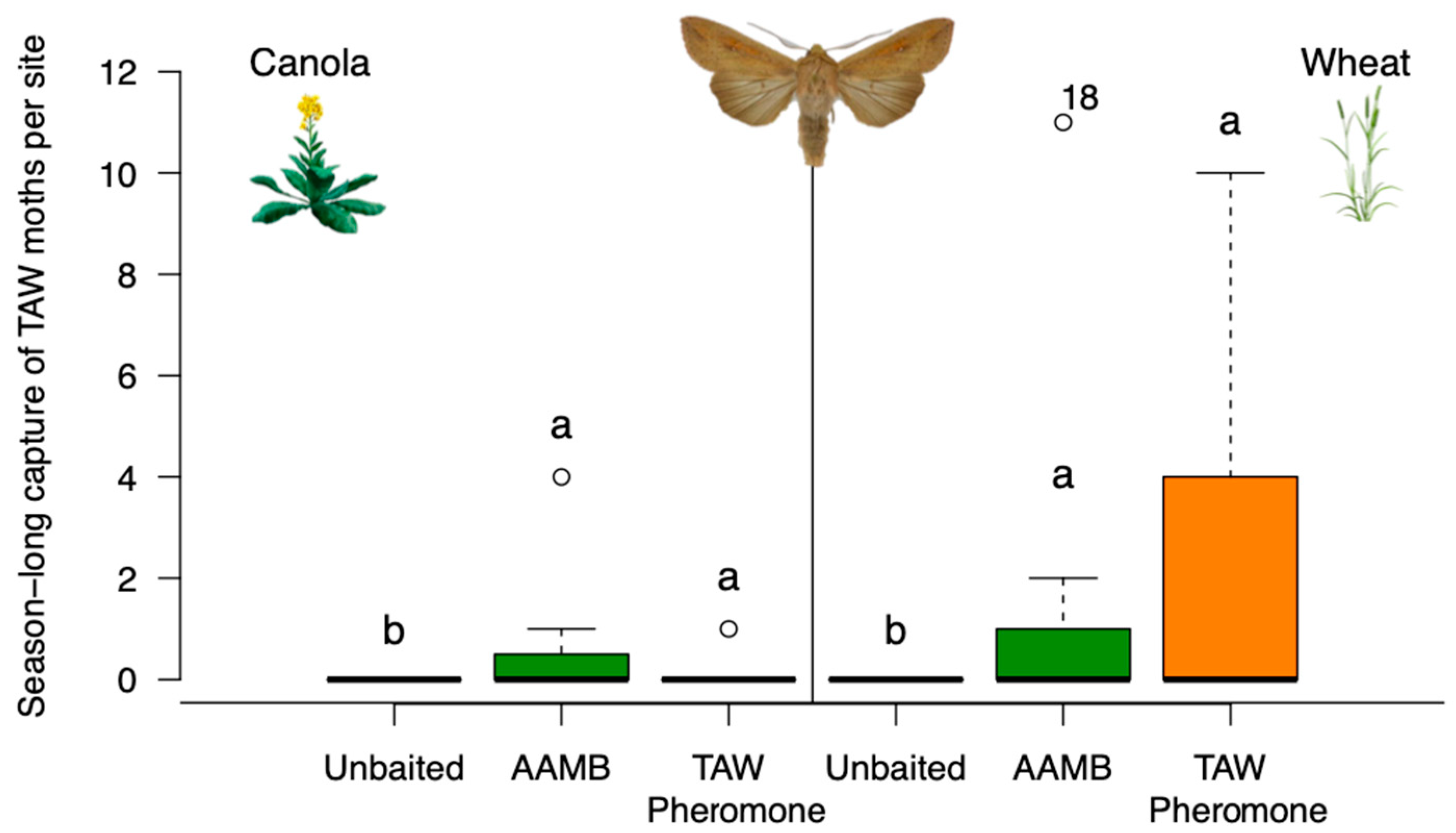
3.1. Experiment 1—Efficacy of AAMB Lures to Monitor Cutworm and Armyworm Moths
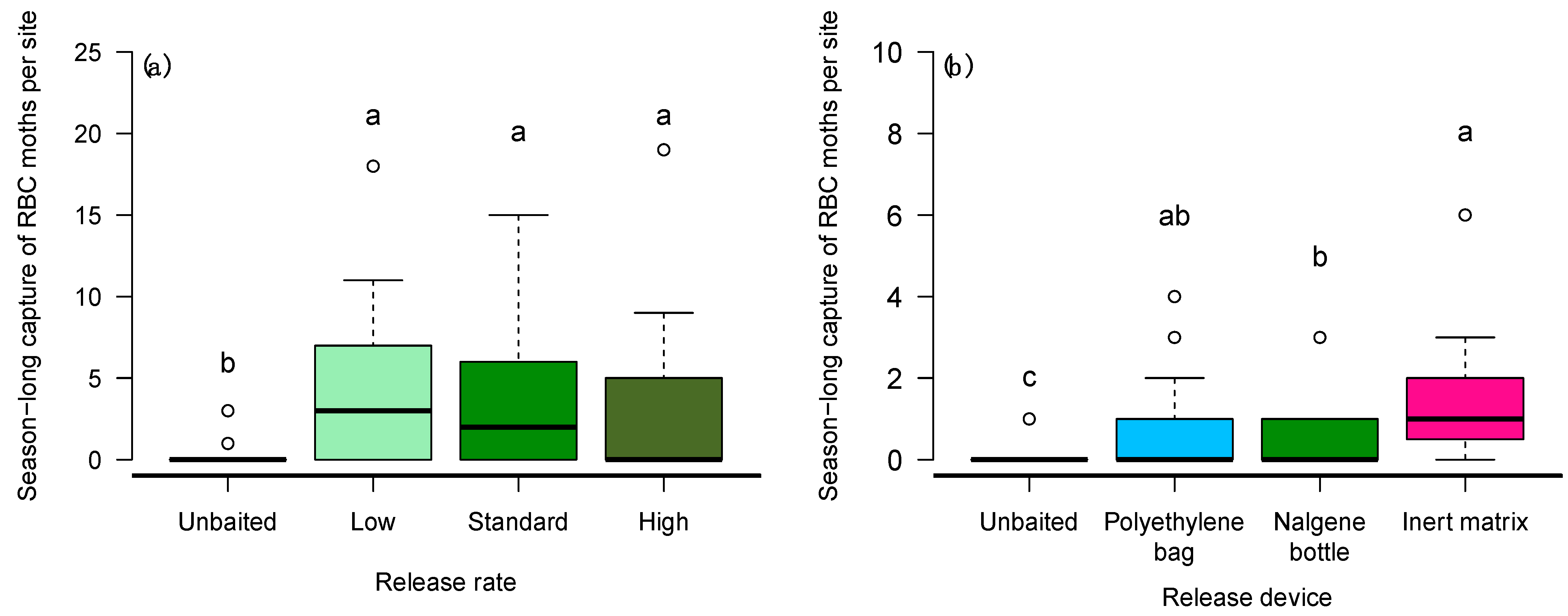
3.2. Experiment 2—Attractiveness of AAMB Lures at Different Release Rates
3.3. Experiment 3—Attractiveness of AAMB Lure Released from Different Devices
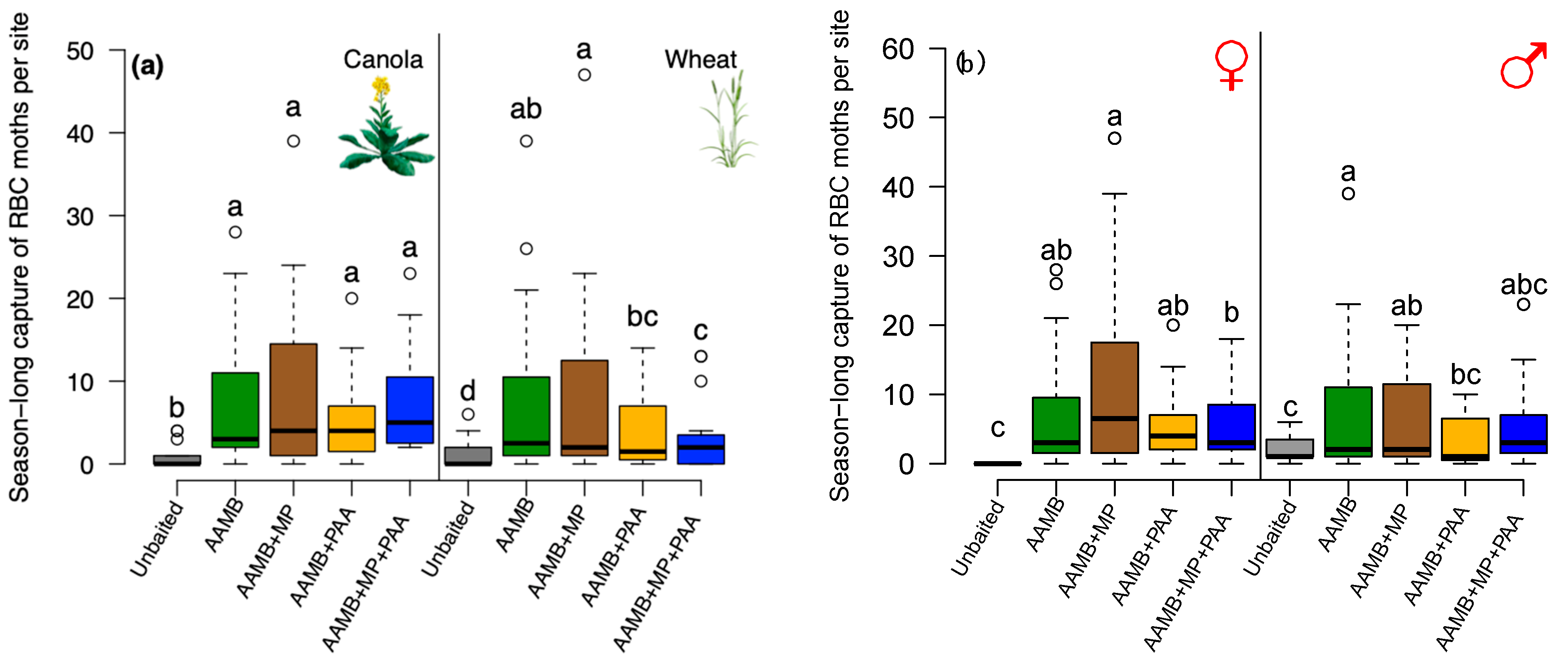
3.4. Experiment 4—Augmentation of AAMB Lures with Additional Food-Based Semiochemicals
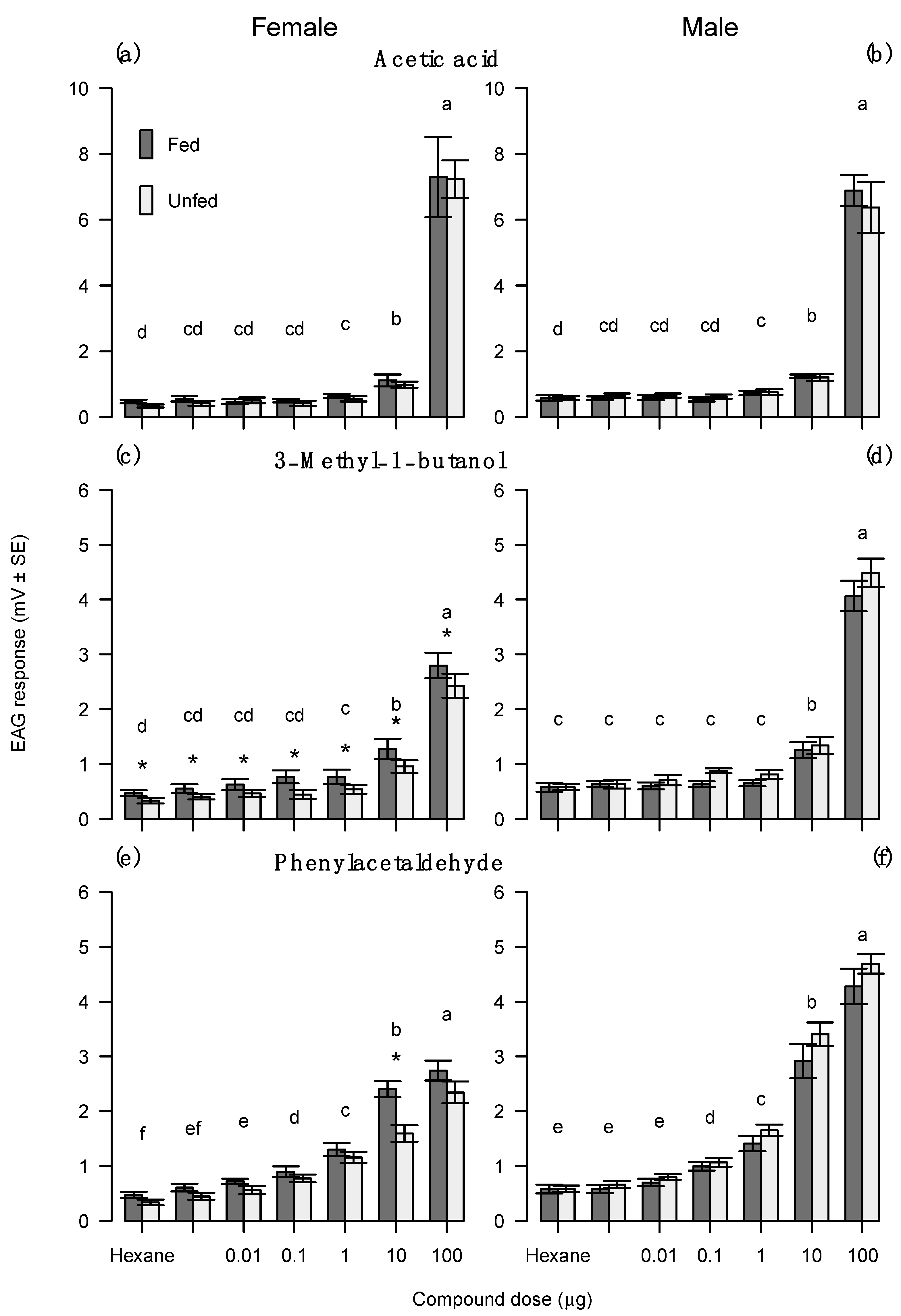
3.5. Experiment 5—Electrophysiological Response of Redbacked Cutworm to Food Bait Volatiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Floate, K.D. Cutworm Pest of Crops on the Canadian Prairie: Indentification and Management Field Guide; Agriculture and Agri-Food: Lethbridge, AB, Canada, 2017; p. 94. [Google Scholar]
- Beirne, B.P. Pest insects of annual crop plants in Canada: Part I, Lepidoptera; II, Diptera; III, Coleoptera. Mem. Entomol. Soc. Can. 1971, 103, 1–124. [Google Scholar] [CrossRef]
- Breeland, S.G. Biological studies on the armyworm, Pseudaletia unipuncta (Haworth), in Tennessee (Lepidoptera: Noctuidae). J. Tenn. Acad. 1958, 33, 263–350. [Google Scholar]
- Mason, P.; Arthur, A.; Olfert, O.; Erlandson, M. The bertha armyworm (Mamestra configurata) (Lepidoptera: Noctuidae) in Western Canada. Can. Entomol. 1998, 130, 321–336. [Google Scholar] [CrossRef]
- Hendrix, W.H., III; Showers, W.B. Tracing black cutworm and armyworm (Lepidoptera: Noctuidae) northward migration using Pithecellobium and Calliandra pollen. Environ. Entomol. 1992, 21, 1092–1096. [Google Scholar] [CrossRef]
- McNeil, J.N. The true armyworm, Pseudaletia unipuncta: A victim of the pied piper or a seasonal migrant? Int. J. Trop. Insect Sci. 1987, 8, 591–597. [Google Scholar] [CrossRef]
- Showers, W.B.; Whitford, F.; Smelser, R.B.; Keaster, A.J.; Robinson, J.F.; Lopez, J.D.; Taylor, S.E. Direct evidence for meteorologically driven long-range dispersal of an economically important moth. Ecology 1989, 70, 987–992. [Google Scholar] [CrossRef]
- Byers, J.; Yu, D.; Jones, J. Parasitism of the army cutworm, Euxoa auxiliaris (Grt.) (Lepidoptera: Noctuidae), by Copidosoma bakeri (Howard) (Hymenoptera: Encyrtidae) and effect on crop damage. Can. Entomol. 1993, 125, 329–335. [Google Scholar] [CrossRef]
- Guppy, J.C. Life history and behaviour of the armyworm, Pseudaletia unipuncta (Haw.) (Lepidoptera: Noctuidae), in eastern Ontario. Can. Entomol. 1961, 93, 1141–1153. [Google Scholar] [CrossRef]
- Dosdall, L.M.; Otani, J.; Byers, R.J.; Seward, D.; Yoder, C.; Huffman, J.; Guitard, R.; Goudreau, H. The Outbreak Of Glassy Cutworm (Apamea devastator) (Lepidoptera: Noctuidae) in Alberta, 2000. In Pest Management Research Report—2000 Growing Season: Compiled for the Expert Committee on Integrated Pest Management; Agriculture and Agri-Food Canada: London, ON, Canada, 2001. [Google Scholar]
- W.C.C.P. Minutes of the Western Forum for Pest Management Annual Meeting. In Proceedings of the Western Forum for Pest Management Annual Meeting, Sandman Hotel, Abbotsford, BC, Canada, 22 October 2015. [Google Scholar]
- W.C.C.P. Minutes of the Western Forum for Pest Management Annual Meeting. In Proceedings of the Western Forum for Pest Management Annual Meeting, Wanuskewin Heritage Park, Saskatoon, SK, Canada, 20 October 2016. [Google Scholar]
- Ayre, G.L.; Lamb, R.J. Life histories, flight patterns, and relative abundance of 9 cutworms (Lepidoptera: Noctuidae) in Manitoba. Can. Entomol. 1990, 122, 1059–1070. [Google Scholar] [CrossRef]
- Kogan, M. Integrated Pest Management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef]
- Steck, W.; Underhill, E.; Chisholm, M. Structure-activity relationships in sex attractants for North American noctuid moths. J. Chem. Ecol. 1982, 8, 731–754. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.R.; Struble, D.L. Monitoring population levels of 8 species of noctuids with sex attractant traps in Southern Alberta, 1978–1983: Specificity of attractants and effect of target species abundance. Can. Entomol. 1987, 119, 541–556. [Google Scholar] [CrossRef]
- Grocock, N.L.; Batallas, R.E.; McNamara, E.A.; Sturm, A.B.; Manson, J.S.; Evenden, M.L. Bumble bees (Hymenoptera: Apidae) respond to moth (Lepidoptera: Noctuidae) pheromone components, leading to bee bycatch in monitoring traps targeting moth pests. Front. Ecol. Evol. 2020, 8, 576692. [Google Scholar] [CrossRef]
- Gross, H.; Carpenter, J. Role of the fall armyworm (Lepidoptera: Noctuidae) pheromone and other factors in the capture of bumblebees (Hymenoptera: Aphidae) by universal moth traps. Environ. Entomol. 1991, 20, 377–381. [Google Scholar] [CrossRef]
- Mori, B.A.; Evenden, M.L. Factors affecting pheromone-baited trap capture of male Coleophora deauratella, an invasive pest of clover in Canada. J. Econ. Entomol. 2013, 106, 844–854. [Google Scholar] [CrossRef]
- Spears, L.R.; Looney, C.; Ikerd, H.; Koch, J.B.; Griswold, T.; Strange, J.P.; Ramirez, R.A. Pheromone lure and trap color affects bycatch in agricultural landscapes of Utah. Environ. Entomol. 2016, 45, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.S.; Landolt, P.J. A survey of insect assemblages responding to volatiles from a ubiquitous fungus in an agricultural landscape. J. Chem. Ecol. 2013, 39, 860–868. [Google Scholar] [CrossRef]
- Visser, J.H. Host-plant finding by insects—Orientation, sensory input and search patterns. J. Insect Physiol. 1988, 34, 259–268. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. (Eds.) Insect-Plant Biology, 2nd ed.; Oxford University Press: Oxford, UK, 2005; p. 421. [Google Scholar]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial volatile emissions as insect semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Herrera, C.M.; García, I.M.; Pérez, R. Invisible floral larcenies: Microbial communities degrade floral nectar of bumble bee-pollinated plants. Ecology 2008, 89, 2369–2376. [Google Scholar] [CrossRef] [Green Version]
- Landolt, P.J.; Tóth, M.; Meagher, R.L.; Szarukán, I. Interaction of acetic acid and phenylacetaldehyde as attractants for trapping pest species of moths (Lepidoptera: Noctuidae). Pest Manage. Sci. 2013, 69, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Joyce, R.J.; Lingren, P.D. Potential for development of technology to control adult noctuid pests with plant attractants: Background and world perspective. Southwest. Entomol. 1998, 21, 9–24. [Google Scholar]
- Gregg, P.C.; Del Socorro, A.P.; Henderson, G.S. Development of a synthetic plant volatile-based attracticide for female noctuid moths. II. Bioassays of synthetic plant volatiles as attractants for the adults of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Aust. J. Entomol. 2010, 49, 21–30. [Google Scholar] [CrossRef]
- Light, D.M.; Knight, A.L.; Henrick, C.A.; Rajapaska, D.; Lingren, B.; Dickens, J.C.; Reynolds, K.M.; Buttery, R.G.; Merrill, G.; Roitman, J. A pear-derived kairomone with pheromonal potency that attracts male and female codling moth, Cydia pomonella (L.). Naturwissenschaften 2001, 88, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Cantelo, W.; Jacobson, M. Phenylacetaldehyde attracts moths to bladder flower and to blacklight traps. Environ. Entomol. 1979, 8, 444–447. [Google Scholar] [CrossRef]
- Landolt, P.J.; Smithhisler, C.L. Characterization of the floral odor of Oregongrape: Possible feeding attractants for moths. Northwest Sci. 2003, 77, 81–86. [Google Scholar]
- Meagher, R.L., Jr. Collection of soybean looper and other noctuids in phenylacetaldehyde-baited field traps. Fla. Entomol. 2001, 84, 154–155. [Google Scholar] [CrossRef]
- Guédot, C.; Landolt, P.J.; Smithhisler, C.L. Odorants of the flowers of butterfly bush, Buddleja davidii, as possible attractants of pest species of moths. Fla. Entomol. 2008, 91, 576–582. [Google Scholar]
- Landolt, P.J.; Pantoja, A.; Hagerty, A.; Crabo, L.; Green, D. Moths trapped in Alaska with feeding attractant lures and the seasonal flight patterns of potential agricultural pests. Can. Entomol. 2007, 139, 278–291. [Google Scholar] [CrossRef]
- Meagher, R.L., Jr.; Mitchell, E.R. Nontarget Hymenoptera collected in pheromone- and synthetic floral volatile- baited traps. Environ. Entomol. 1999, 28, 367–371. [Google Scholar] [CrossRef]
- Utrio, P.; Eriksson, K. Volatile fermentation products as attractants for Macrolepidoptera. Ann. Zool. Fenn. 1977, 14, 98–104. [Google Scholar]
- El-Sayed, A.; Heppelthwaite, V.; Manning, L.; Gibb, A.; Suckling, D. Volatile constituents of fermented sugar baits and their attraction to lepidopteran species. J. Agric. Food Chem. 2005, 53, 953–958. [Google Scholar] [CrossRef]
- Tóth, M.; Szarukán, I.; Dorogi, B.; Gulyás, A.; Nagy, P.; Rozgonyi, Z. Male and female noctuid moths attracted to synthetic lures in Europe. J. Chem. Ecol. 2010, 36, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J. New chemical attractants for trapping Lacanobia subjuncta, Mamestra configurata, and Xestia c-nigrum (Lepidoptera: Noctuidae). J. Econ. Entomol. 2000, 93, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J.; Higbee, B.S. Both sexes of the true armyworm (Lepidoptera: Noctuidae) trapped with the feeding attractant composed of acetic acid and 3-methyl-1-butanol. Fla. Entomol. 2002, 85, 182–185. [Google Scholar] [CrossRef]
- Lingren, P.D.; Raulston, J.R.; Shaver, T.N. Potential for development of technology to control adult noctuid pests with plant attractants—Introduction. Southwest. Entomol. 1998, 21, 1–7. [Google Scholar]
- Shorthouse, J.D. Ecoregions of Canada’s Prairie Grasslands. In Arthropods of Canadian Grasslands; Shorthouse, P.D., Floate, K.D., Eds.; Biological Survey of Canada: Ottowa, ON, Canada, 2010; pp. 53–81. [Google Scholar]
- Hardwick, D. Preparation of slide mounts of lepidopterous genitalia. Can. Entomol. 1950, 82, 231–235. [Google Scholar] [CrossRef]
- Lafontaine, J.D.; Poole, R.W. The Moths of America North of Mexico Including Greenland. Vol. Fascicle 25.1 Noctuoidea, Noctuidae (Part): Plusiinae. In The Moths of America North of Mexico; The Wedge Entomological Reseach Foundation: Washington, DC, USA, 1991; p. 182. [Google Scholar]
- Lafontaine, J.D. The Moths of America North of Mexico Including Greenland. Fascicle 27.2. Noctuoidea, Noctuidae. Vol. Fascicle 27.2. Noctuoidea, Noctuidae (Part), Noctuinae (Part—Euxoa). In The Moths of America North of Mexico; The Wedge Entomological Research Foundation: Washinton, DC, USA, 1987; p. 237. [Google Scholar]
- Lafontaine, J.D. The Moths of America North of Mexico Including Greenland: Fascicle 27.1 Noctuoidea, Noctuidae. Vol. Fascicle 27.1 Noctuoidea, Noctuidae (Part): Noctuinae (Part—Agrotini). In The Moths of America North of Mexico; The Wedge Entomological Reseach Foundation: Washington, DC, USA, 2004; p. 385. [Google Scholar]
- Lafontaine, J.D. The Moths of America North of Mexico Including Greenland. Vol. Fascicle 27.3 Noctuoidea, Noctuidae (Part): Noctuinae (Part—Noctuini). In The Moths of America North of Mexico; The Wedge Entomological Reseach Foundation: Washington, DC, USA, 1998; p. 348. [Google Scholar]
- Mikkola, K.; Lafontaine, J.D.; Gill, J. The Moths of North America Including Greenland. Vol. Fascicle 26.9 Noctuoidea: Noctuidae. Xyleninae, Apameini (Apamea Group). In The Moths of America North of Mexico; The Wedge Entomological Reseach Foundation: Washington, DC, USA, 2009; p. 191. [Google Scholar]
- Batallas, R.E.; Evenden, M.L. Influence of host plant species and fertilization regime on larval performance and feeding preference of the redbacked cutworm and the pale western cutworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 2020, 113, 731–741. [Google Scholar] [CrossRef]
- Crawley, M.J. The R Book, 2nd ed.; John Wiley & Sons: West Sussex, UK, 2012; p. 1051. [Google Scholar]
- Zuur, A.; Ieno, E.; Walker, N.; Saveliev, A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R. Spring Science: New York, NY, USA, 2009; p. 574. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models; R Foundation: Vienna, Austria, 2014; Available online: http://CRAN.R-project.org/package=nlme (accessed on 7 April 2018).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011; p. 449. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, L. Mating and oviposition of the pale western cutworm, Agrotis orthogonia Morrison (Lepidoptera: Noctuidae), in the laboratory. Can. Entomol. 1965, 97, 994–1000. [Google Scholar] [CrossRef]
- Schneider, J.C. Dispersal of a highly vagile insect in a heterogeneous environment. Ecology 1999, 80, 2740–2749. [Google Scholar] [CrossRef]
- Delisle, J.; McNeil, J.N. Calling behavior and pheromone titer of the true armyworm Pseudaletia unipuncta (Haw) (Lepidoptera: Noctuidae) under different temperature and photoperiodic conditions. J. Insect Physiol. 1987, 33, 315–324. [Google Scholar] [CrossRef]
- Fields, P.G.; McNeil, J.N. The overwintering potential of true armyworm, Pseudaletia unipuncta (Lepidoptera: Noctuidae), populations in Quebec. Can. Entomol. 1984, 116, 1647–1652. [Google Scholar] [CrossRef]
- Lemmen-Lechelt, J.K.; Wist, T.J.; Evenden, M.L. State-dependent plasticity in response to host-plant volatiles in a long-lived moth, Caloptilia fraxinella (Lepidoptera: Gracillariidae). J. Chem. Ecol. 2018, 44, 276–287. [Google Scholar] [CrossRef]
- Saveer, A.M.; Kromann, S.H.; Birgersson, G.; Bengtsson, M.; Lindblom, T.; Balkenius, A.; Hansson, B.S.; Witzgall, P.; Becher, P.G.; Ignell, R. Floral to green: Mating switches moth olfactory coding and preference. Proc. R. Soc. B 2012, 279, 2314–2322. [Google Scholar] [CrossRef]
- Reddy, G.; Guerrero, A. Behavioral responses of the diamondback moth, Plutella xylostella, to green leaf volatiles of Brassica oleracea subsp. Capitata. J. Agric. Food Chem. 2000, 48, 6025–6029. [Google Scholar] [CrossRef]
- Miluch, C.; Dosdall, L.; Evenden, M. Factors influencing male Plutella xylostella (Lepidoptera: Plutellidae) capture rates in sex pheromone-baited traps on canola in western canada. J. Econ. Entomol. 2014, 107, 2067–2076. [Google Scholar] [CrossRef]
- Evenden, M.; Whitehouse, C.; Onge, A.S.; Vanderark, L.; Lafontaine, J.-P.; Meers, S.; Cárcamo, H. Potential for semiochemical-based monitoring of the pea leaf weevil (Coleoptera: Curculionidae) on field pea (Fabaceae) in the Canadian Prairie Provinces. Can. Entomol. 2016, 148, 595–602. [Google Scholar] [CrossRef]
- Veromann, E.; Toome, M.; Kanaste, A.; Kaasik, R.; Copolovici, L.; Flink, J.; Kovacs, G.; Narits, L.; Luik, A.; Niinemets, U. Effects of nitrogen fertilization on insect pests, their parasitoids, plant diseases and volatile organic compounds in Brassica napus. Crop Protect. 2013, 43, 79–88. [Google Scholar] [CrossRef]
- Piesik, D.; Panka, D.; Delaney, K.J.; Skoczek, A.; Lamparski, R.; Weaver, D.K. Cereal crop volatile organic compound induction after mechanical injury, beetle herbivory (Oulema spp.), or fungal infection (Fusarium spp.). J. Plant Physiol. 2011, 168, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Piesik, D.; Lyszczarz, A.; Tabaka, P.; Lamparski, R.; Bocianowski, J.; Delaney, K.J. Volatile induction of three cereals: Influence of mechanical injury and insect herbivory on injured plants and neighbouring uninjured plants. Ann. Appl. Biol. 2010, 157, 425–434. [Google Scholar] [CrossRef]
- Meagher, R.L., Jr. Trapping noctuid moths with synthetic floral volatile lures. Entomol. Exp. Appl. 2002, 103, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Bestmann, H.J.; Vostrowsky, O.; Platz, H. Male sex pheromones of Noctuidae. Experientia 1977, 33, 874–875. [Google Scholar] [CrossRef]
- Fitzpatrick, S.M.; Mcneil, J.N.; Miller, D. Age-specific titer and antennal perception of acetic acid, a component of male Pseudaletia unipuncta (Haw.) hairpencil secretion. J. Chem. Ecol. 1989, 15, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J.; Smithhisler, C.S.; Reed, H.C.; McDonough, L.M. Trapping social wasps (Hymenoptera: Vespidae) with acetic acid and saturated short chain alcohols. J. Econ. Entomol. 2000, 93, 1613–1618. [Google Scholar] [CrossRef]
- Burguiere, L.; Marion-Poll, F.; Cork, A. Electrophysiological responses of female Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) to synthetic host odours. J. Insect Physiol. 2001, 47, 509–514. [Google Scholar] [CrossRef]
- Dötterl, S.; Jürgens, A.; Seifert, K.; Laube, T.; Weissbecker, B.; Schütz, S. Nursery pollination by a moth in Silene latifolia: The role of odours in eliciting antennal and behavioural responses. New Phytol. 2006, 169, 707–718. [Google Scholar] [CrossRef]
- Ômura, H.; Honda, K.; Hayashi, N. Chemical and chromatic bases for preferential visiting by the cabbage butterfly, Pieris rapae, to rape flowers. J. Chem. Ecol. 1999, 25, 1895–1906. [Google Scholar] [CrossRef]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects–finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L. Trends in floral scent chemistry in pollination syndromes: Floral scent composition in moth-pollinated taxa. Bot. J. Linn. Soc. 1993, 113, 263–284. [Google Scholar] [CrossRef]
- Anton, S.; Dufour, M.C.; Gadenne, C. Plasticity of olfactory-guided behaviour and its neurobiological basis: Lessons from moths and locusts. Entomol. Exp. Appl. 2007, 123, 1–11. [Google Scholar] [CrossRef]
- Rojas, J.C. Electrophysiological and behavioral responses of the cabbage moth to plant volatiles. J. Chem. Ecol. 1999, 25, 1867–1883. [Google Scholar] [CrossRef]
- Byers, J.A.; Anderbrant, O.; Lofqvist, J. Effective attraction radius—A method for comparing species attractants and determining densities of flying insects. J. Chem. Ecol. 1989, 15, 749–765. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Lure | Components | Ratio | Amount | Time Deployed |
|---|---|---|---|---|---|
| 2014 | Redbacked cutworm (RBC) | Z5-12Ac, Z7-12Ac, Z9-12Ac, Z5-10Ac | 200 2 1 1 | 1000 µg | 23 Jun–10 Oct |
| Bertha armyworm (BAW) | Z11-16Ac, Z9-14Ac | 95 5 | 500 µg | 10 Jun–02 Sep | |
| True armyworm (TAW) | Z11-16Ac | 1 | 1000 µg | 10 Jun–10 Oct | |
| Army cutworm (ACW) | Z5-14Ac, Z7-14Ac, Z9-14Ac | 100 1 10 | 100 µg | 02 Sep–10 Oct | |
| AAMB | Acetic acid, 3-methyl-1-butanol | 1 1 | 10 mL | 10 Jun–10 Oct | |
| Unbaited control | - | - | - | 10 Jun–10 Oct | |
| 2015 | RBC | Z5-12Ac, Z7-12Ac, Z9-12Ac, Z5-10Ac | 200 2 1 1 | 1000 µg | 22 Jun–15 Sept |
| BAW | Z11-16Ac, Z9-14Ac | 95 5 | 500 µg | 22 Jun–04 Aug | |
| TAW | Z11-16Ac | 1 | 1000 µg | 22 Jun–15 Sept | |
| Pale western cutworm (PWC) | Z7-12Ac, Z5-12Ac | 2 1 | 500 µg | 22 Jun–15 Sept | |
| AAMB | Acetic acid, 3-methyl-1-butanol | 1 1 | 10 mL | 22 Jun–15 Sept | |
| AAMB+MP | Acetic acid, 3-methyl-1-butanol, 2-methyl-1-propanol | 1 1 1 | 10 mL | 22 Jun–15 Sept | |
| AAMB+PAA | Acetic acid, 3-methyl-1-butanol, phenylacetaldehyde | 1 1 1 | 10 mL | 22 Jun–15 Sept | |
| AAMB+MP+PAA | Acetic acid, 3-methyl-1-butanol, 2-methyl-1-propanol, phenylacetaldehyde | 1 1 1 1 | 10 mL | 22 Jun–15 Sept | |
| Unbaited control | - | - | - | 22 Jun–15 Sept |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batallas, R.E.; Evenden, M.L. Fermented or Floral? Developing a Generalized Food Bait Lure to Monitor Cutworm and Armyworm Moths (Lepidoptera: Noctuidae) in Field Crops. Insects 2023, 14, 106. https://doi.org/10.3390/insects14020106
Batallas RE, Evenden ML. Fermented or Floral? Developing a Generalized Food Bait Lure to Monitor Cutworm and Armyworm Moths (Lepidoptera: Noctuidae) in Field Crops. Insects. 2023; 14(2):106. https://doi.org/10.3390/insects14020106
Chicago/Turabian StyleBatallas, Ronald E., and Maya L. Evenden. 2023. "Fermented or Floral? Developing a Generalized Food Bait Lure to Monitor Cutworm and Armyworm Moths (Lepidoptera: Noctuidae) in Field Crops" Insects 14, no. 2: 106. https://doi.org/10.3390/insects14020106
APA StyleBatallas, R. E., & Evenden, M. L. (2023). Fermented or Floral? Developing a Generalized Food Bait Lure to Monitor Cutworm and Armyworm Moths (Lepidoptera: Noctuidae) in Field Crops. Insects, 14(2), 106. https://doi.org/10.3390/insects14020106