
Cricket Meal (Gryllus bimaculatus) as a Protein Supplement on In Vitro Fermentation Characteristics and Methane Mitigation

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Chemical Composition of Experimental Feeds
3.2. Measurement of Parameters Based on In Vitro Gas Production Technique and Rumen Degradability of Nutrients
3.3. Volatile Fatty Acid and Methane Production
3.4. Ruminal pH, Ammonia–Nitrogen (NH3-N) Concentration and Protozoal Population
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.C.; Rosegrant, M.W.; Koo, J.; Robertson, R.; Sulser, T.; Zhu, T.; Ringler, C.; Msangi, S.; Palazzo, A.; Batka, M.; et al. Climate change: Impact on Agriculture and Costs of Adaptation. Int. Food Policy Res. Inst. 2009. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu, O.; Olaleru, I.F.; Oke, T.D.; Adepegba, V.A.; Usman, B. Performance of broiler chicken fed diets containing cassava peel and leaf meals as replacements for maize and soya bean meal. Int. J. Sci. Technol. 2015, 4, 169–173. [Google Scholar]
- Jayanegara, A.; Yantina, N.; Novandri, B.; Laconi, E.B. Evaluation of some insects as potential feed ingredients for ruminants: Chemical composition, in vitro rumen fermentation and methane emissions. J. Indones. Trop. Anim. Agric. 2017, 42, 247–254. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State of the art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Moyo, S.; Masika, P.J.; Muchenje, V. The potential of Imbrasia belina worm as a poultry and fish feed. A review. J. Anim. Feed Sci. 2019, 28, 209–219. [Google Scholar] [CrossRef]
- Marareni, M.; Mnisi, C.M. Growth performance, serum biochemistry and meat quality traits of Jumbo quails fed with mopane worm (Imbrasia belina) meal-containing diets. Vet. Anim. Sci. 2020, 10, 100141. [Google Scholar] [CrossRef]
- Sanchez-Muros, M.J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Wang, D.; Yao, Y.B.; Li, J.H.; Zhang, C.X. Nutriotional value of the field cricket (Gryllus testaceus Walker). Insect Sci. 2004, 11, 275–283. [Google Scholar] [CrossRef]
- Chung, Y.C.; Su, Y.P.; Chen, C.C.; Jia, G.; Wang, H.L.; Wu, J.C.G.; Lin, J.G. Relationship between antibacterial activity of chitosans and surface characteristics of cell wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar] [PubMed]
- Frye, F.L.; Calvert, C.C. Preliminary information on the nutritional content of mulberry silk moth (Bombyx mori) larvae. J. Zoo Wildl. Med. 1989, 20, 73–75. [Google Scholar]
- Kohler, R.; Kariuki, L.; Lambert, C.; Biesalski, H.K. Protein, amino acid and mineral composition of some edible insects from Thailand. J. Asia Pac. Entomol. 2019, 22, 372–378. [Google Scholar] [CrossRef]
- Jozefiak, D.; Jozefiak, A.; Kieronczyk, B.; Rawski, M.; Swiatkiewicz, S.; Dlugosz, J.; Engberg, R.M. Insects–a natural nutrient source for poultry–a review. Ann. Anim. Sci. 2016, 16, 297–313. [Google Scholar] [CrossRef] [Green Version]
- Piccolo, G.; Iaconisi, V.; Marono, S.; Gasco, L.; Loponte, R.; Nizza, S.; Parisi, G. Effect of Tenebrio molitor larvae meal on growth performance, in vivo nutrients digestibility, somatic and marketable indexes of gilthead sea bream (Sparus aurata). Anim. Feed Sci. Technol. 2017, 226, 12–20. [Google Scholar] [CrossRef]
- Secci, G.; Moniello, G.; Gasco, L.; Bovera, F.; Parisi, G. Barbary partridge meat quality as affected by Hermetia illucens and Tenebrio molitor larva meals in feeds. Food Res. Int. 2018, 112, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Cullere, M.; Tasoniero, G.; Giaccone, V.; Acuti, G.; Marangon, A.; Dalle Zotte, A. Black soldier fly as dietary protein source for broiler quails: Meat proximate composition, fatty acid and amino acid profile, oxidative status and sensory traits. Animal 2018, 12, 640–647. [Google Scholar] [CrossRef]
- Getachew, G.; Blümmel, M.; Makkar, H.; Becker, K. In vitro gas measuring techniques for assessment of nutritional quality of feeds: A review. Anim. Feed Sci. Technol. 1998, 72, 261–281. [Google Scholar] [CrossRef]
- Patra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Wang, D.; Zhai, S.W.; Zhang, C.X.; Bai, Y.Y.; An, S.H.; Xu, Y.N. Evaluation on nutritional value of field crickets as a poultry feedstuff. Asian-Australas. J. Anim. Sci. 2005, 18, 667–670. [Google Scholar] [CrossRef]
- Permatahati, D.; Mutia, R.; Astuti, D.A. Effect of Cricket Meal (Gryllus bimaculatus) on Production and Physical Quality of Japanese Quail Egg. Trop. Anim. Sci. J. 2019, 42, 53–58. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Wanapat, M.; Viennasay, B. Supplementation of banana flower powder pellet and plant oil sources on in vitro ruminal fermentation, digestibility, and methane production. Trop. Anim. Sci. J. 2016, 48, 1673–1678. [Google Scholar] [CrossRef] [PubMed]
- Orskov, E.R.; McDonal, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Mathew, S.; Sagathewan, S.; Thomas, J.; Mathen, G. An HPLC method for estimation of volatile fatty acids of ruminal fluid. Indian J. Anim. Sci. 1997, 67, 805–811. [Google Scholar]
- Galyean, M. Laboratory Procedure in Animal Nutrition Research; Department of Animal and Range Sciences, New Mexico State University: Las Cruces, NM, USA, 1989; Volume 188. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B. A Laboratory Manual for Animal Science; Cornell University Press: Ithaca, NY, USA, 1985. [Google Scholar]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Anim. Res. 2000, 49, 231–253. [Google Scholar] [CrossRef] [Green Version]
- SAS (Statistical Analysis System). User’s Guide: Statistic, 9, 3th ed.; SAS Inst. Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Crichton, N. Information point: Tukey multiple comparison test. J. Clin. Nurs. 1999, 8, 299–304. [Google Scholar]
- Wanapat, M.; Sundstøl, F.; Garmo, T.H. A comparison of alkali treatment methods to improve the nutritive value of straw. Anim. Feed Sci. Technol. 1985, 12, 295–309. [Google Scholar] [CrossRef]
- Chakravorty, J.; Ghosh, S.; Meyer-Rochow, V.B. Nutritional composition of Chondacris rosea and Brachytrupes orientalis: Two common insects used as food by tribes of Arunachal Pradesh, India. J. Asia-Pac. Entomol. 2014, 17, 48–56. [Google Scholar] [CrossRef]
- Hanboonsong, Y.; Jamjanya, T.; Durst, P.B. Six-Legged Livestock: Edible Insect Farming, Collecting and Marketing in Thailand; Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 2013. [Google Scholar]
- Shumo, M.; Osuga, I.M.; Khamis, F.M.; Tanga, C.M.; Fiaboe, K.K.M.; Subramanian, S.; Ekesi, S.; Van Huis, A.; Borgemeister, C. The nutritive value of black soldier fly larvae reared on common organic waste streams in Kenya. Sci. Rep. 2019, 9, 10110. [Google Scholar] [CrossRef] [PubMed]
- Veldkamp, T.; Dong, L.; Paul, A.; Govers, C. Bioactive properties of insect products for monogastric animals—A review. J. Insects Food Feed. 2021, 1–14. [Google Scholar] [CrossRef]
- Ahmed, E.; Fukuma, N.; Hanada, M.; Nishida, T. Insects as Novel Ruminant Feed and a Potential Mitigation Strategy for Methane Emissions. Animals 2021, 11, 2648. [Google Scholar] [CrossRef] [PubMed]
- Bellezza-Oddon, S.; Biasato, I.; Imarisio, A.; Pipan, M.; Dekleva, D.; Capucchio, M.T.; Meneguz, M.; Stefania, B.; Barbero, R.; Gariglio, M.; et al. Black Soldier Fly and Yellow Mealworm live larvae for broiler chickens: Effects on bird performance and health status. J. Anim. Physiol. Anim. Nutr. 2021, 105, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Getachew, G.; DePeters, E.J.; Robinson, P.H.; Taylor, S.J. In vitro rumen fermentation and gas production: Influence of yellow grease, tallow, corn oil and their potassium soaps. Anim. Feed Sci. Technol. 2001, 93, 1–15. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; McGinn, S.M.; Petit, H. Methane abatement strategies for cattle: Lipid supplementation of diets. Can. J. Anim. Sci. 2007, 87, 431–440. [Google Scholar] [CrossRef]
- Jayanegara, A.; Goel, G.; Makkar, H.P.S.; Becker, K. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol. 2015, 209, 60–68. [Google Scholar] [CrossRef]
- Wu, D.; Xu, L.; Tang, S.; Guan, L.; He, Z.; Guan, Y.; Tan, Z.; Han, X.; Zhou, C.; Kang, J.; et al. Influence of oleic acid on rumen fermentation and fatty acid formation in vitro. PLoS ONE 2016, 11, e0156835. [Google Scholar] [CrossRef]
- Machmuller, A. Medium-chain fatty acids and their potential to reduce methanogenesis in domestic ruminants. Agric. Ecosyst. Environ. 2006, 112, 107–114. [Google Scholar] [CrossRef]
- Calabrò, S.; Cutrignelli, M.I.; Piccolo, G.; Bovera, F.; Zicarelli, F.; Gazaneo, M.P.; Infascelli, F. In vitro fermentation kinetics of fresh and dried silage. Anim. Feed Sci. Technol. 2005, 123, 129–137. [Google Scholar] [CrossRef]
- Russell, J.B.; Rychlik, J.L. Factors that alter rumen microbial ecology. Science 2001, 292, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Kazemi-Bonchenari, M.; Rezayazdi, K.; Nikkhah, A.; Kohram, H.; Dehghan-Banadaky, M. The effects of different levels of sodium caseinate on rumen fermentation pattern, digestibility and microbial protein synthesis of Holstein dairy cows. Afr. J. Biotechnol. 2010, 9, 1990–1998. [Google Scholar]
- Owens, F.N.; Qi, S.; Sapienza, D.A. Applied protein nutrition of ruminants–current status and future directions. Prof. Anim. Sci. 2014, 30, 150–179. [Google Scholar] [CrossRef]
- Jayanegara, A.; Dewi, S.P.; Ridla, M. Nutrient content, protein fractionation, and utilization of some beans as potential alternatives to soybean for ruminant feeding. Med. Pet. 2016, 39, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Phesatcha, K.; Phesatcha, B.; Wanapat, M.; Cherdthong, A. Roughage to concentrate ratio and saccharomyces cerevisiae inclusion could modulate feed digestion and in vitro ruminal fermentation. Vet. Sci. 2020, 7, 151. [Google Scholar] [CrossRef]
- Viennasay, B.; Wanpat, M.; Totkul, P.; Phesatcha, B.; Ampapon, T.; Cherdthong, A. Effect of Flemingia macrophylla silage on in vitro fermentation characteristics and reduced methane production. Anim. Prod. Sci. 2020, 60, 1918–1924. [Google Scholar] [CrossRef]
- Brown, M.S.; Ponce, C.H.; Pulikanti, R. Adaptation of beef cattle to high-concentrate diets: Performance and ruminal metabolism. J. Anim. Sci. 2006, 84, 25–33. [Google Scholar] [CrossRef]
- Boussaada, A.; Arhab, R.; Calabrò, S.; Grazioli, R.; Ferrara, M.; Musco, N.; Thlidjane, M.; Cutrignelli, M.I. Effect of Eucalyptus globulus leaves extracts on in vitro rumen fermentation, methanogenesis, degradability and protozoa population. Ann. Anim. Sci. 2018, 18, 753–767. [Google Scholar] [CrossRef] [Green Version]
- Benchaar, C.; McAllister, T.A.; Chouinard, P.Y. Digestion, ruminal fermentation, ciliate protozoal populations, and milk production from dairy cows fed cinnamaldehyde, quebracho condensed tannin, or Yucca schidigera saponin extracts. J. Dairy Sci. 2008, 91, 4765–4777. [Google Scholar] [CrossRef]
- Lee, M.R.F.; Harris, L.J.; Dewhurst, R.J.; Merry, R.J.; Scollan, N.D. The effect of clover silages on long chain fatty acid rumen transformations and digestion in beef steers. J. Anim. Sci. 2003, 76, 491–501. [Google Scholar] [CrossRef]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.J. Effects and mode of action of chitosan and ivy fruit saponins on the microbiome, fermentation and methanogenesis in the rumen simulation technique. FEMS Microbiol. Ecol. 2016, 92, 1. [Google Scholar]


{kind=link}
| Items | Replacement Levels of CM for SBM (% Fresh Basis) | CM | RS | SBM | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 25 | 50 | 75 | 100 | ||||
| Ingredients (% as fed) | ||||||||
| Cassava chip | 60.0 | 60.0 | 58.0 | 55.0 | 55.0 | |||
| Soybean meal | 16.0 | 12.0 | 8.0 | 4.0 | 0.0 | |||
| Cricket meal | 0.0 | 4.0 | 8.0 | 12.0 | 16.0 | |||
| Coconut meal | 7.0 | 7.0 | 7.0 | 8.0 | 8.0 | |||
| Rice bran | 7.0 | 7.0 | 8.0 | 7.8 | 7.6 | |||
| Palm meal | 5.0 | 5.0 | 6.0 | 7.0 | 7.0 | |||
| Molasses | 1.0 | 1.0 | 1.0 | 2.0 | 2.0 | |||
| Urea | 1.0 | 1.0 | 1.0 | 1.2 | 1.4 | |||
| Sulfur | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |||
| Mineral mixed | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |||
| Salt | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | |||
| Chemical composition | ||||||||
| Dry matter (%) | 90.3 | 90.8 | 92.3 | 92.7 | 93.1 | 94.2 | 91.6 | 90.5 |
| ------------------------------------% dry matter ----------------------------------- | ||||||||
| Organic matter | 93.6 | 94.5 | 94.1 | 93.6 | 94.4 | 96.6 | 90.7 | 94.3 |
| Ash | 6.4 | 5.5 | 5.9 | 6.4 | 5.6 | 3.4 | 9.3 | 5.7 |
| Crude protein | 14.0 | 14.1 | 14.0 | 14.3 | 14.4 | 68.5 | 2.2 | 40.1 |
| Neutral detergent fiber | 20.1 | 20.8 | 24.7 | 24.5 | 24.3 | 33.1 | 75.5 | 12.8 |
| Acid detergent fiber | 10.7 | 10.5 | 12.2 | 13.5 | 13.1 | 12.7 | 47.4 | 9.7 |
| Ether extract | 2.2 | 7.2 | 8.0 | 10.6 | 11.3 | 12.5 | - | 13.2 |
| Treatments | R:C Ratio | SBM:CM | Gas Production Kinetics | Cumulative Gas Production at 96 h (mL/0.5 g DM Substrate) | In Vitro DM Degradability (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| a | b | c | a + b | 12 h | 24 h | ||||
| T1 | 60:40 | 100:0 | −0.4 | 70.7 | 0.04 | 70.3 | 65.7 | 52.7 | 59.3 |
| T2 | 75:25 | −0.8 | 71.5 | 0.03 | 70.7 | 68.8 | 55.9 | 59.7 | |
| T3 | 50:50 | 1.3 | 69.2 | 0.04 | 71.5 | 69.7 | 57.0 | 60.2 | |
| T4 | 25:75 | 1.1 | 69.8 | 0.04 | 70.9 | 69.1 | 57.5 | 60.5 | |
| T5 | 0:100 | 1.6 | 69.1 | 0.04 | 70.7 | 70.2 | 56.5 | 61.6 | |
| T6 | 40:60 | 100:0 | −3.1 | 76.4 | 0.03 | 73.3 | 65.3 | 62.8 | 62.8 |
| T7 | 75:25 | −2.7 | 74.3 | 0.03 | 66.1 | 66.7 | 66.5 | 62.4 | |
| T8 | 50:50 | −0.8 | 71.5 | 0.03 | 70.7 | 68.7 | 64.6 | 64.0 | |
| T9 | 25:75 | −2.2 | 73.4 | 0.03 | 71.7 | 70.1 | 65.1 | 64.1 | |
| T10 | 0:100 | −1.1 | 70.8 | 0.05 | 69.7 | 72.1 | 69.6 | 65.6 | |
| SEM | 0.38 | 0.58 | 0.008 | 1.42 | 1.18 | 0.45 | 0.58 | ||
| Comparison R:C ratio SBM:CM R:C ratio × SBM:CM | |||||||||
| 0.06 | 0.03 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | |||
| 0.92 | 0.95 | 0.006 | 0.04 | 0.03 | 0.04 | 0.03 | |||
| 0.84 | 0.85 | 0.12 | 0.14 | 0.78 | 0.36 | 0.28 | |||
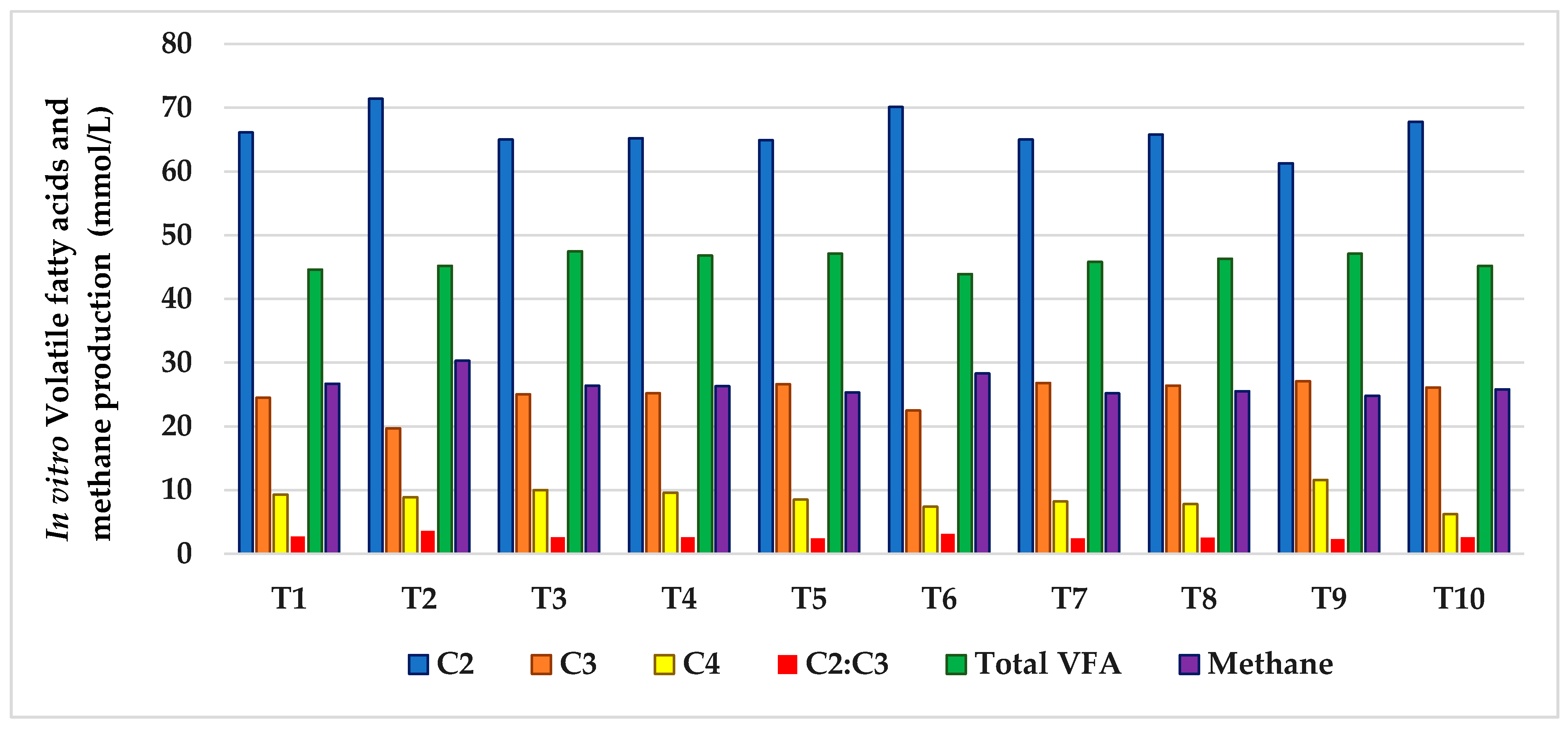
| Treatments | R:C Ratio | SBM:CM | Molar Proportions of VFA (mmol/L) | Total VFA (mmol/L) | CH4 (mmol/L) | |||
|---|---|---|---|---|---|---|---|---|
| C2 | C3 | C4 | C2:C3 | |||||
| T1 | 60:40 | 100:0 | 72.0 | 19.0 | 8.9 | 3.8 | 59.8 | 30.7 |
| T2 | 75:25 | 71.9 | 19.2 | 8.9 | 3.7 | 52.5 | 30.6 | |
| T3 | 50:50 | 71.5 | 20.5 | 7.9 | 3.5 | 56.3 | 29.7 | |
| T4 | 25:75 | 69.1 | 22.1 | 8.8 | 3.1 | 68.9 | 28.5 | |
| T5 | 0:100 | 65.2 | 24.6 | 10.2 | 2.6 | 63.6 | 26.6 | |
| T6 | 40:60 | 100:0 | 66.7 | 22.9 | 10.3 | 2.9 | 70.9 | 27.8 |
| T7 | 75:25 | 61.9 | 27.2 | 11.9 | 2.2 | 78.5 | 25.7 | |
| T8 | 50:50 | 56.9 | 31.4 | 11.7 | 1.8 | 73.9 | 21.6 | |
| T9 | 25:75 | 56.2 | 30.9 | 12.8 | 1.8 | 73.1 | 21.9 | |
| T10 | 0: 100 | 55.1 | 31.5 | 13.4 | 1.7 | 71.2 | 21.5 | |
| SEM | 0.04 | 0.06 | 0.26 | 0.05 | 1.05 | 0.14 | ||
| Comparison | ||||||||
| R:C ratio | 0.001 | 0.001 | 0.08 | 0.001 | 0.004 | 0.001 | ||
| SBM:CM | 0.001 | 0.009 | 0.81 | 0.004 | 0.54 | 0.004 | ||
| R:C ratio × SBM:CM | 0.26 | 0.65 | 0.14 | 0.81 | 0.66 | 0.07 | ||
| Treatments | R:C Ratio | SBM:CM | pH | NH3-N (mg/dL) | Protozoa (×105 Cells/mL) |
|---|---|---|---|---|---|
| T1 | 60:40 | 100:0 | 6.5 | 18.7 | 9.1 |
| T2 | 75:25 | 6.5 | 22.4 | 9.1 | |
| T3 | 50:50 | 6.5 | 26.3 | 8.7 | |
| T4 | 25:75 | 6.6 | 25.0 | 7.4 | |
| T5 | 0:100 | 6.7 | 26.0 | 8.1 | |
| T6 | 40:60 | 100:0 | 6.5 | 23.5 | 6.5 |
| T7 | 75:25 | 6.4 | 26.6 | 6.1 | |
| T8 | 50:50 | 6.4 | 27.8 | 6.4 | |
| T9 | 25:75 | 6.4 | 28.1 | 7.2 | |
| T10 | 0:100 | 6.4 | 28.4 | 6.7 | |
| SEM | 0.14 | 1.28 | 1.42 | ||
| Comparison | |||||
| R:C ratio | 0.001 | 0.04 | 0.001 | ||
| SBM:CM | 0.72 | 0.03 | 0.02 | ||
| R:C ratio × SBM:CM | 0.36 | 0.86 | 0.65 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phesatcha, B.; Phesatcha, K.; Viennaxay, B.; Matra, M.; Totakul, P.; Wanapat, M. Cricket Meal (Gryllus bimaculatus) as a Protein Supplement on In Vitro Fermentation Characteristics and Methane Mitigation. Insects 2022, 13, 129. https://doi.org/10.3390/insects13020129
Phesatcha B, Phesatcha K, Viennaxay B, Matra M, Totakul P, Wanapat M. Cricket Meal (Gryllus bimaculatus) as a Protein Supplement on In Vitro Fermentation Characteristics and Methane Mitigation. Insects. 2022; 13(2):129. https://doi.org/10.3390/insects13020129
Chicago/Turabian StylePhesatcha, Burarat, Kampanat Phesatcha, Bounnaxay Viennaxay, Maharach Matra, Pajaree Totakul, and Metha Wanapat. 2022. "Cricket Meal (Gryllus bimaculatus) as a Protein Supplement on In Vitro Fermentation Characteristics and Methane Mitigation" Insects 13, no. 2: 129. https://doi.org/10.3390/insects13020129
APA StylePhesatcha, B., Phesatcha, K., Viennaxay, B., Matra, M., Totakul, P., & Wanapat, M. (2022). Cricket Meal (Gryllus bimaculatus) as a Protein Supplement on In Vitro Fermentation Characteristics and Methane Mitigation. Insects, 13(2), 129. https://doi.org/10.3390/insects13020129