Intraspecific Relationships and Nest Mound Shape Are Affected by Habitat Features in Introduced Populations of the Red Wood Ant Formica paralugubris

 , , ,
, , , 
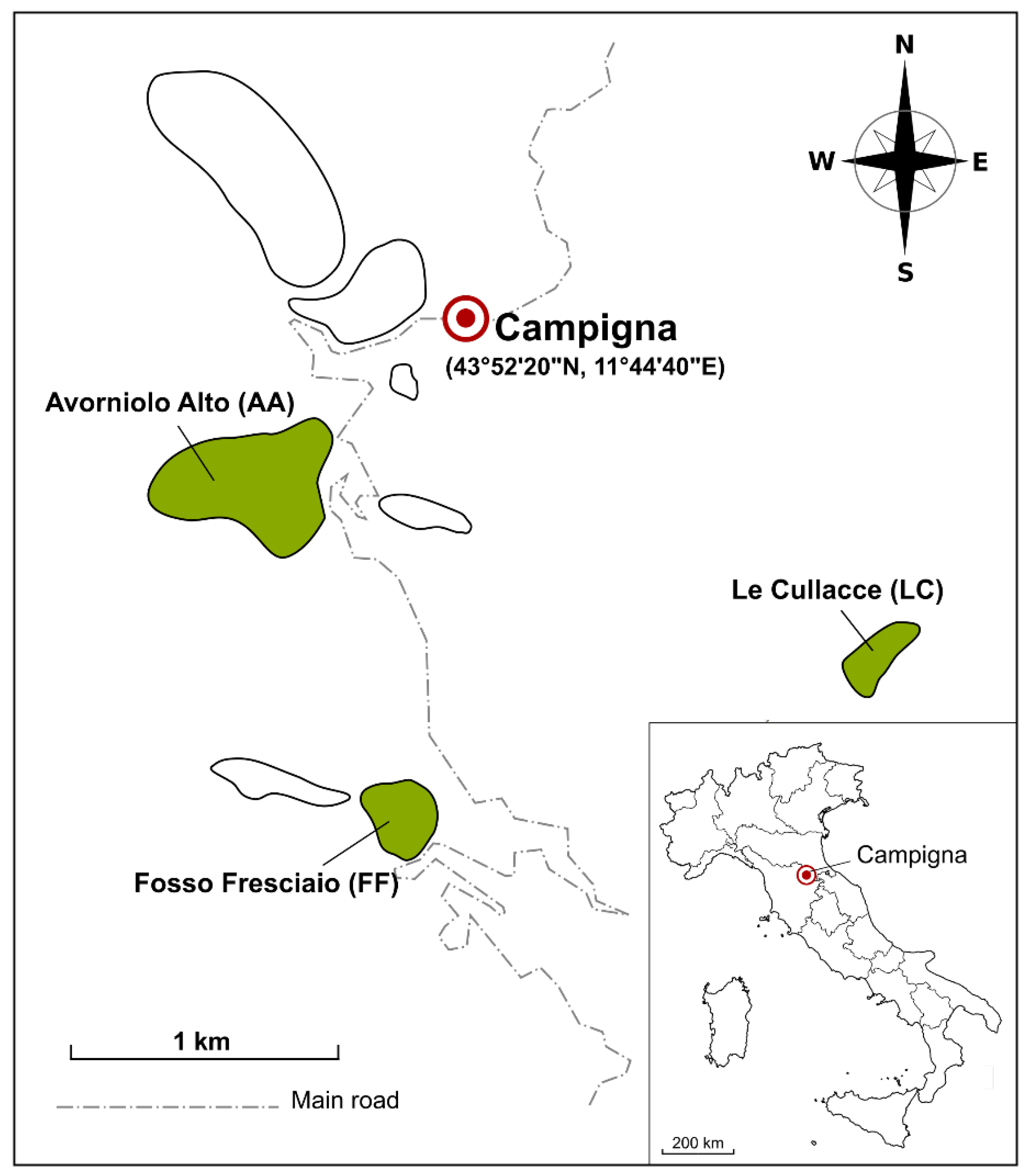{kind=link}
{kind=link}
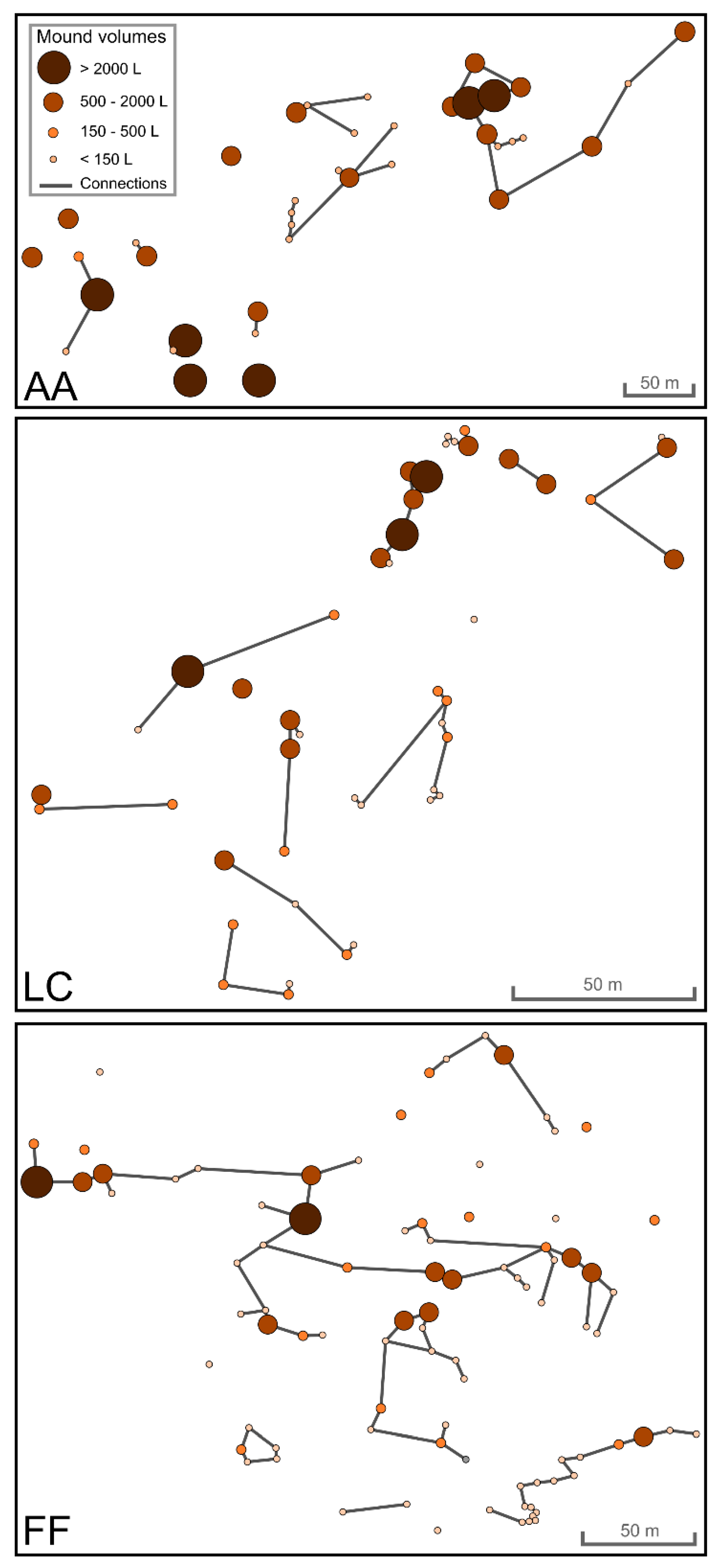{kind=link}
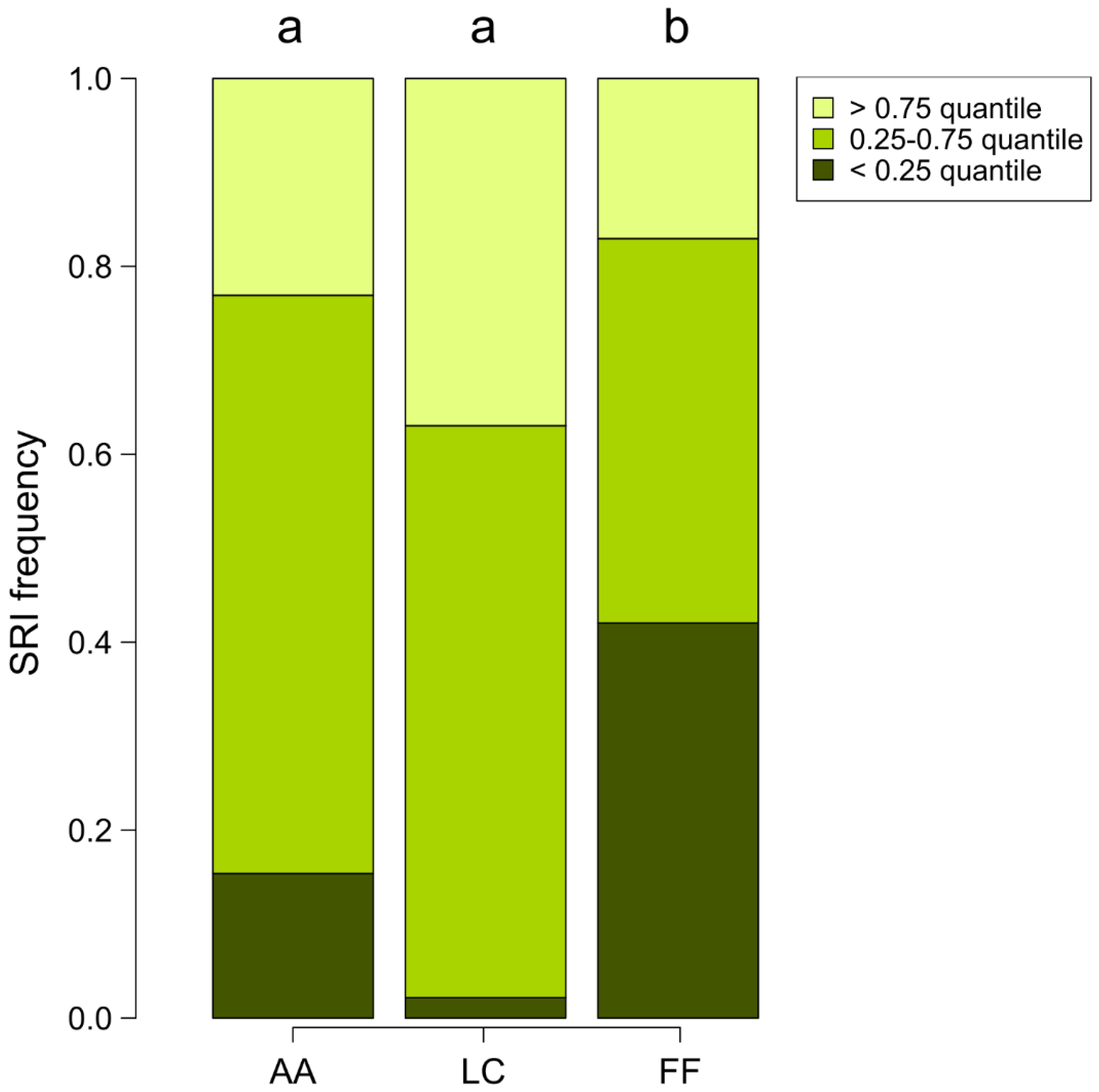{kind=link}
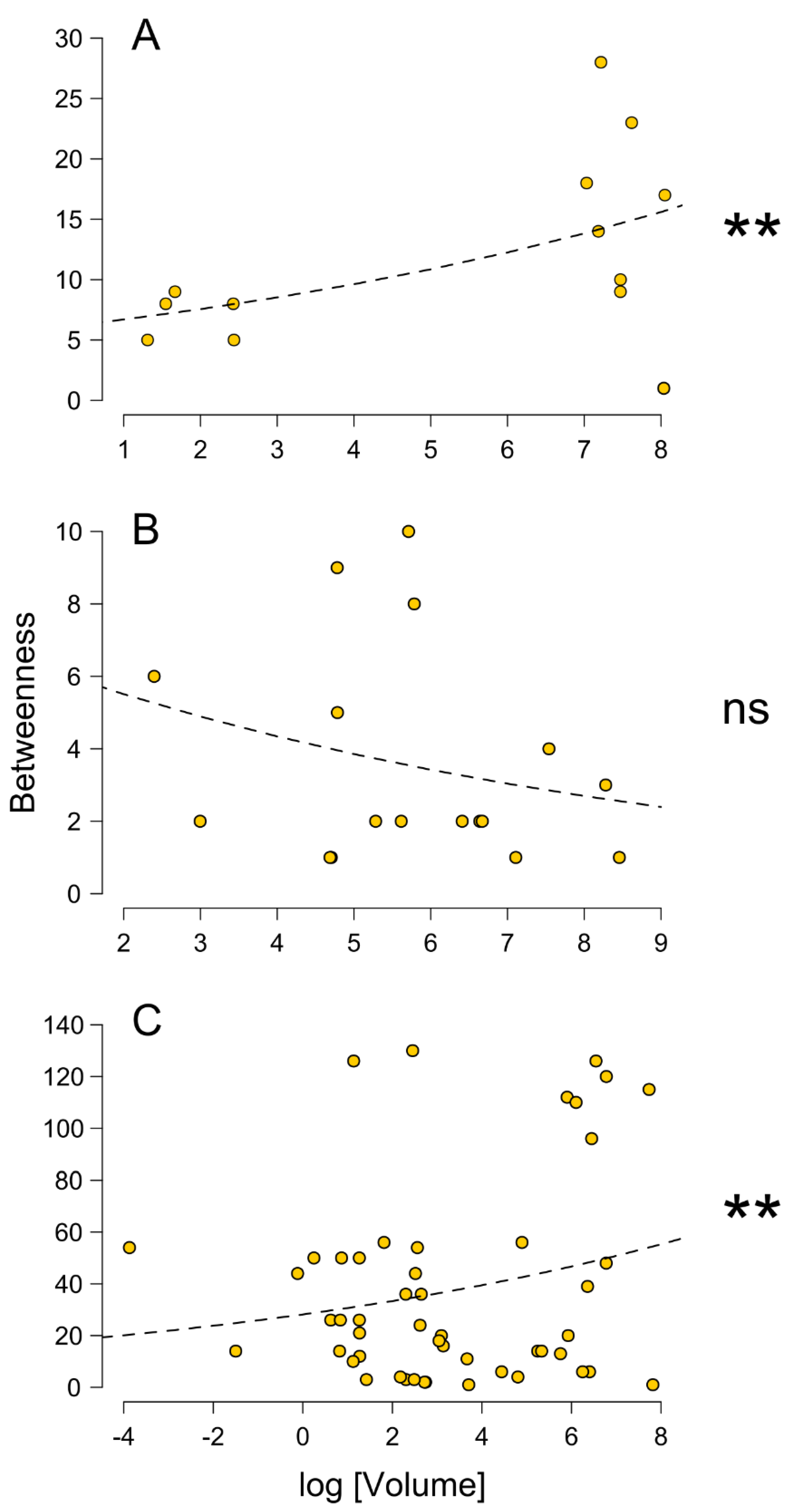{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stockan, J.; Robinson, E.J.H.; Trager, J.C.; Yao, I.; Seifert, B. Introducing Wood Ants: Evolution, Phylogeny, Identification and Distribution. In Wood Ant Ecology and Conservation; Cambridge University Press: Cambridge, UK, 2016; pp. 1–36. [Google Scholar]
- Rosengren, R.; Fortelius, W.; Lindström, K.; Luther, A. Phenology and Causation of Nest Heating and Thermoregulation in Red Wood Ants of the Formica Rufa Group Studied in Coniferous Forest Habitats in Southern Finland. Ann. Zool. Fenn. 1987, 24, 147–155. [Google Scholar]
- Frouz, J.; Kalčík, J.; Cudlín, P. Accumulation of phosphorus in nests of red wood ants Formica s. str. Ann. Zool. Fenn. 2005, 42, 269–275. [Google Scholar]
- Risch, A.C.; Jurgensen, M.F.; Schütz, M.; Page-Dumroese, D.S. The Contribution of Red Wood Ants to Soil C and N Pools and CO2 Emissions in Subalpine Forests. Ecology 2005, 86, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, M.; Kilpeläinen, J.; Finér, L.; Risch, A.C.; Domisch, T.; Neuvonen, S.; Niemelä, P. The Effect of Red Wood Ant (Formica rufa Group) Mounds on Root Biomass, Density, and Nutrient Concentrations in Boreal Managed Forests. J. For. Res. 2007, 12, 113–119. [Google Scholar] [CrossRef]
- Balzani, P.; Masoni, A.; Venturi, S.; Frizzi, F.; Bambi, M.; Fani, R.; Nisi, B.; Tassi, F.; Vaselli, O.; Zaccaroni, M.; et al. CO2 Biogeochemical Investigation and Microbial Characterization of Red Wood Ant Mounds in a Southern Europe Montane Forest. Soil Biol. Biochem. 2022, 166, 108536. [Google Scholar] [CrossRef]
- Domisch, T.; Risch, A.C.; Robinson, E.J.H. Wood Ant Foraging and Mutualism with Aphids. In Wood Ant Ecology and Conservation; Stockan, J.A., Robinson, E.J.H., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 145–176. ISBN 978-1-107-26140-2. [Google Scholar]
- Risch, A.C.; Ellis, S.; Wiswell, H. Where and Why? Wood Ant Population Ecology. In Wood Ant Ecology and Conservation; Cambridge University Press: Cambridge, UK, 2016; pp. 81–105. [Google Scholar]
- Vandegehuchte, M.L.; Wermelinger, B.; Fraefel, M.; Baltensweiler, A.; Düggelin, C.; Brändli, U.-B.; Freitag, A.; Bernasconi, C.; Cherix, D.; Risch, A.C. Distribution and Habitat Requirements of Red Wood Ants in Switzerland: Implications for Conservation. Biol. Conserv. 2017, 212, 366–375. [Google Scholar] [CrossRef]
- Debout, G.; Schatz, B.; Elias, M.; Mckey, D. Polydomy in Ants: What We Know, What We Think We Know, and What Remains to Be Done. Biol. J. Linn. Soc. 2007, 90, 319–348. [Google Scholar] [CrossRef] [Green Version]
- Ellis, S.; Robinson, E.J.H. Polydomy in Red Wood Ants. Insectes Sociaux 2014, 61, 111–122. [Google Scholar] [CrossRef]
- Sorvari, J.; Hakkarainen, H. Habitat-Related Aggressive Behaviour between Neighbouring Colonies of the Polydomous Wood Ant Formica Aquilonia. Anim. Behav. 2004, 67, 151–153. [Google Scholar] [CrossRef]
- Ellis, S.; Franks, D.W.; Robinson, E.J.H. Resource Redistribution in Polydomous Ant Nest Networks: Local or Global? Behav. Ecol. 2014, 25, 1183–1191. [Google Scholar] [CrossRef]
- Bernasconi, C.; Maeder, A.; Cherix, D.; Pamilo, P. Diversity and Genetic Structure of the Wood Ant Formica Lugubris in Unmanaged Forests. Ann. Zool. Fenn. 2005, 42, 189–199. [Google Scholar]
- Buhl, J.; Hicks, K.; Miller, E.R.; Persey, S.; Alinvi, O.; Sumpter, D.J. Shape and Efficiency of Wood Ant Foraging Networks. Behav. Ecol. Sociobiol. 2009, 63, 451–460. [Google Scholar] [CrossRef]
- Burns, D.D.; Franks, D.W.; Parr, C.; Robinson, E.J.H. Ant Colony Nest Networks Adapt to Resource Disruption. J. Anim. Ecol. 2021, 90, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, B. Formica Paralugubris Nov. Spec.—A Sympatric Sibling Species of Formica lugubris from the Western Alps (Insecta: Hymenoptera: Formicoidea: Formicidae. Reichenbachia 1996, 35, 193–201. [Google Scholar]
- Chapuisat, M.; Keller, L. Extended Family Structure in the Ant Formica paralugubris: The Role of the Breeding System. Behav. Ecol. Sociobiol. 1999, 46, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Chapuisat, M.; Bernasconi, C.; Hoehn, S.; Reuter, M. Nestmate recognition in the unicolonial ant Formica paralugubris. Behav. Ecol. 2005, 16, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Helanterä, H.; Strassmann, J.E.; Carrillo, J.; Queller, D.C. Unicolonial Ants: Where Do They Come from, What Are They and Where Are They Going? Trends Ecol. Evol. 2009, 24, 341–349. [Google Scholar] [CrossRef]
- Pavan, M. Attività Italiana per la Lotta Biologica con Formiche del Gruppo Formica rufa Contro Gli Insetti Dannosi Alle Foreste; Collana Verde, Ministero dell’Agricoltura e Delle Foreste: Rome, Italy, 1959. [Google Scholar]
- Masoni, A.; Frizzi, F.; Natali, C.; Bernasconi, C.; Ciofi, C.; Santini, G. Molecular identification of imported red wood ant populations in the Campigna biogenetic nature Reserve (foreste Casentinesi national Park, Italy). Conserv. Genet. Resour. 2019, 11, 231–236. [Google Scholar] [CrossRef]
- Balzani, P.; Vizzini, S.; Frizzi, F.; Masoni, A.; Lessard, J.-P.; Bernasconi, C.; Francoeur, A.; Ibarra-Isassi, J.; Brassard, F.; Cherix, D.; et al. Plasticity in the Trophic Niche of an Invasive Ant Explains Establishment Success and Long-term Coexistence. Oikos 2021, 130, 691–696. [Google Scholar] [CrossRef]
- Ronchetti, G.; Mazzoldi, P.; Groppali, R. Venticinque Anni di Osservazioni sui Trapianti di Formica lugubris zett. dalle Alpi alle Foreste Demaniali Casentinesi, Italia Centrale: Hymen Formicidae; Università di Pavia: Pavia, Italy, 1986. (In Italian) [Google Scholar]
- Frizzi, F.; Masoni, A.; Quilghini, G.; Ciampelli, P.; Santini, G. Chronicle of an Impact Foretold: The Fate and Effect of the Introduced Formica paralugubris Ant. Biol. Invasions 2018, 20, 3575–3589. [Google Scholar] [CrossRef]
- Frizzi, F.; Masoni, A.; Migliorini, M.; Fanciulli, P.P.; Cianferoni, F.; Balzani, P.; Giannotti, S.; Davini, G.; Wendt, C.F.; Santini, G. A Comparative Study of the Fauna Associated with Nest Mounds of Native and Introduced Populations of the Red Wood Ant Formica paralugubris. Eur. J. Soil Biol. 2020, 101, 103241. [Google Scholar] [CrossRef]
- Dekoninck, W.; Maebe, K.; Breyne, P.; Hendrickx, F. Polygyny and Strong Genetic Structuring within an Isolated Population of the Wood Ant Formica rufa. J. Hymenopt. Res. 2014, 41, 95. [Google Scholar] [CrossRef] [Green Version]
- Sondej, I.; Domisch, T.; Finér, L.; Czechowski, W. Wood Ants Prefer Conifers to Broadleaved Trees in Mixed Temperate Forests. Agric. For. Entomol. 2021, 23, 287–296. [Google Scholar] [CrossRef]
- Frączek, J.; Złobecki, A.; Zemanek, J. Assessment of Angle of Repose of Granular Plant Material Using Computer Image Analysis. J. Food Eng. 2007, 83, 17–22. [Google Scholar] [CrossRef]
- Blonder, B.; Wey, T.W.; Dornhaus, A.; James, R.; Sih, A. Temporal Dynamics and Network Analysis. Methods Ecol. Evol. 2012, 3, 958–972. [Google Scholar] [CrossRef]
- Cook, Z.; Franks, D.W.; Robinson, E.J.H. Efficiency and Robustness of Ant Colony Transportation Networks. Behav. Ecol. Sociobiol. 2014, 68, 509–517. [Google Scholar] [CrossRef]
- Batchelor, T.P.; Santini, G.; Briffa, M. Size Distribution and Battles in Wood Ants: Group Resource-Holding Potential Is the Sum of the Individual Parts. Anim. Behav. 2012, 83, 111–117. [Google Scholar] [CrossRef]
- Ellis, S.; Franks, D.W.; Robinson, E.J.H. Ecological consequences of colony structure in dynamic ant nest networks. Ecol. Evol. 2017, 7, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Van Wilgenburg, E.; Elgar, M.A. Confirmation Bias in Studies of Nestmate Recognition: A Cautionary Note for Research into the Behaviour of Animals. PLoS ONE 2013, 8, e53548. [Google Scholar] [CrossRef] [Green Version]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2002; ISBN 978-0-521-81128-6. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Sorvari, J.; Elo, R.A.; Härkönen, S.K. Forest-Built Nest Mounds of Red Wood Ant Formica Aquilonia Are No Good in Clear Fells. Appl. Soil Ecol. 2016, 101, 101–106. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Robinson, E.J.H. The Relationship between Canopy Cover and Colony Size of the Wood Ant Formica Lugubris-Implications for the Thermal Effects on a Keystone Ant Species. PLoS ONE 2014, 9, e116113. [Google Scholar] [CrossRef] [PubMed]
- Punttila, P.; Kilpeläinen, J. Distribution of Mound-Building Ant Species (Formica Spp., Hymenoptera) in Finland: Preliminary Results of a National Survey. Ann. Zool. Fenn. 2009, 46, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sorvari, J.; Haatanen, M.K.; Vesterlund, S.R. Combined Effects of Overwintering Temperature and Habitat Degradation on the Survival of Boreal Wood Ant. J. Insect Conserv. 2011, 15, 727–731. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Robinson, E.J.H. A Comparison of Mark—Release—Recapture Methods for Estimating Colony Size in the Wood Ant Formica lugubris. Insectes Sociaux 2013, 60, 351–359. [Google Scholar] [CrossRef]
- Moyano, F.E.; Kutsch, W.L.; Rebmann, C. Soil Respiration Fluxes in Relation to Photosynthetic Activity in Broad-Leaf and Needle-Leaf Forest Stands. Agric. For. Meteorol. 2008, 148, 135–143. [Google Scholar] [CrossRef]
- Burns, D.D.R.; Franks, D.W.; Parr, C.; Hawke, C.; Ellis, S.; Robinson, E.J.H. A longitudinal study of nest occupancy, trail networks and foraging in a polydomous wood ant population. Insectes Sociaux 2020, 67, 419–427. [Google Scholar] [CrossRef]
- Wuorenrinne, H. Effects of urban pressure on colonies of Formica rufa group (Hymenoptera, Formicidae) in the town of Espoo (Finland). Ann. Zool. 1989, 42, 13–17. [Google Scholar]
- Eeva, T.; Sorvari, J.; Koivunen, V. Effects of Heavy Metal Pollution on Red Wood Ant (Formica s. Str.) Populations. Environ. Pollut. 2004, 132, 533–539. [Google Scholar] [CrossRef]
- Herbers, J.M.; Banschbach, V.S. Plasticity of social organization in a forest ant species. Behav. Ecol. Sociobiol. 1999, 45, 451–465. [Google Scholar] [CrossRef]
- Lanan, M.C.; Dornhaus, A.; Bronstein, J.L. The Function of Polydomy: The Ant Crematogaster torosa Preferentially Forms New Nests near Food Sources and Fortifies Outstations. Behav. Ecol. Sociobiol. 2011, 65, 959–968. [Google Scholar] [CrossRef]
- Buczkowski, G.; Bennett, G. Colony Budding and Its Effects on Food Allocation in the Highly Polygynous Ant, Monomorium pharaonis. Ethology 2009, 115, 1091–1099. [Google Scholar] [CrossRef]
- Groppali, R.; Omati, M.; Piccardi, B. Preferenze ambientali di Formica lugubris zett. (Hymenoptera: Formicidae) nelle Prealpi Italiane: Indagine nella Riserva Naturale Biogenetica “Giovetto di Palline. Ann. Mus. Civ. Stor. Nat. Brescia 2003, 33, 81–88. [Google Scholar]
- Robinson, E.J.H. Polydomy: The Organisation and Adaptive Function of Complex Nest Systems in Ants. Curr. Opin. Insect Sci. 2014, 5, 37–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorvari, J.; Hakkarainen, H. Deforestation Reduces Nest Mound Size and Decreases the Production of Sexual Offspring in the Wood Ant Formica aquilonia. Ann. Zool. Fenn. 2005, 42, 259–267. [Google Scholar]
- Holzer, B.; Chapuisat, M.; Kremer, N.; Finet, C.; Keller, L. Unicoloniality, Recognition and Genetic Differentiation in a Native Formica Ant. J. Evol. Biol. 2006, 19, 2031–2039. [Google Scholar] [CrossRef] [Green Version]
- Christensen, C.; Radford, A.N. Dear Enemies or Nasty Neighbors? Causes and Consequences of Variation in the Responses of Group-Living Species to Territorial Intrusions. Behav. Ecol. 2018, 29, 1004–1013. [Google Scholar] [CrossRef] [Green Version]
- Newey, P.S.; Robson, S.K.; Crozier, R.H. Weaver Ants Oecophylla Smaragdina Encounter Nasty Neighbors Rather than Dear Enemies. Ecology 2010, 91, 2366–2372. [Google Scholar] [CrossRef]
- Frizzi, F.; Ciofi, C.; Dapporto, L.; Natali, C.; Chelazzi, G.; Turillazzi, S.; Santini, G. The Rules of Aggression: How Genetic, Chemical and Spatial Factors Affect Intercolony Fights in a Dominant Species, the Mediterranean Acrobat Ant Crematogaster scutellaris. PLoS ONE 2015, 10, e0137919. [Google Scholar] [CrossRef] [Green Version]
- Benedek, K.; Kobori, O.T. “Nasty Neighbour” Effect in Formica pratensis Retz.(Hymenoptera: Formicidae. North-West. J. Zool. 2014, 10, 245–250. [Google Scholar]
- Pamilo, P.; Seppä, P.; Helanterä, H. Population Genetics of Wood Ants. In Wood Ant Ecology and Conservation; Stockan, J.A., Robinson, E.J.H., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 51–80. ISBN 978-1-107-26140-2. [Google Scholar]
- Seifert, B. The Supercolonial European Wood Ant Formica paralugubris Seifert, 1996 (Hymenoptera: Formicidae) Introduced to Canada and Its Predicted Role in Nearctic Forests. Myrmecol. News 2016, 22, 11–20. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frizzi, F.; Masoni, A.; Santedicola, M.; Servini, M.; Simoncini, N.; Palmieri, J.; Santini, G. Intraspecific Relationships and Nest Mound Shape Are Affected by Habitat Features in Introduced Populations of the Red Wood Ant Formica paralugubris. Insects 2022, 13, 198. https://doi.org/10.3390/insects13020198
Frizzi F, Masoni A, Santedicola M, Servini M, Simoncini N, Palmieri J, Santini G. Intraspecific Relationships and Nest Mound Shape Are Affected by Habitat Features in Introduced Populations of the Red Wood Ant Formica paralugubris. Insects. 2022; 13(2):198. https://doi.org/10.3390/insects13020198
Chicago/Turabian StyleFrizzi, Filippo, Alberto Masoni, Margherita Santedicola, Martina Servini, Nicola Simoncini, Jessica Palmieri, and Giacomo Santini. 2022. "Intraspecific Relationships and Nest Mound Shape Are Affected by Habitat Features in Introduced Populations of the Red Wood Ant Formica paralugubris" Insects 13, no. 2: 198. https://doi.org/10.3390/insects13020198
APA StyleFrizzi, F., Masoni, A., Santedicola, M., Servini, M., Simoncini, N., Palmieri, J., & Santini, G. (2022). Intraspecific Relationships and Nest Mound Shape Are Affected by Habitat Features in Introduced Populations of the Red Wood Ant Formica paralugubris. Insects, 13(2), 198. https://doi.org/10.3390/insects13020198