
Phlebotomine Sand Flies in Southern Thailand: Entomological Survey, Identification of Blood Meals and Molecular Detection of Trypanosoma spp.

 ,
, 
 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
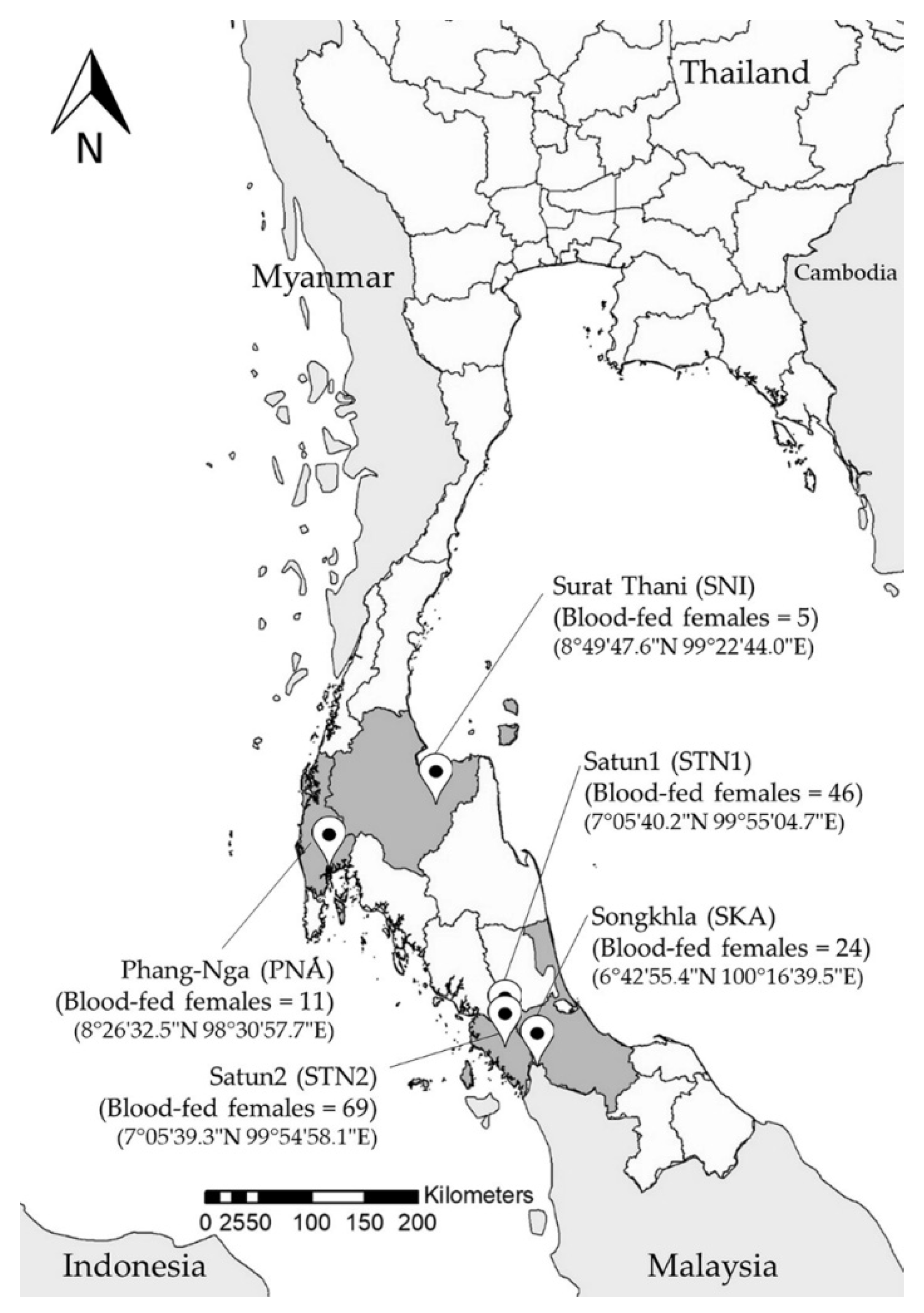
2.2. Sand Fly Collection
2.3. Sample Preparation
2.4. Morphological Identification of Sand Flies
2.5. Genomic DNA Extraction
2.6. Polymerase Chain Reaction (PCR) Analysis and DNA Sequencing
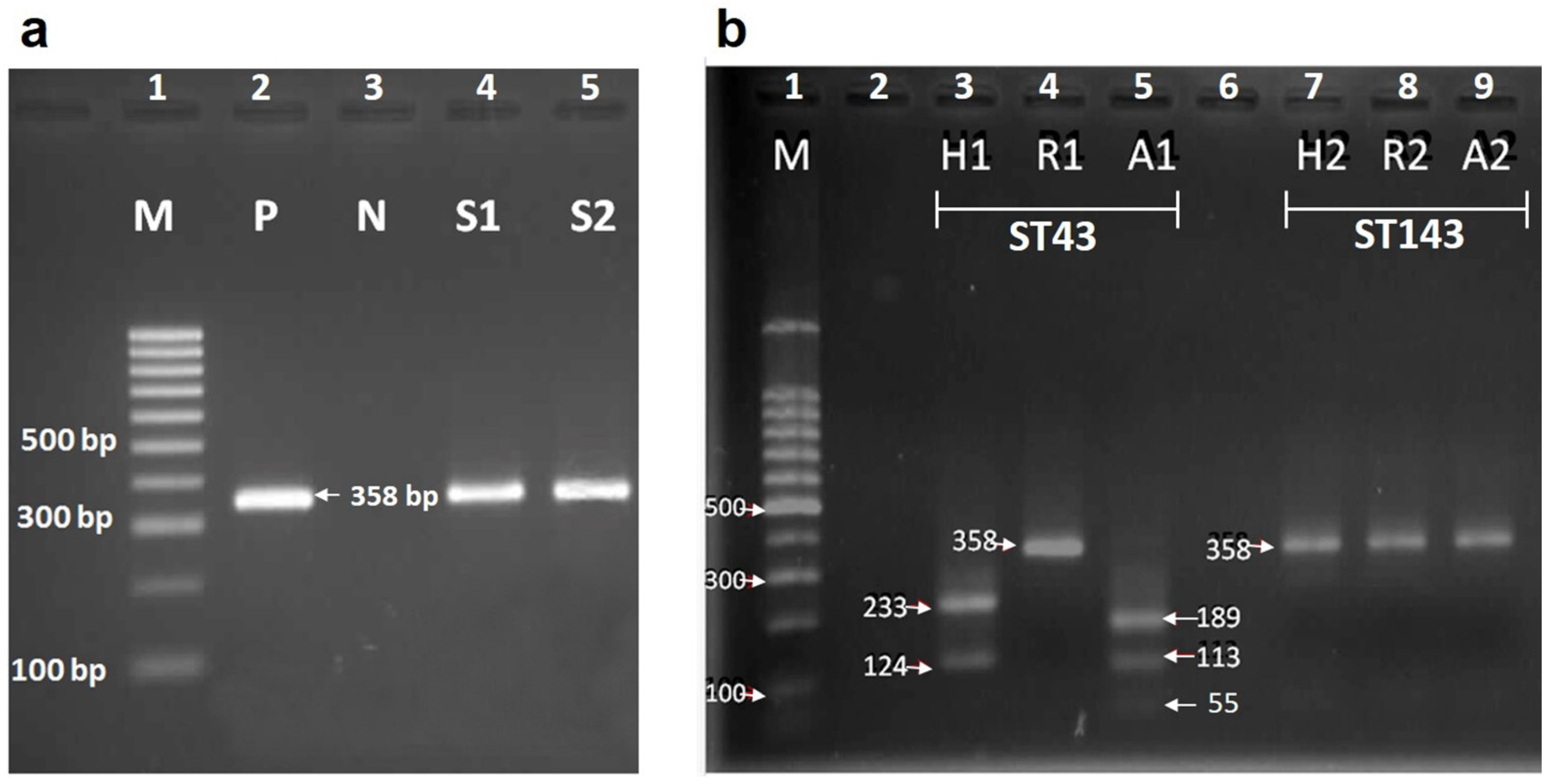
2.7. Identification of Blood Meal Source by PCR-RFLP
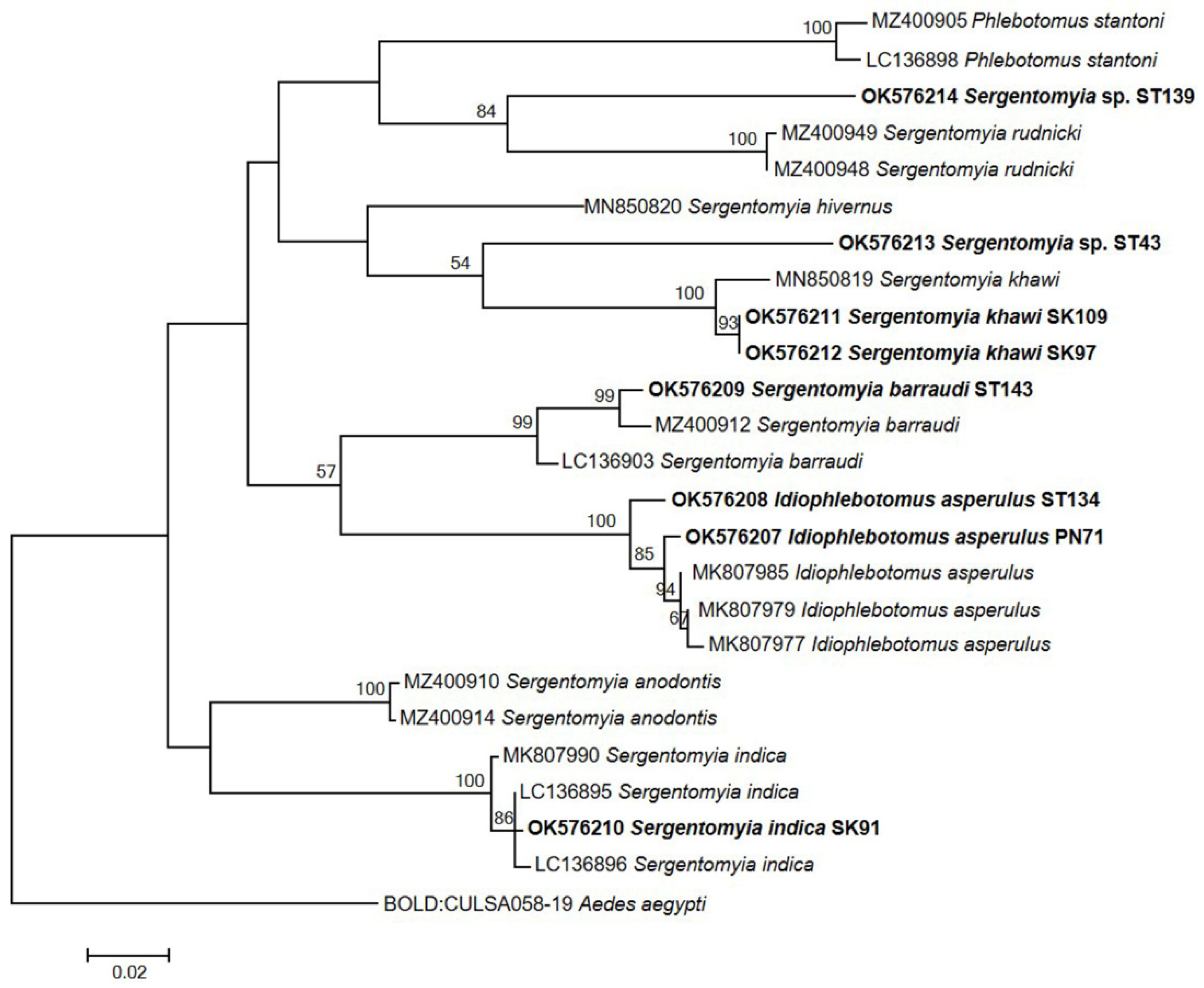
2.8. Phylogenetic Analysis of Sand Fly Species and Detected Parasites
3. Results
3.1. Collection and Morphological Identification of Sand Flies
3.2. Detection of Leishmania and Trypanosoma spp. in Sand Flies
3.3. Molecular Identification of Trypanosoma-Positive Sand Flies
3.4. Identification of Blood Source by RFLP
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rêgo, F.D.; Rugani, J.M.N.; Shimabukuro, P.H.F.; Tonelli, G.B.; Quaresma, P.F.; Gontijo, C.M.F. Molecular Detection of Leishmania in Phlebotomine Sand Flies (Diptera: Psychodidae) from a Cutaneous Leishmaniasis Focus at Xakriabá Indigenous Reserve, Brazil. PLoS ONE 2015, 10, e0122038. [Google Scholar] [CrossRef]
- Srisuton, P.; Phumee, A.; Sunantaraporn, S.; Boonserm, R.; Sor-Suwan, S.; Brownell, N.; Pengsakul, T.; Siriyasatien, P. Detection of Leishmania and Trypanosoma DNA in Field-Caught Sand Flies from Endemic and Non-Endemic Areas of Leishmaniasis in Southern Thailand. Insects 2019, 10, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, R.C.; De Souza, A.A.; Freitas, R.A.; Campaner, M.; Takata, C.S.A.; Barrett, T.V.; Shaw, J.J.; Teixeira, M.M.G. A Phylogenetic Lineage of Closely Related Trypanosomes (Trypanosomatidae, Kinetoplastida) of Anurans and Sand Flies (Psychodidae, Diptera) Sharing the Same Ecotopes in Brazilian Amazonia. J. Eukaryot. Microbiol. 2008, 55, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Rugna, G.; Clementi, E.; Carra, E.; Pinna, M.; Bergamini, F.; Fabbi, M.; Dottori, M.; Sacchi, L.; Votypka, J. Isolation of a Trypanosome Related to Trypanosoma theileri (Kinetoplastea: Trypanosomatidae) from Phlebotomus perfiliewi (Diptera: Psychodidae). BioMed Res. Int. 2018, 2018, 2597074. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Global Leishmaniasis Surveillance: 2019–2020, a Baseline for the 2030 Roadmap. Wkly. Epidemiol. Rec. 2021, 96, 401–419. [Google Scholar]
- Sarasombath, P.T. Leishmaniasis: An Evolving Public Health Concern in Thailand. Siriraj Med. J. 2018, 70, 363–376. [Google Scholar]
- Jariyapan, N.; Daroontum, T.; Jaiwong, K.; Chanmol, W.; Intakhan, N.; Sor-suwan, S.; Siriyasatien, P.; Somboon, P.; Bates, M.D.; Bates, P.A. Leishmania (Mundinia) orientalis n. sp. (Trypanosomatidae), a Parasite from Thailand Responsible for Localised Cutaneous Leishmaniasis. Parasit. Vectors 2018, 11, 351. [Google Scholar] [CrossRef]
- Kongkaew, W.; Siriarayaporn, P.; Leelayoova, S.; Supparatpinyo, K.; Areechokchai, D.; Duang-ngern, P.; Chanachai, K.; Sukmee, T.; Samung, Y.; Sridurongkathum, P. Autochthonous Visceral Leishmaniasis: A Report of a Second Case in Thailand. Southeast Asian J. Trop. Med. Public Health 2007, 38, 8–12. [Google Scholar]
- Maharom, P.; Siripattanapipong, S.; Mungthin, M.; Naaglor, T.; Sukkawee, R.; Pudkorn, R.; Wattana, W.; Wanachiwanawin, D.; Areechokchai, D.; Leelayoova, S. Visceral Leishmaniasis Caused by Leishmania infantum in Thailand. Southeast Asian J. Trop. Med. Public Health 2008, 39, 988–990. [Google Scholar]
- Bualert, L.; Charungkiattikul, W.; Thongsuksai, P.; Mungthin, M.; Siripattanapipong, S.; Khositnithikul, R.; Naaglor, T.; Ravel, C.; El Baidouri, F.; Leelayoova, S. Autochthonous Disseminated Dermal and Visceral Leishmaniasis in an AIDS Patient, Southern Thailand, Caused by Leishmania siamensis. Am. J. Trop. Med. Hyg. 2012, 86, 821–824. [Google Scholar] [CrossRef] [Green Version]
- Suankratay, C.; Suwanpimolkul, G.; Wilde, H.; Siriyasatien, P. Autochthonous Visceral Leishmaniasis in a Human Immunodeficiency Virus (HIV)-Infected Patient: The First in Thailand and Review of the Literature. Am. J. Trop. Med. Hyg. 2010, 82, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siriwardana, Y.; Deepachandi, B.; Gunasekara, C.; Warnasooriya, W.; Karunaweera, N.D. Leishmania donovani Induced Cutaneous Leishmaniasis: An Insight into Atypical Clinical Variants in Sri Lanka. J. Trop. Med. 2019, 4538597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svobodova, M.; Alten, B.; Zídková, L.; Dvorak, V.; Hlavacková, J.; Myskova, J.; Seblova, V.; Erisoz Kasap, O.; Belen, A.S.; Votypka, J.; et al. Cutaneous Leishmaniasis Caused by Leishmania infantum and Transmitted by Phlebotomus tobbi. Int. J. Parasitol. 2009, 39, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Chusri, S.; Hortiwakul, T.; Silpapojakul, K.; Siriyasatien, P. Consecutive Cutaneous and Visceral Leishmaniasis Manifestations Involving a Novel Leishmania Species in Two HIV Patients in Thailand. Am. J. Trop. Med. Hyg. 2012, 87, 76–80. [Google Scholar] [CrossRef]
- Alcover, M.M.; Rocamora, V.; Guillén, M.C.; Berenguer, D.; Cuadrado, M.; Riera, C.; Fisa, R. Case Report: Diffuse Cutaneous Leishmaniasis by Leishmania infantum in a Patient Undergoing Immunosuppressive Therapy: Risk Status in an Endemic Mediterranean Area. Am. J. Trop. Med. Hyg. 2018, 98, 1313–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pothirat, T.; Tantiworawit, A.; Chaiwarith, R.; Jariyapan, N.; Wannasan, A.; Siriyasatien, P.; Supparatpinyo, K.; Bates, M.D.; Kwakye-Nuako, G.; Bates, P.A. First Isolation of Leishmania from Northern Thailand: Case Report, Identification as Leishmania martiniquensis and Phylogenetic Position within the Leishmania enriettii Complex. PLoS Negl. Trop. Dis. 2014, 8, e3339. [Google Scholar] [CrossRef]
- Kanjanopas, K.; Siripattanapipong, S.; Ninsaeng, U.; Hitakarun, A.; Jitkaew, S.; Kaewtaphaya, P.; Tan-ariya, P.; Mungthin, M.; Charoenwong, C.; Leelayoova, S. Sergentomyia (Neophlebotomus) gemmea, a Potential Vector of Leishmania siamensis in Southern Thailand. BMC Infect. Dis. 2013, 13, 333. [Google Scholar] [CrossRef] [Green Version]
- Siripattanapipong, S.; Leelayoova, S.; Ninsaeng, U.; Mungthin, M. Detection of DNA of Leishmania siamensis in Sergentomyia (Neophlebotomus) iyengari (Diptera: Psychodidae) and Molecular Identification of Blood Meals of Sand Flies in an Affected Area, Southern Thailand. J. Med. Entomol. 2018, 55, 1277–1283. [Google Scholar] [CrossRef]
- Johnson, P.D. A Case of Infection by Trypanosoma lewisi in a Child. Trans. R. Soc. Trop. Med. Hyg. 1933, 26, 467–468. [Google Scholar] [CrossRef]
- Sarataphan, N.; Vongpakorn, M.; Nuansrichay, B.; Autarkool, N.; Keowkarnkah, T.; Rodtian, P.; Stich, R.W.; Jittapalapong, S. Diagnosis of a Trypanosoma lewisi-like (Herpetosoma) Infection in a Sick Infant from Thailand. J. Med. Microbiol. 2007, 56, 1118–1121. [Google Scholar] [CrossRef]
- Verma, A.; Manchanda, S.; Kumar, N.; Sharma, A.; Goel, M.; Banerjee, P.S.; Garg, R.; Singh, B.P.; Balharbi, F.; Lejon, V.; et al. Trypanosoma lewisi or T. lewisi-like Infection in a 37-Day-Old Indian Infant. Am. J. Trop. Med. Hyg. 2011, 85, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Vinh Chau, N.; Buu Chau, L.; Desquesnes, M.; Herder, S.; Phu Huong Lan, N.; Campbell, J.I.; Van Cuong, N.; Yimming, B.; Chalermwong, P.; Jittapalapong, S.; et al. A Clinical and Epidemiological Investigation of the First Reported Human Infection With the Zoonotic Parasite Trypanosoma evansi in Southeast Asia. Clin. Infect. Dis. 2016, 62, 1002–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikijkasemwat, K.; Kaewhom, P. Prevalence and Genetic Diversity of Trypanosoma evansi Infection Causing Abortion among Cattles and Buffaloes in Eastern Border Area of Thailand-Cambodia. Int. J. Agric. Technol. 2019, 15, 1021–1032. [Google Scholar]
- Kamyingkird, K.; Chalermwong, P.; Saechan, V.; Kaewnoi, D.; Desquesnes, M.; Ngasaman, R. Investigation of Trypanosoma evansi Infection in Bullfighting Cattle in Southern Thailand. Vet. World 2020, 13, 1674–1678. [Google Scholar] [CrossRef]
- Phumee, A.; Tawatsin, A.; Thavara, U.; Pengsakul, T.; Thammapalo, S.; Depaquit, J.; Gay, F.; Siriyasatien, P. Detection of an Unknown Trypanosoma DNA in a Phlebotomus stantoni (Diptera: Psychodidae) Collected From Southern Thailand and Records of New Sand Flies With Reinstatement of Sergentomyia hivernus Raynal & Gaschen, 1935 (Diptera: Psychodidae). J. Med. Entomol. 2017, 54, 429–434. [Google Scholar] [CrossRef]
- Alexandre, J.; Sadlova, J.; Lestinova, T.; Vojtkova, B.; Jancarova, M.; Podesvova, L.; Yurchenko, V.; Dantas-Torres, F.; Brandão-Filho, S.P.; Volf, P. Experimental Infections and Co-Infections with Leishmania braziliensis and Leishmania infantum in Two Sand Fly Species, Lutzomyia migonei and Lutzomyia longipalpis. Sci. Rep. 2020, 10, 3566. [Google Scholar] [CrossRef]
- Rogers, M.; Chance, M.; Bates, P. The Role of Promastigote Secretory Gel in the Origin and Transmission of the Infective Stage of Leishmania mexicana by the Sandfly Lutzomyia longipalpis. Parasitology 2002, 124, 495–507. [Google Scholar] [CrossRef] [Green Version]
- De Colmenares, M.; Portús, M.; Botet, J.; Dobaño, C.; Gállego, M.; Wolff, M.; Seguí, G. Identification of Blood Meals of Phlebotomus perniciosus (Diptera: Psychodidae) in Spain by a Competitive Enzyme-Linked Immunosorbent Assay Biotin/Avidin Method. J. Med. Entomol. 1995, 32, 229–233. [Google Scholar] [CrossRef]
- Bongiorno, G.; Habluetzel, A.; Khoury, C.; Maroli, M. Host Reference of Phlebotomine Sand Flies at a Hypodermic Focus of Canine Leishmaniosis in Central Italy. Acta Trop. 2003, 88, 109–116. [Google Scholar] [CrossRef]
- Kent, R.J. Molecular Methods for Arthropod Bloodmeal Identification and Applications to Ecological and Vector-Borne Disease Studies. Mol. Ecol. Resour. 2009, 9, 4–18. [Google Scholar] [CrossRef]
- Soares, V.Y.R.; da Silva, J.C.; da Silva, K.R.; Pires e Cruz, M.D.S.; Santos, M.P.D.; Ribolla, P.E.M.; Alonso, D.P.; Coelho, L.F.L.; Costa, D.L.; Costa, C.H.N. Identification of Blood Meal Sources of Lutzomyia longipalpis Using Polymerase Chain Reaction-Restriction Fragment Length Polymorphism Analysis of the Cytochrome B Gene. Mem. Inst. Oswaldo Cruz 2014, 109, 379–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaresma, P.F.; Carvalho, G.M.; Ramos, M.C.; Andrade Filho, J.D. Natural Leishmania sp. Reservoirs and Phlebotomine Sandfly Food Source Identification in Ibitipoca State Park, Minas Gerais, Brazil. Mem. Inst. Oswaldo Cruz 2012, 107, 480–485. [Google Scholar] [CrossRef] [PubMed]
- González, E.; Gállego, M.; Molina, R.; Abras, A.; Alcover, M.M.; Ballart, C.; Fernández, A.; Jiménez, M. Identification of Blood Meals in Field Captured Sand Flies by a PCR-RFLP Approach Based on Cytochrome b Gene. Acta Trop. 2015, 152, 96–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, L.E. Hoyer’s Solution as a Rapid Permanent Mounting Medium for Bryophytes. Bryol. 1954, 57, 242–244. [Google Scholar] [CrossRef]
- Lewis, D.J. The Phlebotomine Sandflies (Diptera: Psychodidae) of the Oriental Region. Bull. Br. Mus. Nat. Hist. B Entomol. 1978, 37, 217–343. [Google Scholar] [CrossRef]
- Lane, R.P. Sandflies (Phlebotominae). In Medical Insects and Arachnids; Lane, R.P., Crosskey, R.W., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 78–119. [Google Scholar] [CrossRef]
- Abonnenc, E. Les Phlébotomes de la Région Éthiopienne (Diptera, Psychodidae); Memoirs ORSTOM: Paris, France, 1972; Volume 1. [Google Scholar]
- Kakarsulemankhel, J. Taxonomic Morphology of Sergentomyia (Neophlebotomus) hodgsoni hodgsoni Sinton (1933) (Diptera, Psychodidae) from Pakistan. Pak. J. Biol. Sci. 2004, 7, 1148–1153. [Google Scholar] [CrossRef]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D.N. DNA Barcodes Distinguish Species of Tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef] [Green Version]
- Spanakos, G.; Piperaki, E.-T.; Menounos, P.G.; Tegos, N.; Flemetakis, A.; Vakalis, N.C. Detection and Species Identification of Old World Leishmania in Clinical Samples Using a PCR-Based Method. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 46–53. [Google Scholar] [CrossRef]
- Oshaghi, M.A.; Chavshin, A.R.; Vatandoost, H. Analysis of Mosquito Bloodmeals Using RFLP Markers. Exp. Parasitol. 2006, 114, 259–264. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apiwathnasorn, C.; Samung, Y.; Prummongkol, S.; Phayakaphon, A.; Panasopolkul, C. Cavernicolous Species of Phlebotomine Sand Flies from Kanchanaburi Province, with an Updated Species List for Thailand. Southeast Asian J. Trop. Med. Public Health 2011, 42, 1405–1409. [Google Scholar] [PubMed]
- Depaquit, J.; Vongphayloth, K.; Siriyasatien, P.; Polseela, R.; Phumee, A.; Loyer, M.; Vol, A.; Varlot, G.; Rahola, N.; Brey, P.T.; et al. On the True Identity of Sergentomyia gemmea and Description of a Closely Related Species: Se. raynali n. sp. Med. Vet. Entomol. 2019, 33, 521–529. [Google Scholar] [CrossRef]
- Borland, E.M.; Kading, R.C. Modernizing the Toolkit for Arthropod Bloodmeal Identification. Insects 2021, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Hlavackova, K.; Dvorak, V.; Chaskopoulou, A.; Volf, P.; Halada, P. A Novel MALDI-TOF MS-Based Method for Blood Meal Identification in Insect Vectors: A Proof of Concept Study on Phlebotomine Sand Flies. PLoS Negl. Trop. Dis. 2019, 13, e0007669. [Google Scholar] [CrossRef]
- Chusri, S.; Thammapalo, S.; Chusri, S.; Thammapalo, S.; Silpapojakul, K.; Siriyasatien, P. Animal Reservoirs and Potential Vectors of Leishmania siamensis in Southern Thailand. Southeast Asian J. Trop. Med. Public Health 2014, 45, 13–19. [Google Scholar]
- Panthawong, A.; Chareonviriyaphap, T.; Phasuk, J. Species Diversity and Seasonality of Phlebotmomine Sand Flies (Diptera: Psychodidae) in Satun Province, Thailand. Southeast Asian J. Trop. Med. Public Health 2015, 46, 857–865. [Google Scholar]
- Kato, H.; Uezato, H.; Sato, H.; Bhutto, A.M.; Soomro, F.R.; Baloch, J.H.; Iwata, H.; Hashiguchi, Y. Natural Infection of the Sand Fly Phlebotomus kazeruni by Trypanosoma Species in Pakistan. Parasit. Vectors 2010, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Nzelu, C.O.; Kato, H.; Puplampu, N.; Desewu, K.; Odoom, S.; Wilson, M.D.; Sakurai, T.; Katakura, K.; Boakye, D.A. First Detection of Leishmania tropica DNA and Trypanosoma Species in Sergentomyia Sand Flies (Diptera: Psychodidae) from an Outbreak Area of Cutaneous Leishmaniasis in Ghana. PLoS Negl. Trop. Dis. 2014, 8, e2630. [Google Scholar] [CrossRef] [Green Version]
- Molee, P.W.; Sakulsak, N.; Saengamnatdej, S. Detection of Trypanosoma spp. in Bandicota Indica from the Thai-Myanmar Border Area, Mae Sot District Tak Province, Thailand. Asian Pac. J. Trop. Med. 2019, 12, 457–462. [Google Scholar] [CrossRef]
- Pumhom, P.; Pognon, D.; Yangtara, S.; Thaprathorn, N.; Milocco, C.; Douangboupha, B.; Herder, S.; Chaval, Y.; Morand, S.; Jittapalapong, S.; et al. Molecular Prevalence of Trypanosoma spp. In Wild Rodents of Southeast Asia: Influence of Human Settlement Habitat. Epidemiol. Infect. 2014, 142, 1221–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Meal Sources | Fragments Size (bp) | ||
|---|---|---|---|
| Hae III | Rsa I | Aci I | |
| Homo sapiens (Human) | 233, 124 | 358 | 189, 113, 55 |
| Canis familiaris (Dog) | 358 | 358 | 358 |
| Rattus novergicus (Rats) | 358 | 267, 59, 31 | 358 |
| Felis catus (Domestic cats) | 272, 74, 11 | 214, 119, 24 | 244, 113 |
| Sus domesticus (Swine) | 153, 130, 74 | 358 | 358 |
| Bus Taurus (Cattle) | 159, 124, 74 | 322, 31, 4 | 358 |
| Gallus gallus (Chicken) | 159, 124, 74 | 208, 149 | 308, 49 |
| Equus caballus (Horse) | 159, 124, 74 | 358 | 244, 113 |
| Sand Fly Species | No. of Sand Flies in Collection Sites | Blood Meal Source | |||||||
|---|---|---|---|---|---|---|---|---|---|
| SKA | SNI | STN1 | STN2 | PNA | Total | Human | Dog | Rat | |
| Id. asperulus | 0 | 2 | 5 (1) | 5 | 7 (1) | 19 | 3 | 0 | 0 |
| P. argentipes | 1 | 0 | 1 | 1 | 0 | 3 | 2 | 0 | 1 |
| P. stantoni | 2 | 0 | 1 | 1 | 0 | 4 | 0 | 0 | 0 |
| S. anodontis | 0 | 3 | 8 | 8 | 1 | 20 | 2 | 0 | 1 |
| S. barraudi | 2 | 0 | 5 (2) | 2 | 0 | 9 | 2 | 1 (1) | 0 |
| S. hamidi | 0 | 0 | 19 | 4 | 0 | 23 | 7 | 0 | 0 |
| S. hodgsoni | 0 | 0 | 2 | 1 | 1 | 4 | 2 | 0 | 0 |
| S. hodgsoni hodgsoni | 0 | 0 | 0 | 32 | 0 | 32 | 4 | 8 | 11 |
| S. indica | 4 (1) | 0 | 0 | 1 | 0 | 5 | 0 | 0 | 0 |
| S. iyengari | 0 | 0 | 1 | 1 | 0 | 2 | 0 | 0 | 0 |
| S. khawi | 14 (2) | 0 | 2 | 0 | 1 | 17 | 2 | 0 | 0 |
| S. silvatica | 0 | 0 | 1 | 10 | 0 | 11 | 7 | 0 | 1 |
| Sergentomyia sp. | 1 | 0 | 1 | 3 (1) | 1 | 6 | 2 (1) | 0 | 0 |
| Total | 24 | 5 | 46 | 69 | 11 | 155 | 33 | 9 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buatong, J.; Dvorak, V.; Thepparat, A.; Thongkhao, K.; Koyadun, S.; Siriyasatien, P.; Pengsakul, T. Phlebotomine Sand Flies in Southern Thailand: Entomological Survey, Identification of Blood Meals and Molecular Detection of Trypanosoma spp. Insects 2022, 13, 197. https://doi.org/10.3390/insects13020197
Buatong J, Dvorak V, Thepparat A, Thongkhao K, Koyadun S, Siriyasatien P, Pengsakul T. Phlebotomine Sand Flies in Southern Thailand: Entomological Survey, Identification of Blood Meals and Molecular Detection of Trypanosoma spp. Insects. 2022; 13(2):197. https://doi.org/10.3390/insects13020197
Chicago/Turabian StyleBuatong, Jirayu, Vit Dvorak, Arunrat Thepparat, Kanaphot Thongkhao, Surachart Koyadun, Padet Siriyasatien, and Theerakamol Pengsakul. 2022. "Phlebotomine Sand Flies in Southern Thailand: Entomological Survey, Identification of Blood Meals and Molecular Detection of Trypanosoma spp." Insects 13, no. 2: 197. https://doi.org/10.3390/insects13020197
APA StyleBuatong, J., Dvorak, V., Thepparat, A., Thongkhao, K., Koyadun, S., Siriyasatien, P., & Pengsakul, T. (2022). Phlebotomine Sand Flies in Southern Thailand: Entomological Survey, Identification of Blood Meals and Molecular Detection of Trypanosoma spp. Insects, 13(2), 197. https://doi.org/10.3390/insects13020197