
Behavior of Nymphs and Adults of the Black-Legged Tick Ixodes scapularis and the Lone Star Tick Ambylomma americanum in Response to Thermal Stimuli



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
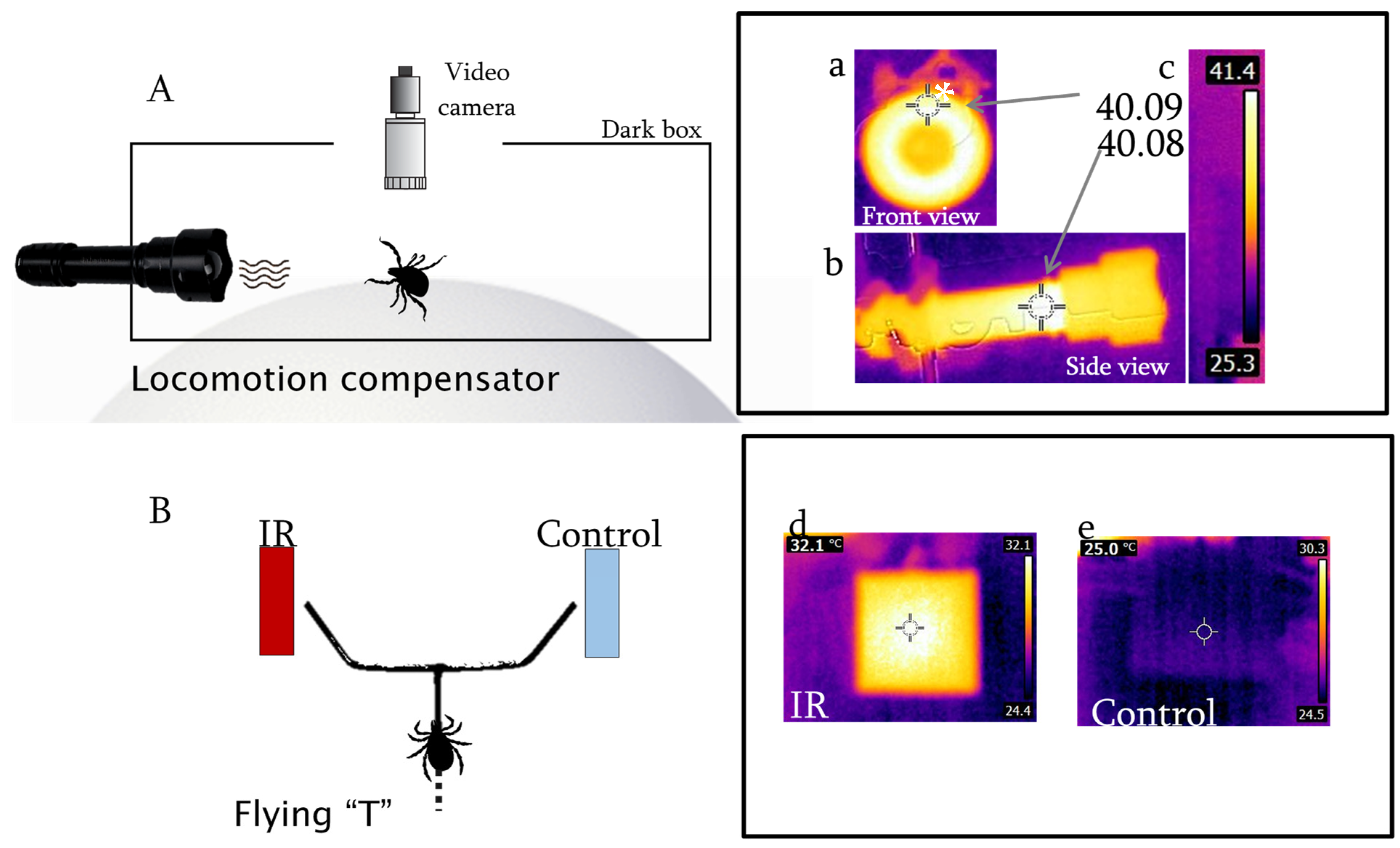
2.2. Behavioral Arenas
2.3. Visual Recording
2.4. Statistics
3. Results
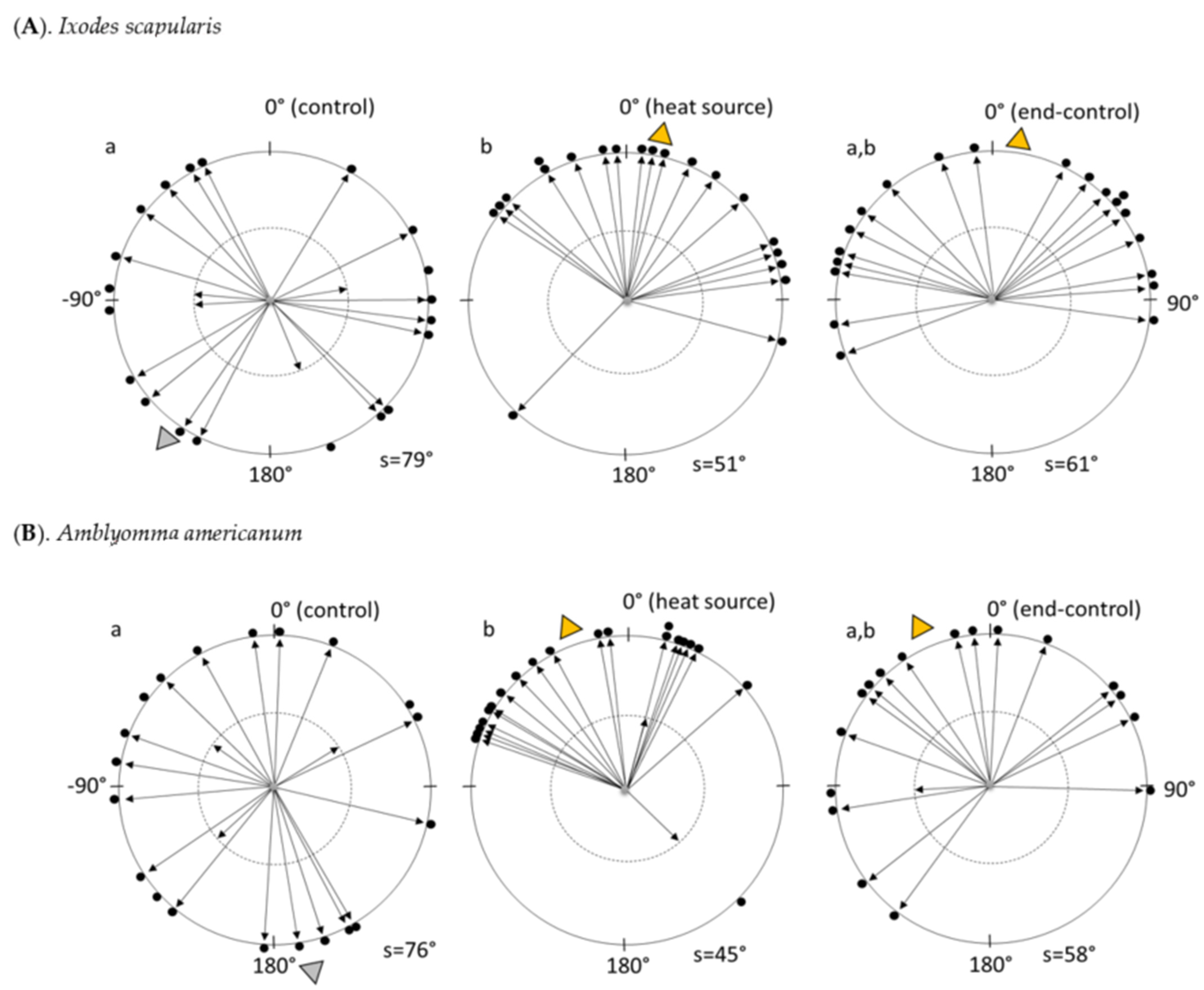
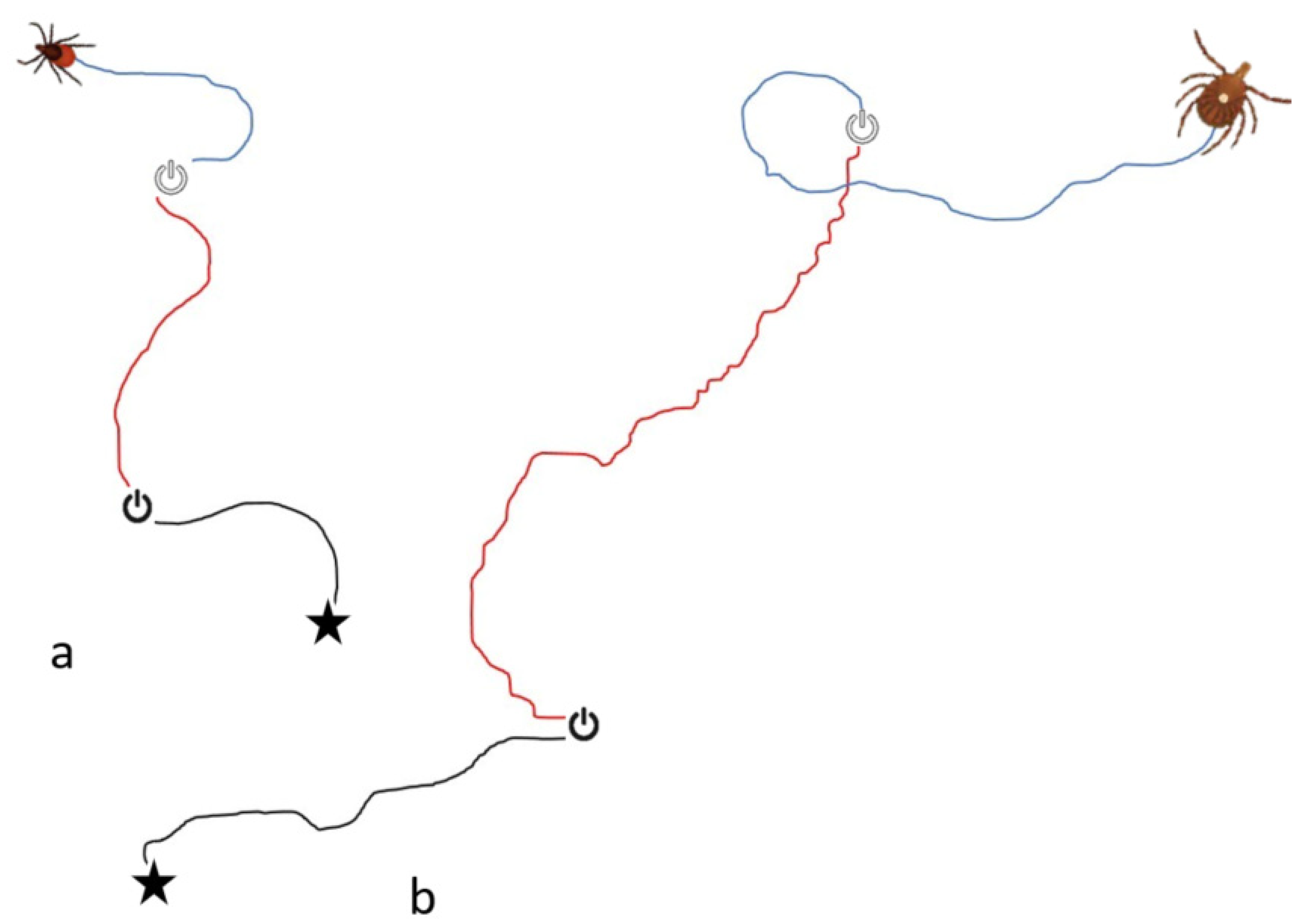
3.1. On the Servosphere
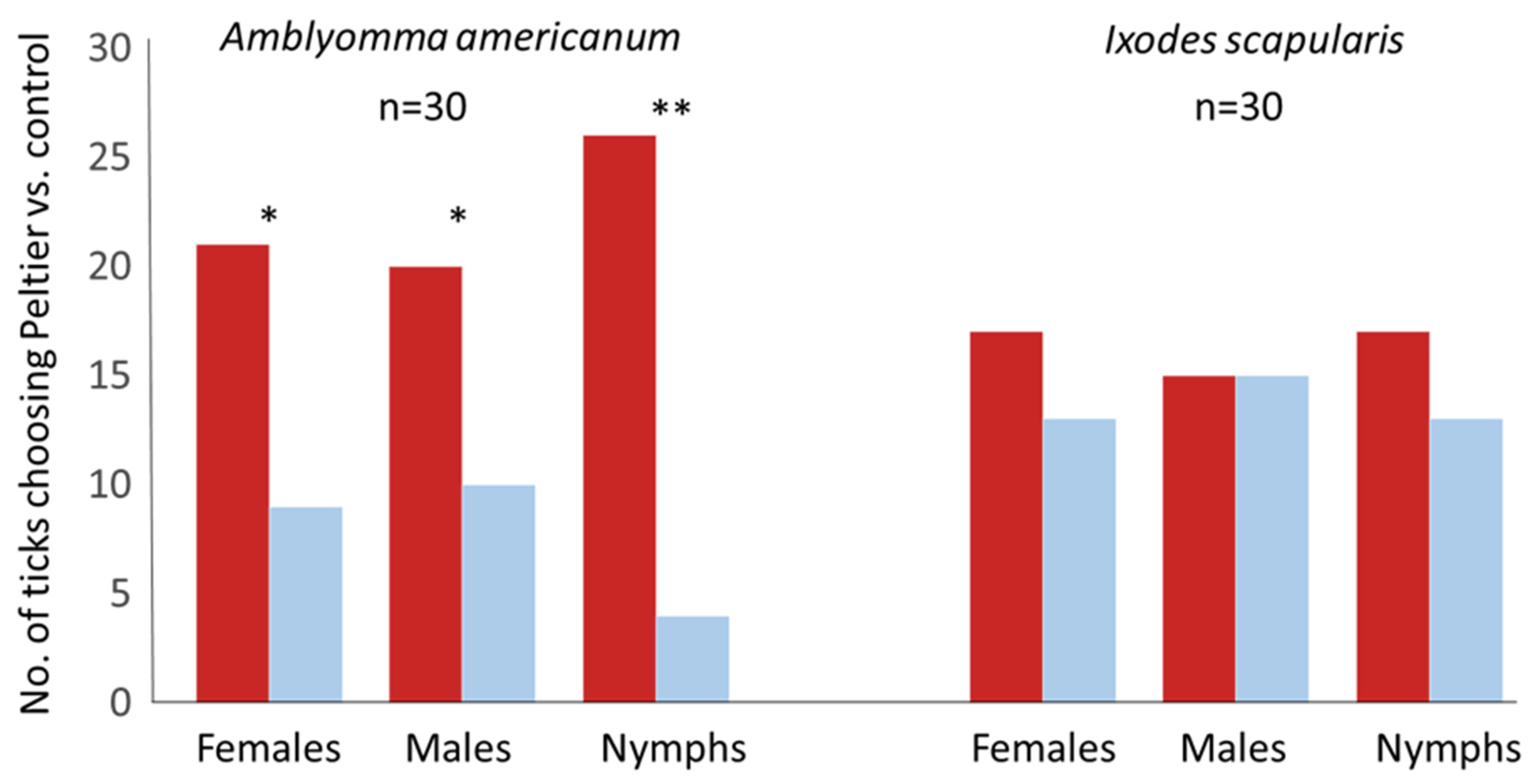
3.2. On the “Flying “T”
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lantos, P.M.; Nigrovic, L.E.; Auwaerter, P.G.; Fowler, V.G., Jr.; Ruffin, F.; Brinkerhoff, R.J.; Reber, J.; Williams, C.; Broyhill, J.; Pan, W.K.; et al. Geographic Expansion of Lyme Disease in the Southeastern United States, 2000–2014. Open Forum Infect. Dis. 2015, 2, ofv143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlgren, F.S.; Paddock, C.D.; Springer, Y.P.; Eisen, R.J.; Behravesh, C.B. Expanding Range of Amblyomma americanum and Simultaneous Changes in the Epidemiology of Spotted Fever Group Rickettsiosis in the United States. Am. J. Trop. Med. Hyg. 2016, 94, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes scapularis: An Increasing Public Health Concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.R.; Gaines, D.; Gilliam, W.F.; Brinkerhoff, R.J. Population Genetic Structure of the Lyme Disease Vector Ixodes scapularis at an Apparent Spatial Expansion Front. Infect. Genet. Evol. 2014, 27, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Brinkerhoff, R.J.; Gilliam, W.F.; Gaines, D. Lyme Disease, Virginia, USA, 2000–2011. Emerg. Infect. Dis 2014, 20, 1661–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diuk-Wasser, M.A.; vanAcker, M.C.; Fernandez, M.P. Impact of Land Use Changes and Habitat Fragmentation on the Eco-Epidemiology of Tick-Borne Diseases. J. Med. Entomol. 2021, 58, 1546–1564. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Radojevic, M.; Wu, X.; Duvvuri, V.R.; Leighton, P.A.; Wu, J. Estimated Effects of Projected Climate Change on the Basic Reproductive Number of the Lyme Disease Vector Ixodes scapularis. Environ. Health. Perspect. 2014, 122, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, R.K.; Peterson, A.T.; Cobos, M.E.; Ganta, R.; Foley, D. Current and Future Distribution of the Lone Star Tick, Amblyomma americanum (L.) (Acari: Ixodidae) in North America. PLoS ONE 2019, 14, e0209082. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Roe, R.M. Biology of Ticks; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Brinkerhoff, R.J.; Dang, L.; Streby, H.M.; Gimpel, M. Life History Characteristics of Birds Influence Patterns of Tick Parasitism. Infect. Ecol. Epidemiol. 2019, 9, 1547096. [Google Scholar] [CrossRef] [Green Version]
- Apperson, C.S.; Levine, J.F.; Evans, T.L.; Braswell, A.; Heller, J. Relative Utilization of Reptiles and Rodents as Hosts by Immature Ixodes scapularis (Acari: Ixodidae) in the Coastal Plain of North Carolina, USA. Exp. Appl. Acarol. 1993, 17, 719–731. [Google Scholar]
- Ogden, N.H.; Lindsay, L.R.; Hanincová, K.; Barker, I.K.; Bigras-Poulin, M.; Charron, D.F.; Heagy, A.; Francis, C.M.; O’Callaghan, C.J.; Schwartz, I.; et al. Role of Migratory Birds in Introduction and Range Expansion of Ixodes scapularis ticks of Borrelia burgdoferi and Anaplasma phagocytophilum in Canada. Appl. Exp. Microbiol. 2008, 74, 1780–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampo, M.; Bayliss, P. The Impact of Ticks on Bufo marinus from Native Habitats. Parasitology 1996, 113, 199–206. [Google Scholar] [CrossRef]
- Crispell, G.; Commins, S.P.; Archer-Hartman, S.A.; Choudhary, S.; Dharmarajan, G.; Azadi, P.; Karim, S. Discovery of Alpha-gal-containing Antigens in North American Tick Species Believed to Induce Red Meat Allergy. Front. Immunol. 2019, 10, 1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falco, R.C.; Fish, D. Horizontal Movement of Adult Ixodes dammini (Acari: Ixodidae) Attracted to CO2-bainted Traps. J. Med. Entomol. 1991, 28, 726–729. [Google Scholar] [CrossRef]
- McMahon, C.; Guerin, P.M.; Syed, Z. 1-Octen-3-ol Isolated from Bont Ticks attracts Amblyomma variegatum. J. Chem. Ecol. 2001, 27, 471–486. [Google Scholar] [CrossRef]
- McMahon, C.; Guerin, P.M. Attraction of the Tropical Bont Tick, Amblyomma variegatum, to Human Breath and to the Breath Components Acetone, NO and CO2. Naturwiss 2002, 89, 311–315. [Google Scholar]
- Donzé, G.; McMahon, C.; Guerin, P.M. Rumen Metabolites Serve Ticks to Exploit Large Mammals. J. Exp. Biol. 2004, 207, 4283–4289. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.F.; Pickens, L.G. Spectral Sensitivity to Light of Two Species of Ticks (Acarina: Ixodidae). Ann. Entomol. Soc. Am. 1987, 80, 256–262. [Google Scholar] [CrossRef]
- Phillis, W.A.; Cromroy, H.L. The Microanatomy of the Eye of Amblyoma americanum (Acari: Ixodidae) and Resultant Implications of its Structure. J. Med. Ent. 1977, 13, 685–698. [Google Scholar] [CrossRef]
- Szlendak, E.; Oliver, J.H., Jr. Anatomy of Synganglia, Including their Neurosecretory Regions, in Unfed, Virgin Female Ixodes scapularis Say (Acari: Ixodidae). J. Morphol. 1992, 213, 349–364. [Google Scholar] [CrossRef]
- Perret, J.L.; Guerin, P.M.; Diehl, P.A.; Vlimant, M.; Gern, L. Darkness Induce Mobility, and Saturation Deficit Limits Questing Duration, in the Tick Ixodes ricinus. J. Exp. Biol. 2003, 206, 1809–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klompen, J.S.H.; Oliver, J.H. Haller’s Organ in the Tick Family Argasidae (Acari: Parasitiformes: Ixodida). J. Parasitol. 1993, 79, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Steullet, P.; Guerin, P.M. Perception of Breath Components by the Tropical Bont Tick, Amblyomma variegatum Fabricius (Ixodidae). J. Comp. Physiol. A 1992, 170, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Guerin, P.M.; Kröber, T.; McMahon, C.; Guerenstein, P.; Grenacher, S.; Vlimant, M.; Diehl, P.A.; Steullet, P.; Syed, Z. Chemosensory and Behavioral Adaptations of Ectoparasitic Arthropods. Nova Acta Leopold. 2000, 83, 213–229. [Google Scholar]
- Soares, S.F.; Borges, L.M.F. Electrophysiological Responses of the Olfactory Receptors of the Tick Amblyomma cajennense (Acari: Ixodidae) to Host-related and Tick Pheromone-related Synthetic Compounds. Acta Tropica 2012, 124, 192–198. [Google Scholar] [CrossRef]
- Foelix, R.F.; Axtell, R.C. Ultrastructure of Haller’s Organ in the Tick Amblyomma americnaum (L.). Z. Zellforsch. Mikrosk. Anat. 1972, 124, 275–292. [Google Scholar] [CrossRef]
- Mitchell, R.D.; Zhu, J.; Carr, A.L.; Dhammi, A.; Cave, G.; Sonenshine, D.E.; Roe, R.M. Infrared Light Detection by the Haller’s Organ of Adult American Dog Ticks, Dermacentor variabilis (Ixodida: Ixodidae). Ticks Tick-Borne Dis. 2017, 8, 764–771. [Google Scholar] [CrossRef]
- Carr, A.L.; Salgado, V.L. Ticks Home in on Body Heat: A new Understanding of Haller’s Organ and Repellent Action. PLoS ONE 2019, 14, e0221659. [Google Scholar] [CrossRef] [Green Version]
- Lazzari, C.R.; Núñez, J.A. The Response to Radiant Heat and the Estimation of the Temperature of Distant Sources in Triatoma infestans. J. Insect Physiol. 1989, 35, 525–529. [Google Scholar] [CrossRef]
- Schmitz, H.; Trenner, S.; Hofmann, M.H.; Bleckmann, H. The Ability of Rhodnius prolixus (Hemiptera; Reduviidae) to Approach a Thermal Source Solely by its Infrared Radiation. J. Insect Physiol. 2000, 46, 745–751. [Google Scholar] [CrossRef]
- Schmitz, H.; Schmitz, A.; Bleckmann, H. A New Type of Infrared Organ in the Australian “Fire-Beetle” Merimna atrata (Coleoptera: Buprestidae). Naturwissenschaften 2000, 87, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.A.; Hartline, P.H. The Infrared Vision of Snakes. Sci. Am. 1982, 246, 116–127. [Google Scholar] [CrossRef]
- Kürten, L.; Schmidt, U. Thermoreception in the Common Vampire Bat (Desmodus rotundus). J. Comp. Physiol. 1981, 146, 223–228. [Google Scholar] [CrossRef]
- Gracheva, E.; Cordero-Morales, J.F.; González-Carcacía, J.A.; Ingolia, N.T.; Manno, C.; Aranguren, C.I.; Weissman, J.S.; Julius, D. Ganglion-specific Splicing of TRPV1 Underlies Infrared Sensation in Vampire Bats. Nature 2011, 476, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katić, K.; Li, R.; Kingma, B.; Zeiler, W. Modelling Hand Skin Temperature in Relation to Body Composition. J. Therm. Biol. 1917, 69, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kramer, E. The Orientation of Walking Honeybees in Odour Fields with Small Concentration Gradients. Physiol. Entomol. 1976, 1, 27–37. [Google Scholar] [CrossRef]
- Otálora-Luna, F.; Lapointe, S.L.; Dickens, J.C. Olfactory Cues are Subordinate to Visual Stimuli in a Neotropical Generalist Weevil. PLoS ONE 2013, 8, e53120. [Google Scholar] [CrossRef]
- Otálora-Luna, F.; Guerin, P.M. Amines from Vertebrates Guide Triatomine Bugs to Resources. J. Insect Physiol. 2014, 71, 52–60. [Google Scholar] [CrossRef]
- Bergman, T.L.; Lavine, A.S.; Incropera, F.P.; Dewitt, D.P. Introduction to Heat Transfer, 6th ed.; John Wiley & Sons, Inc.: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Bell, W.J. Searching Behaviour: The Behavioural Ecology of Finding Resources; Chapman and Hall: London, UK, 1991. [Google Scholar]
- Kitching, R.L.; Zalucki, M.P. Component Analysis and Modeling of the Movement Process: Analysis of Simple Tracks. Res. Popul. Ecol. 1982, 24, 224–238. [Google Scholar] [CrossRef]
- Batschelet, E. Circular Statistics in Biology; Academic Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Visser, J.H.; Piron, P.G.M. An Open Y-track Olfactometer for Recording of Aphid Behavioral Responses to Plant Odours. Exp. Appl. Entomol. 1998, 9, 41–46. [Google Scholar]
- Dickens, J.C. Predator-prey Interactions: Olfactory Adaptations of Generalist and Specialist Predators. Agric. For. Entomol. 1999, 1, 47–54. [Google Scholar] [CrossRef]
- Otálora-Luna, F.; Hammock, J.A.; Alessandro, R.T.; Lapointe, S.L.; Dickens, J.C. Discovery and Characterization of Chemical Signals for Citrus Root Weevil, Diaprepes abbreviatus. Arthropod.-Plant Interact. 2009, 3, 63–73. [Google Scholar] [CrossRef]
- Kastberger, G.; Stachl, R. Infrared Imaging Technology and Biological Implications. Behav. Res. Methods Instrum. Comput. 2003, 35, 429–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, N.P.; Niu, J.L. CFD Study of the Thermal Environment Around Human Body: A Review. Indoor Built. Environ. 2004, 14, 15–16. [Google Scholar] [CrossRef]
- Barnes, R.B. Thermography of the Human Body: Infrared-radiant Energy Provides New Concepts and Instrumentation for Medical Diagnosis. Science 1963, 140, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Ihaka, R.; Gentleman, R. R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 2 February 2021).
- Ruxton, G.D.; Beauchamp, G. Time for Some A Priori Thinking About Post Hoc Testing. Behav. Ecol. 2008, 19, 690–693. [Google Scholar] [CrossRef] [Green Version]
- Jammalamadaka, S.R.; Sengupta, A. Topics in Circular Statistics; World Scientific: Singapore, 2001; Volume 5. [Google Scholar]
- Brase, C.H.; Brase, C.P. Understanding Statistics; D.C. Health and Co.: Lexington, MA, USA, 1983. [Google Scholar]
- Liu, Y.; Liu, Z.; Luo, J. Numerical Investigation of the Unsteady Thermal Plume Around Human Body in Closed Space. Procedia Eng. 2015, 121, 1919–1926. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, A.N. The Concept of Nature; Project Gutenberg; Cosimo, Inc.: New York, NY, USA, 2006. [Google Scholar]
- García Bacca, J.D. Sobre Realismo; Anthropos Editorial: Barcelona, Spain, 2001. [Google Scholar]
- Otálora-Luna, F.; Fulmore, T.; Páez-Rondón, O.; Aldana, E.; Brinkerhoff, J. Good (and Bad) Words for the Ontological (and Anthropomorphic) Description of Behavior. Biosemiotics 2021, 1–22. [Google Scholar] [CrossRef]
- Dusenbery, D.B. Sensory Ecology; W.H. Freeman and Company: New York, NY, USA, 1992. [Google Scholar]
- Webb, J.P. Host-locating Behavior of Nymphal Ornithodoros concanensis (Acarina: Argasidae). J. Med. Entomol. 1979, 16, 437–447. [Google Scholar] [CrossRef]
- Oorebeek, M.; Sharrad, R.; Kleindorfer, S. What Attracts Ixodes hirsti (Acari: Ixodidae) to their Host? Parasitol. Res. 2009, 104, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, H.; Schmitz, A.; Schneider, E.S. Matched filter properties of infrared receptors used for fire and heat detection in insects. In The Ecology of Animal Senses: Matched Filters for Economical Sensing; von der Emde, G., Warrant, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 207–234. [Google Scholar]
- Benellli, G.; Romano, D.; Rocchigiani, G.; Caselli, A.; Mancianti, F.; Canale, A.; Stefanini, C. Behavioral Asymmetries in Ticks—Lateralized Questing of Ixodes ricinus to a Mechatronic Apparatus Delivering Host-borne Cues. Acta Trop. 2018, 178, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.D. The Radiation of Heat from the Human Body: III. The Human Skin as a Black-Body Radiator. J. Clin. Investig. 1934, 13, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Voelker, C.; Maempel, S.; Kornadt, O. Measuring the Human Body’s Microclimate Using a Thermal Manikin. Indoor Air 2014, 24, 567–579. [Google Scholar] [CrossRef]
- Otálora-Luna, F.; Dickens, J.; Brinkerhoff, J.; Li, A.Y. Geotropic, Hydrokinetic and Random Walking Differ Between Sympatric Tick Species: The Deer Tick Ixodes scapularis and the Lone Star tick Ambylomma americanum. J. Ethol. 2022. [Google Scholar] [CrossRef]
- Otálora-Luna, F.; Páez-Rondón, O.; Aldana, E.; Sandoval, C.M. Orientation of Belminus Triatomines to Cockroaches and Cockroaches’ Fecal Volatiles: An Ethological Approach. acta ethologica 2021, 24, 53–66. [Google Scholar] [CrossRef]
- Lighton, J.R.; Fielden, L.J. Mass Scaling of Standard Metabolism in Ticks: A Valid Case of Low Metabolic Rates in Sit-and-Wait Strategists. Physiol. Zool. 1995, 6, 43–62. [Google Scholar] [CrossRef]
- Páez-Rondón, O.; Aldana, E.; Dickens, J.C.; Otálora-Luna, F. Ethological Description of a Fixed Action Pattern in a Kissing Bug (Triatominae): Vision, Gustation, Proboscis Extension and Drinking of Water and Guava. J. Ethol. 2018, 36, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Schulze, T.L.; Jordan, R.A.; Hung, R.W. Effects of Selected Meteorological Factors on Diurnal Questing of Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2001, 38, 318–324. [Google Scholar] [CrossRef]
- Brinkerhoff, R.J.; Clark, C.; Ocasio, K.; Gauthier, D.T.; Hynes, W.L. Factors Affecting the Microbiome of Ixodes scapularis and Amblyomma americanum. PLoS ONE 2020, 15, e0232398. [Google Scholar] [CrossRef]
- Klompen, J.S.H.; Black, W.; Keirans, J.E.; Oliver, J.H., Jr. Evolution of Ticks. Annu. Rev. Entomol. 1996, 41, 141–161. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Ixodes scapularis | |||
|---|---|---|---|
| Parameter | Control | Test (LED Lamp) | End-Control |
| Speed (mm/s) | 2.79 | 3.41 | 3.31 |
| Walking distance (mm) | 296.39 | 437.77 | 438.57 |
| Displacement (mm) | 129.40 | 184.24 | 181.71 |
| Linearity | 0.48 | 0.51 | 0.54 |
| Cosine of mean direction | −0.16 a | 0.51 b | 0.61 b |
| Upward straightness | 0.05 a | 0.57 b | 0.50 b |
| B. Amblyomma americanum | |||
| Parameter | Control | Test (LED Lamp) | End-Control |
| Speed (mm/s) | 5.67 | 6.80 | 4.86 |
| Walking distance (mm) | 680.61 | 807.01 | 579.04 |
| Displacement (mm) | 501.25 | 562.44 | 451.79 |
| Linearity | 0.67 | 0.67 | 0.57 |
| Cosine of mean direction | −0.41 a | 0.57 b | 0.37 b |
| Upward straightness | −0.27 a | 0.45 b | 0.15 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otálora-Luna, F.; Dickens, J.C.; Brinkerhoff, J.; Li, A.Y. Behavior of Nymphs and Adults of the Black-Legged Tick Ixodes scapularis and the Lone Star Tick Ambylomma americanum in Response to Thermal Stimuli. Insects 2022, 13, 130. https://doi.org/10.3390/insects13020130
Otálora-Luna F, Dickens JC, Brinkerhoff J, Li AY. Behavior of Nymphs and Adults of the Black-Legged Tick Ixodes scapularis and the Lone Star Tick Ambylomma americanum in Response to Thermal Stimuli. Insects. 2022; 13(2):130. https://doi.org/10.3390/insects13020130
Chicago/Turabian StyleOtálora-Luna, Fernando, Joseph C. Dickens, Jory Brinkerhoff, and Andrew Y. Li. 2022. "Behavior of Nymphs and Adults of the Black-Legged Tick Ixodes scapularis and the Lone Star Tick Ambylomma americanum in Response to Thermal Stimuli" Insects 13, no. 2: 130. https://doi.org/10.3390/insects13020130
APA StyleOtálora-Luna, F., Dickens, J. C., Brinkerhoff, J., & Li, A. Y. (2022). Behavior of Nymphs and Adults of the Black-Legged Tick Ixodes scapularis and the Lone Star Tick Ambylomma americanum in Response to Thermal Stimuli. Insects, 13(2), 130. https://doi.org/10.3390/insects13020130