
Inter-Alpha-Trypsin Inhibitor Heavy Chain 4 Plays an Important Role in the Development and Reproduction of Nilaparvata lugens

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Cloning the cDNA of NlITIH4
2.3. Sequence Comparison and Bioinformatic Analysis
2.4. Double-Strand RNA Synthesis and Injection
2.5. Real-Time Quantitative PCR Analysis
2.6. Survival Rate Statistic, Dissection Observation and Fertility Analysis
2.7. Data Analysis
3. Results
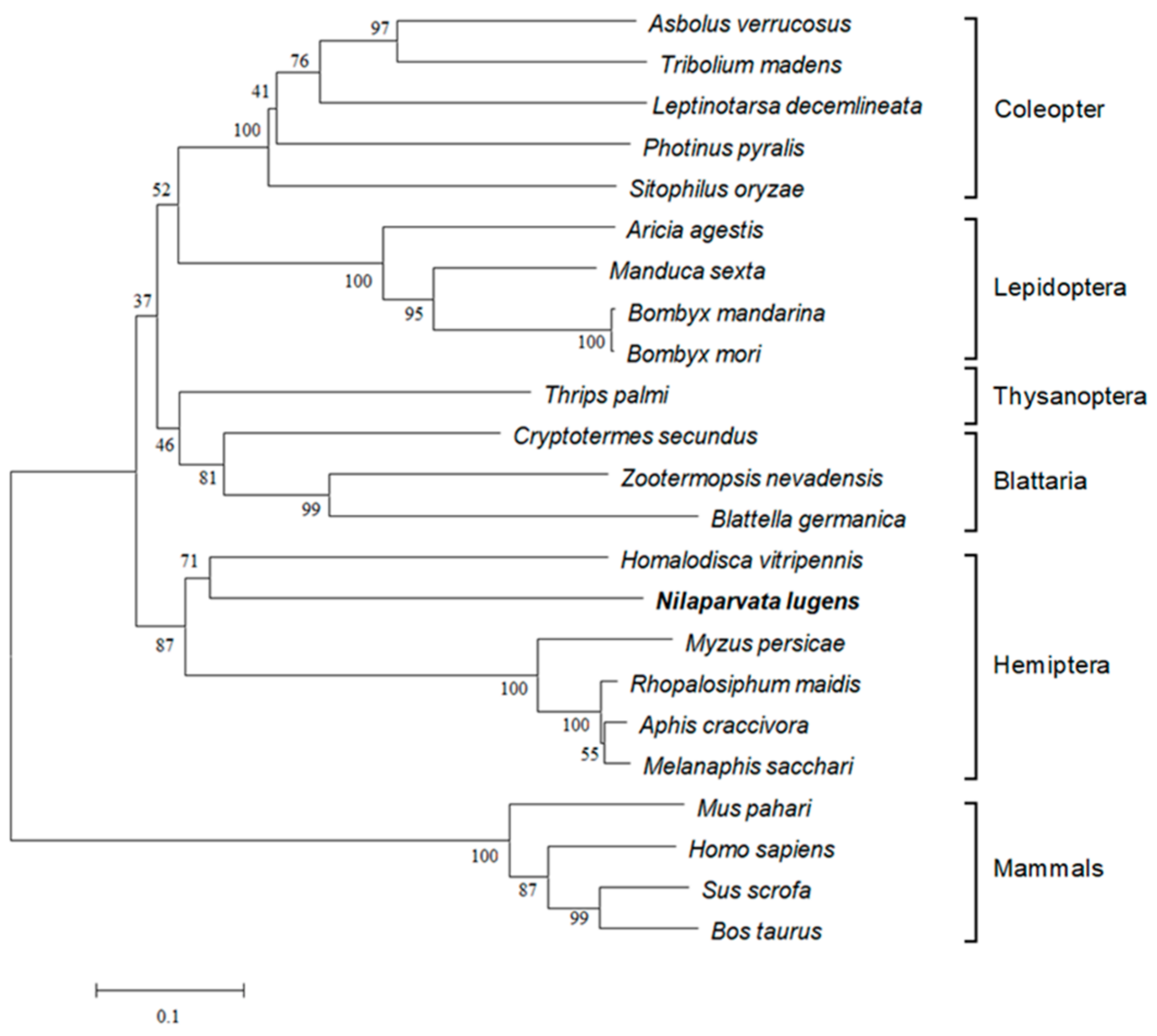
3.1. Identification and Bioinformatic Analysis of NlITIH4
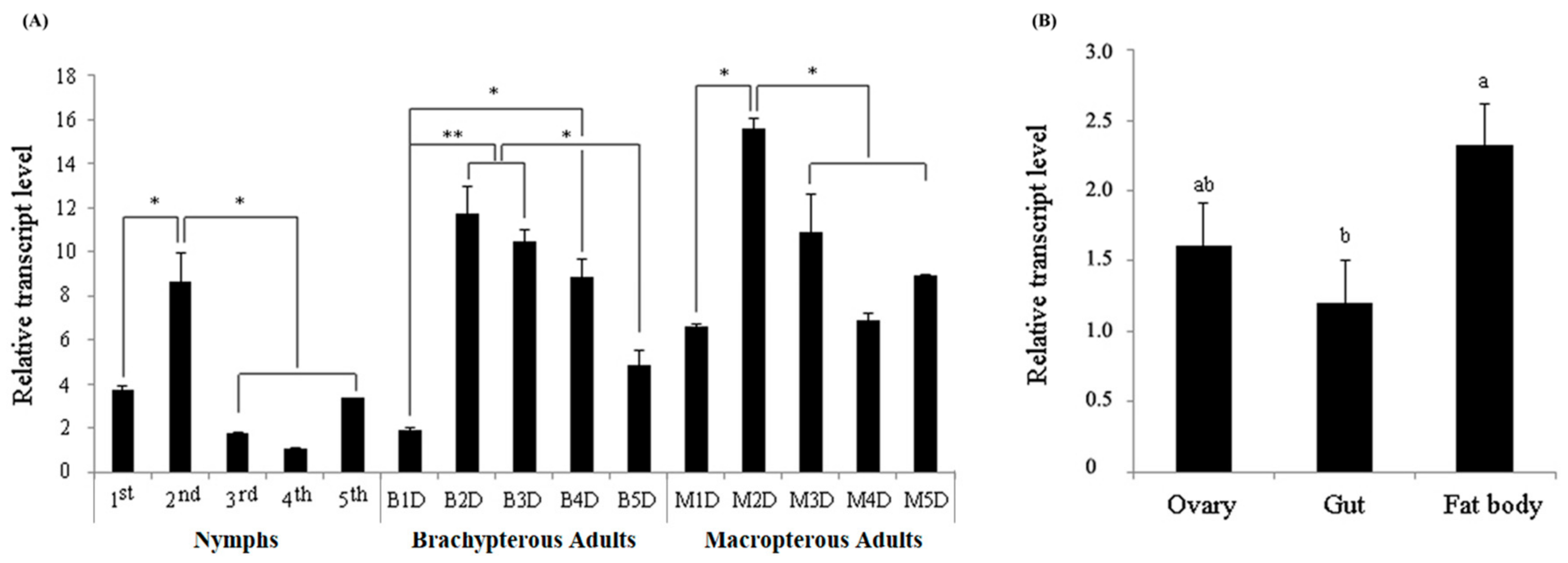
3.2. Developmental and Tissue-Specific Expression of NlITIH4
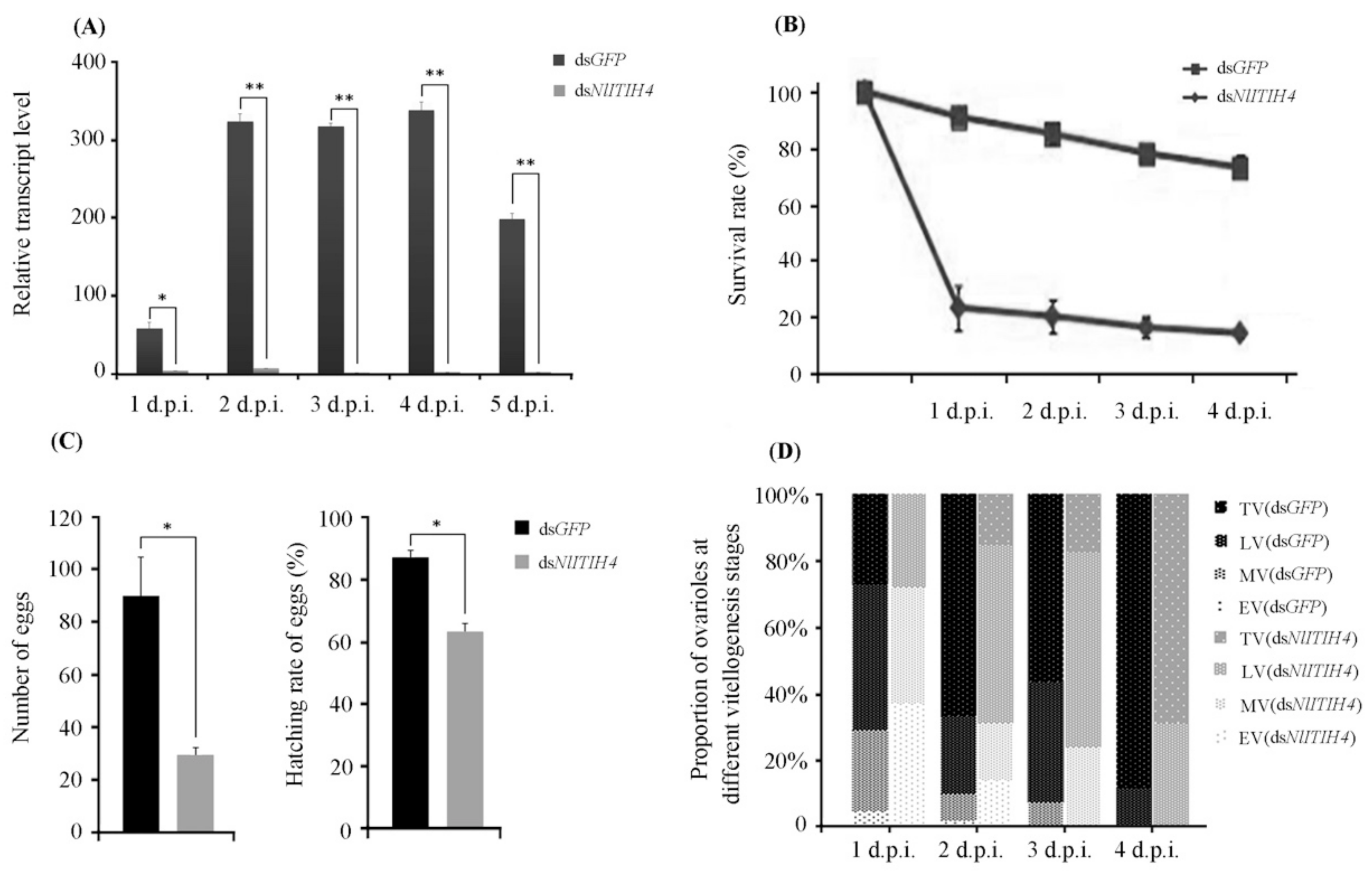
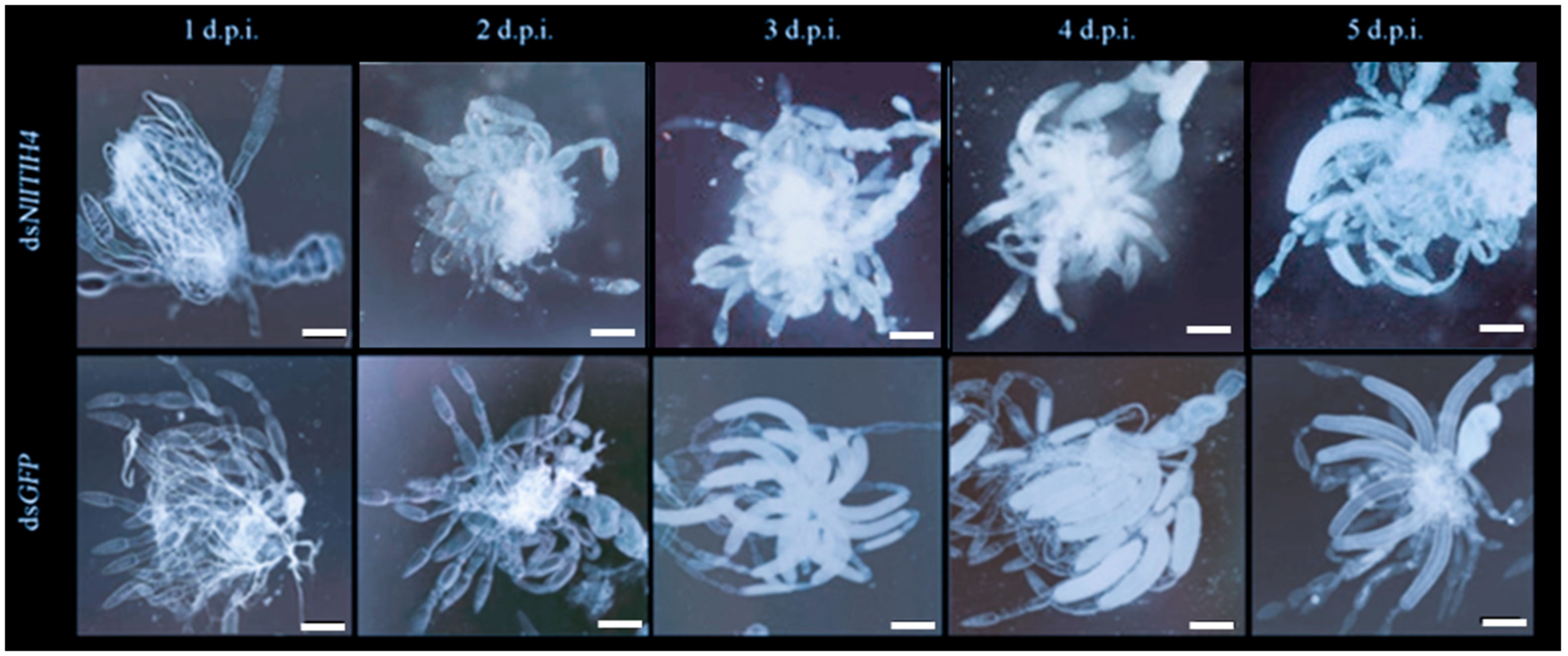
3.3. RNAi Effect of dsNlITIH4 Injection
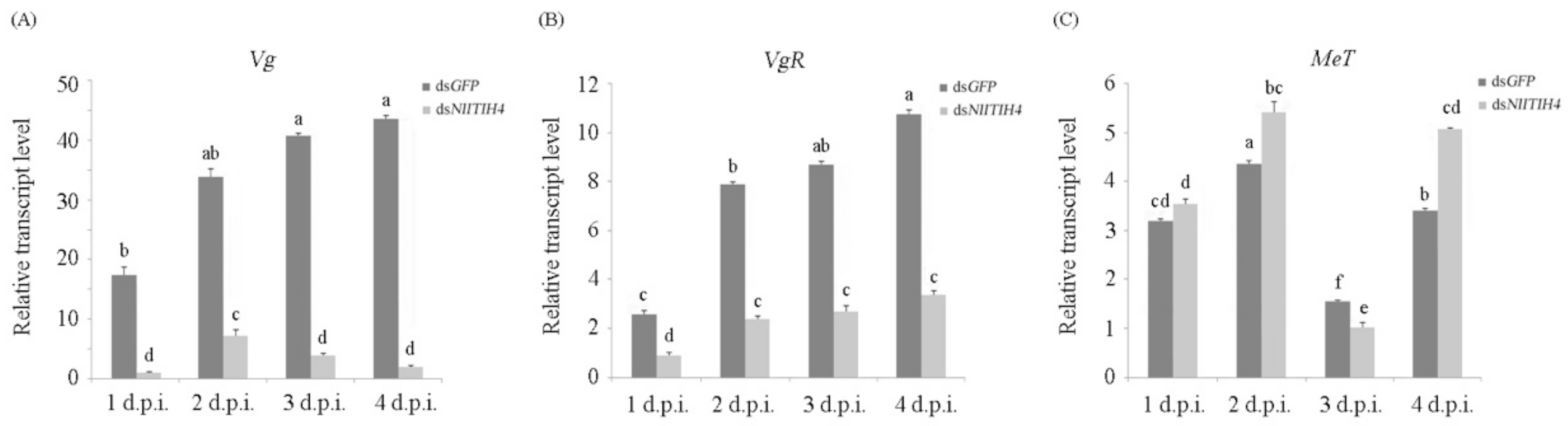
3.4. Effect of RNAi on the Expression of Vg, VgR and MeT
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bao, Y.; Zhang, C. Recent advances in molecular biology research of a rice pest, the brown planthopper. J. Integr. Agric. 2019, 18, 716–728. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Xiao, C.; He, Y. Recent progress on the genetics and molecular breeding of brown planthopper resistance in rice. Rice 2016, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebhard, W.; Hochstrasser, K.; Fritz, H.; Enghild, J.J.; Pizzo, S.V.; Salvesen, G. Structure of inter-alpha-inhibitor (inter-alpha-trypsin inhibitor) and pre-alpha-inhibitor: Current state and proposition of a new terminology. Biol. Chem. Hoppe-Seyler 1990, 371, 13–22. [Google Scholar] [PubMed]
- Salier, J.P. Inter-alpha-trypsin inhibitor: Emergence of a family within the Kunitz-type protease inhibitor superfamily. Trends Biochem. Sci. 1990, 15, 435–439. [Google Scholar] [CrossRef]
- Salier, J.P.; Rouet, P.; Raguenez, G.; Daveau, M. The inter-alpha-inhibitor family: From structure to regulation. Biochem. J. 1996, 315, 1–9. [Google Scholar] [CrossRef]
- Zhuo, L.; Kimata, K. Structure and function of inter-alpha-trypsin inhibitor heavy chains. Connect. Tissue Res. 2008, 49, 311–320. [Google Scholar] [CrossRef]
- Enghild, J.J.; Salvesen, G.; Thøgersen, I.B.; Valnickova, Z.; Pizzo, S.V.; Hefta, S.A. Presence of the protein-glycosaminoglycan-protein covalent cross-link in the inter-alpha-inhibitor-related proteinase inhibitor heavy chain 2/bikunin. J. Biol. Chem. 1993, 268, 8711–8716. [Google Scholar] [CrossRef]
- Zhuo, L.; Hascall, V.C.; Kimata, K. Inter-alpha-trypsin inhibitor, a covalent protein-glycosaminoglycan-protein complex. J. Biol. Chem. 2004, 279, 38079–38082. [Google Scholar] [CrossRef] [Green Version]
- Fries, E.; Blom, A.M. Bikunin—Not just a plasma proteinase inhibitor. Int. J. Biochem. Cell Biol. 2000, 32, 125–137. [Google Scholar] [CrossRef]
- Huang, L.; Yoneda, M.; Kimata, K. A serum-derived hyaluronan-associated protein (SHAP) is the heavy chain of the inter alpha-trypsin inhibitor. J. Biol. Chem. 1993, 268, 26725–26730. [Google Scholar] [CrossRef]
- Yoneda, M.; Suzuki, S.; Kimata, K. Hyaluronic acid associated with the surfaces of cultured fibroblasts is linked to a serum-derived 85-kDa protein. J. Biol. Chem. 1990, 265, 5247–5257. [Google Scholar] [CrossRef]
- Bost, F.; Diarra-Mehrpour, M.; Martin, J.P. Inter-alpha-trypsin inhibitor proteoglycan family—A group of proteins binding and stabilizing the extracellular matrix. Eur. J. Biochem. 1998, 252, 339–346. [Google Scholar] [CrossRef]
- Zhao, M.; Yoneda, M.; Ohashi, Y.; Kurono, S.; Iwata, H.; Ohnuki, Y.; Kimata, K. Evidence for the covalent binding of SHAP, heavy chains of inter-alpha-trypsin inhibitor, to hyaluronan. J. Biol. Chem. 1995, 270, 26657–26663. [Google Scholar] [CrossRef] [Green Version]
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its nature, distribution, functions and turnover. J. Intern. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef]
- Hutadilok, N.; Ghosh, P.; Brooks, P.M. Binding of haptoglobin, inter-alpha-trypsin inhibitor, and alpha 1 proteinase inhibitor to synovial fluid hyaluronate and the influence of these proteins on its degradation by oxygen derived free radicals. Ann. Rheum. Dis. 1988, 47, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Geisert, R.D.; Yelich, J.V.; Pratt, T.; Pomp, D. Expression of an inter-alpha-trypsin inhibitor heavy chain-like protein in the pig endometrium during the oestrous cycle and early pregnancy. J. Reprod. Fertil. 1998, 114, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, L.; Yoneda, M.; Zhao, M.; Yingsung, W.; Yoshida, N.; Kitagawa, Y.; Kawamura, K.; Suzuki, T.; Kimata, K. Defect in SHAP-hyaluronan complex causes severe female infertility. A study by inactivation of the bikunin gene in mice. J. Biol. Chem. 2001, 276, 7693–7696. [Google Scholar] [CrossRef] [Green Version]
- Nagyova, E. The Biological Role of Hyaluronan-Rich Oocyte-Cumulus Extracellular Matrix in Female Reproduction. Int. J. Mol. Sci. 2018, 19, 283. [Google Scholar] [CrossRef] [Green Version]
- Saguchi, K.; Tobe, T.; Hashimoto, K.; Sano, Y.; Nakano, Y.; Miura, N.H.; Tomita, M. Cloning and characterization of cDNA for inter-alpha-trypsin inhibitor family heavy chain-related protein (IHRP), a novel human plasma glycoprotein. J. Biochem. 1995, 117, 14–18. [Google Scholar] [CrossRef]
- Choi-Miura, N.H.; Takahashi, K.; Yoda, M.; Saito, K.; Hori, M.; Ozaki, H.; Mazda, T.; Tomita, M. The novel acute phase protein, IHRP, inhibits actin polymerization and phagocytosis of polymorphonuclear cells. Inflamm. Res. 2000, 49, 305–310. [Google Scholar] [CrossRef]
- Piñeiro, M.; Andrés, M.; Iturralde, M.; Carmona, S.; Hirvonen, J.; Pyörälä, S.; Heegaard, P.M.; Tjørnehøj, K.; Lampreave, F.; Piñeiro, A.; et al. ITIH4 (inter-alpha-trypsin inhibitor heavy chain 4) is a new acute-phase protein isolated from cattle during experimental infection. Infect. Immun. 2004, 72, 3777–3782. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Yarmush, D.; Berthiaume, F.; Jayaraman, A.; Yarmush, M.L. Immunodepletion of albumin for two-dimensional gel detection of new mouse acute-phase protein and other plasma proteins. Proteomics 2005, 5, 3991–4000. [Google Scholar] [CrossRef]
- Nayak, A.R.; Kashyap, R.S.; Kabra, D.; Purohit, H.J.; Taori, G.M.; Daginawala, H.F. Time course of inflammatory cytokines in acute ischemic stroke patients and their relation to inter-alfa trypsin inhibitor heavy chain 4 and outcome. Ann. Indian Acad. Neurol. 2012, 15, 181–185. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, W.; Zhao, B.; Li, L. Relationship between inter-α-trypsin inhibitor heavy chain 4 and ovarian cancer. Chin. J. Cancer Res. 2019, 31, 955–964. [Google Scholar] [CrossRef]
- Fung, E.T.; Yip, T.T.; Lomas, L.; Wang, Z.; Yip, C.; Meng, X.Y.; Lin, S.; Zhang, F.; Zhang, Z.; Chan, D.W.; et al. Classification of cancer types by measuring variants of host response proteins using SELDI serum assays. Int. J. Cancer 2005, 115, 783–789. [Google Scholar] [CrossRef]
- Esplin, M.S.; Merrell, K.; Goldenberg, R.; Lai, Y.; Iams, J.D.; Mercer, B.; Spong, C.Y.; Miodovnik, M.; Simhan, H.N.; van Dorsten, P.; et al. Proteomic identification of serum peptides predicting subsequent spontaneous preterm birth. Am. J. Obstet. Gynecol. 2011, 204, 391.e1–391.e8. [Google Scholar] [CrossRef] [Green Version]
- Soler, L.; Dąbrowski, R.; García, N.; Alava, M.A.; Lampreave, F.; Piñeiro, M.; Wawron, W.; Szczubiał, M.; Bochniarz, M. Acute-phase inter-alpha-trypsin inhibitor heavy chain 4 (ITIH4) levels in serum and milk of cows with subclinical mastitis caused by Streptococcus species and coagulase-negative Staphylococcus species. J. Dairy Sci. 2019, 102, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lu, K.; Shu, Y.; Zhou, J.; Zhang, X.; Zhang, X.; Chen, M.; Yao, Q.; Zhou, Q.; Zhang, W. Molecular characterization and RNA interference analysis of vitellogenin receptor from Nilaparvata lugens (Stål). J. Insect Physiol. 2015, 73, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Springer, T.A. Complement and the multifaceted functions of VWA and integrin I domains. Structure 2006, 14, 1611–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cray, C. Acute phase proteins in animals. Prog. Mol. Biol. Transl. Sci. 2012, 105, 113–150. [Google Scholar] [CrossRef]
- Tiu, S.H.; Benzie, J.; Chan, S.M. From hepatopancreas to ovary: Molecular characterization of a shrimp vitellogenin receptor involved in the processing of vitellogenin. Biol. Reprod. 2008, 79, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, M.L.; Bell, W.J.; Telfer, W.H. Vitellogenic blood protein synthesis by insect fat body. Science 1969, 165, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, Y.; Lou, Y.; Zhang, C. Vitellogenin and Vitellogenin-Like Genes in the Brown Planthopper. Front. Physiol. 2019, 10, 1181. [Google Scholar] [CrossRef]
- Yuan, L.J.; Peng, C.; Liu, B.H.; Feng, J.B.; Qiu, G.F. Identification and Characterization of a Luteinizing Hormone Receptor (LHR) Homolog from the Chinese Mitten Crab Eriocheir sinensis. Int. J. Mol. Sci. 2019, 20, 1736. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Cheng, Y.; Li, Y.; Li, W.; Song, Y.; Zeng, R.; Sun, Z. The KNRL nuclear receptor controls hydrolase-mediated vitellin breakdown during embryogenesis in the brown planthopper, Nilaparvata lugens. Insect Sci. 2021, 28, 1633–1650. [Google Scholar] [CrossRef]
- Bertuso, A.G.; Morooka, S.; Tojo, S. Sensitive periods for wing development and precocious metamorphosis after precocene treatment of the brown planthopper, Nilaparvata lugens. J. Insect Physiol. 2002, 48, 221–229. [Google Scholar] [CrossRef]
- Luo, W.; Liu, S.; Zhang, W.; Yang, L.; Huang, J.; Zhou, S.; Feng, Q.; Palli, S.R.; Wang, J.; Roth, S.; et al. Juvenile hormone signaling promotes ovulation and maintains egg shape by inducing expression of extracellular matrix genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2104461118. [Google Scholar] [CrossRef]
- Lin, X.; Yao, Y.; Wang, B. Methoprene-tolerant (Met) and Krüpple-homologue 1 (Kr-h1) are required for ovariole development and egg maturation in the brown plant hopper. Sci. Rep. 2015, 5, 18064. [Google Scholar] [CrossRef] [Green Version]
- Robker, R.L.; Hennebold, J.D.; Russell, D.L. Coordination of Ovulation and Oocyte Maturation: A Good Egg at the Right Time. Endocrinology 2018, 159, 3209–3218. [Google Scholar] [CrossRef]
- Vigone, G.; Shuhaibar, L.C.; Egbert, J.R.; Uliasz, T.F.; Movsesian, M.A.; Jaffe, L.A. Multiple cAMP Phosphodiesterases Act Together to Prevent Premature Oocyte Meiosis and Ovulation. Endocrinology 2018, 159, 2142–2152. [Google Scholar] [CrossRef] [Green Version]
- Gérard, N.; Robin, E. Cellular and molecular mechanisms of the preovulatory follicle differenciation and ovulation: What do we know in the mare relative to other species. Theriogenology 2019, 130, 163–176. [Google Scholar] [CrossRef]
- Vanderhyden, B.C.; Armstrong, D.T. Role of cumulus cells and serum on the in vitro maturation, fertilization, and subsequent development of rat oocytes. Biol. Reprod. 1989, 40, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, L.; Kimata, K. Cumulus oophorus extracellular matrix: Its construction and regulation. Cell Struct. Funct. 2001, 26, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Fülöp, C.; Szántó, S.; Mukhopadhyay, D.; Bárdos, T.; Kamath, R.V.; Rugg, M.S.; Day, A.J.; Salustri, A.; Hascall, V.C.; Glant, T.T.; et al. Impaired cumulus mucification and female sterility in tumor necrosis factor-induced protein-6 deficient mice. Development 2003, 130, 2253–2261. [Google Scholar] [CrossRef] [Green Version]
- Obayashi, Y.; Yabushita, H.; Kanyama, K.; Noguchi, M.; Zhuo, L.; Kimata, K.; Wakatsuki, A. Role of serum-derived hyaluronan-associated protein-hyaluronan complex in ovarian cancer. Oncol. Rep. 2008, 19, 1245–1251. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequence (5′–3′) | Product Length |
|---|---|---|
| For cDNA cloning: | ||
| NlITIH4-F | TTTTGCTAACATTTTCCTCCTTG | 2798 bp |
| NlITIH4-R | AGTTTCTACGGCTTACTCATCAC | |
| For qPCR: | ||
| NlITIH4-qF | AAGAAAAGGAAGAAGCAAAAGC | 191 bp |
| NlITIH4-qR | ATGAGTGTAATGACCGAGGGA | |
| Nl18S-qF | GTAACCCGCTGAACCTCC | 170 bp |
| Nl18S-qR | GTCCGAAGACCTCACTAAATCA | |
| Vg-qF | TTCCGTTTGCAGCCACCTATG | 154 bp |
| Vg-qR | CTGCTGCTGCTGCTTCTGTCA | |
| VgR-qF | AGGCAGCCACACAGATAACCGC | 136 bp |
| VgR-qR | AGCCGCTCGCTCCAGAACATT | |
| Met-qF | GGTGGTAAACGGATTGGAAA | 100 bp |
| Met-qR | CATCGTCAGCCAACTCGATA | |
| For dsRNA synthesis: | ||
| NlITIH4-dsF | GGATCCTAATACGACTCACTATAGGACATCAGTGGTTCCATGT | 488 bp |
| NlITIH4-dsR | GGATCCTAATACGACTCACTATAGGGATCTGTCAAGTCGACA | |
| GFP-dsF | GGATCCTAATACGACTCACTATAGGGATACGTGCAGGAGAGGAC | 350 bp |
| GFP-dsR | GGATCCTAATACGACTCACTATAGGGCAGATTGTGTGGACAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, J.-L.; Han, S.-J.; Zhang, R.-J.; Yu, J.-B.; Li, Y.-B.; Yu, X.-P.; Liu, G.-F.; Xu, Y.-P. Inter-Alpha-Trypsin Inhibitor Heavy Chain 4 Plays an Important Role in the Development and Reproduction of Nilaparvata lugens. Insects 2022, 13, 303. https://doi.org/10.3390/insects13030303
Ji J-L, Han S-J, Zhang R-J, Yu J-B, Li Y-B, Yu X-P, Liu G-F, Xu Y-P. Inter-Alpha-Trypsin Inhibitor Heavy Chain 4 Plays an Important Role in the Development and Reproduction of Nilaparvata lugens. Insects. 2022; 13(3):303. https://doi.org/10.3390/insects13030303
Chicago/Turabian StyleJi, Jin-Liang, Shan-Jie Han, Rui-Juan Zhang, Jian-Bin Yu, Ya-Bin Li, Xiao-Ping Yu, Guang-Fu Liu, and Yi-Peng Xu. 2022. "Inter-Alpha-Trypsin Inhibitor Heavy Chain 4 Plays an Important Role in the Development and Reproduction of Nilaparvata lugens" Insects 13, no. 3: 303. https://doi.org/10.3390/insects13030303
APA StyleJi, J. -L., Han, S. -J., Zhang, R. -J., Yu, J. -B., Li, Y. -B., Yu, X. -P., Liu, G. -F., & Xu, Y. -P. (2022). Inter-Alpha-Trypsin Inhibitor Heavy Chain 4 Plays an Important Role in the Development and Reproduction of Nilaparvata lugens. Insects, 13(3), 303. https://doi.org/10.3390/insects13030303