
Demography and Fitness of Anastatus japonicus Reared from Antheraea pernyi as a Biological Control Agent of Caligula japonica



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Host Insects
2.2. Parasitoid
2.3. Performance of A. japonicus Reared from COS Eggs on COS and JGS Eggs
2.4. Data Analysis
3. Results
3.1. Longevity and Lifetime Fecundity of A. japonicus Parasitizing COS and JGS Eggs
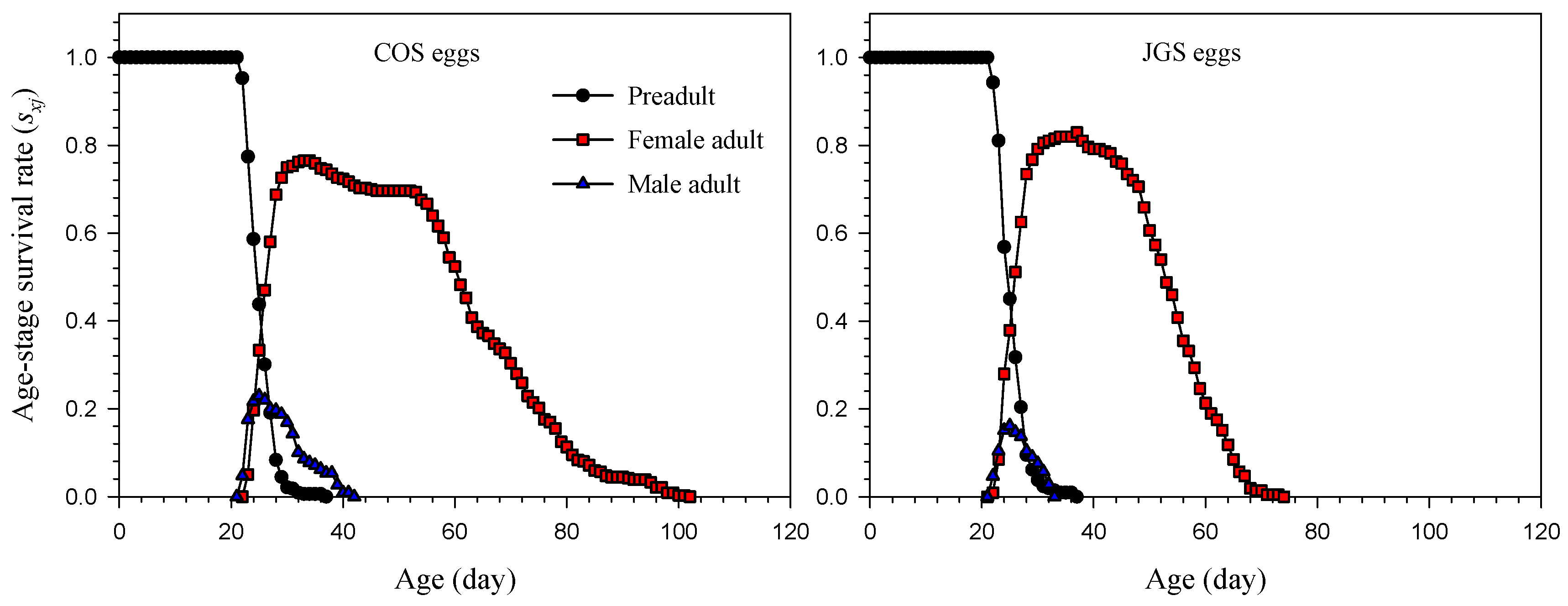
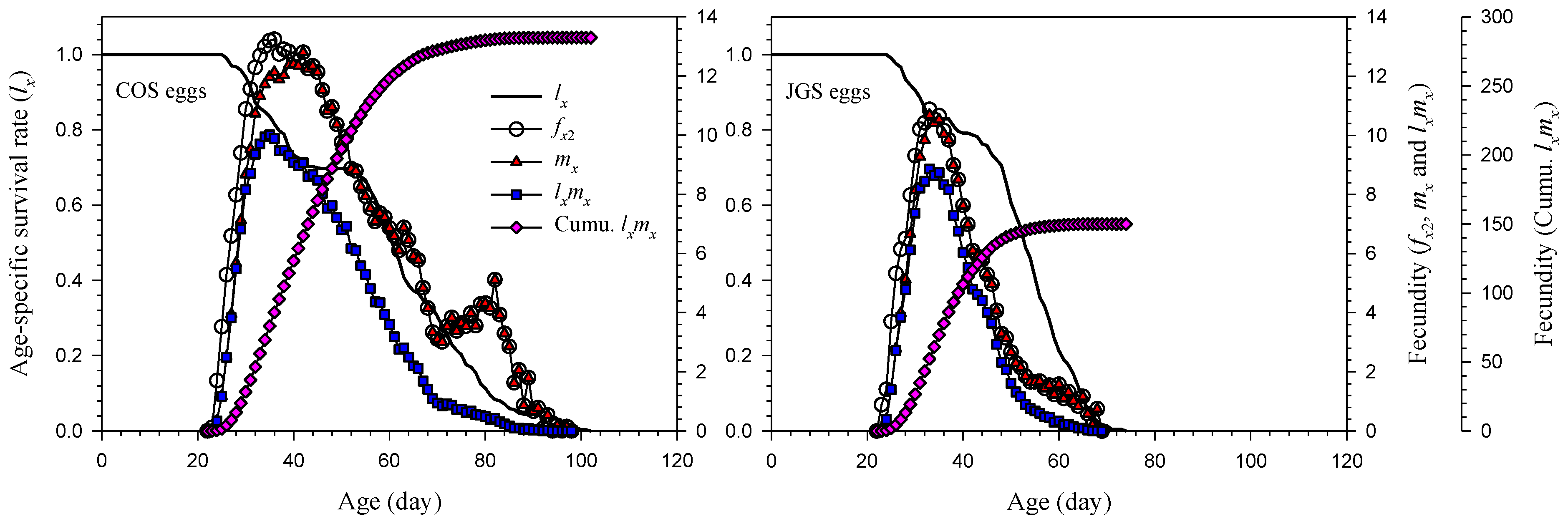
3.2. Age-Stage and Age-Specific Survival Rate and Fecundity of A. japonicus Parasitizing COS and JGS Eggs
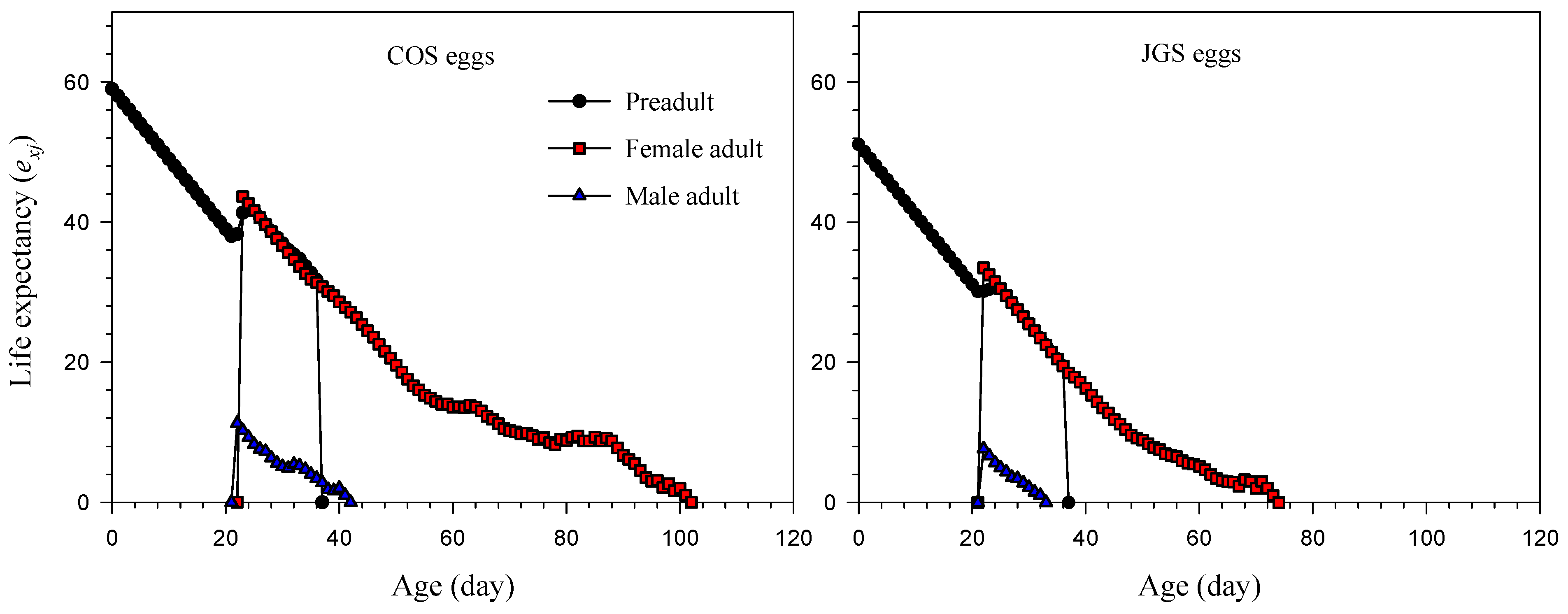
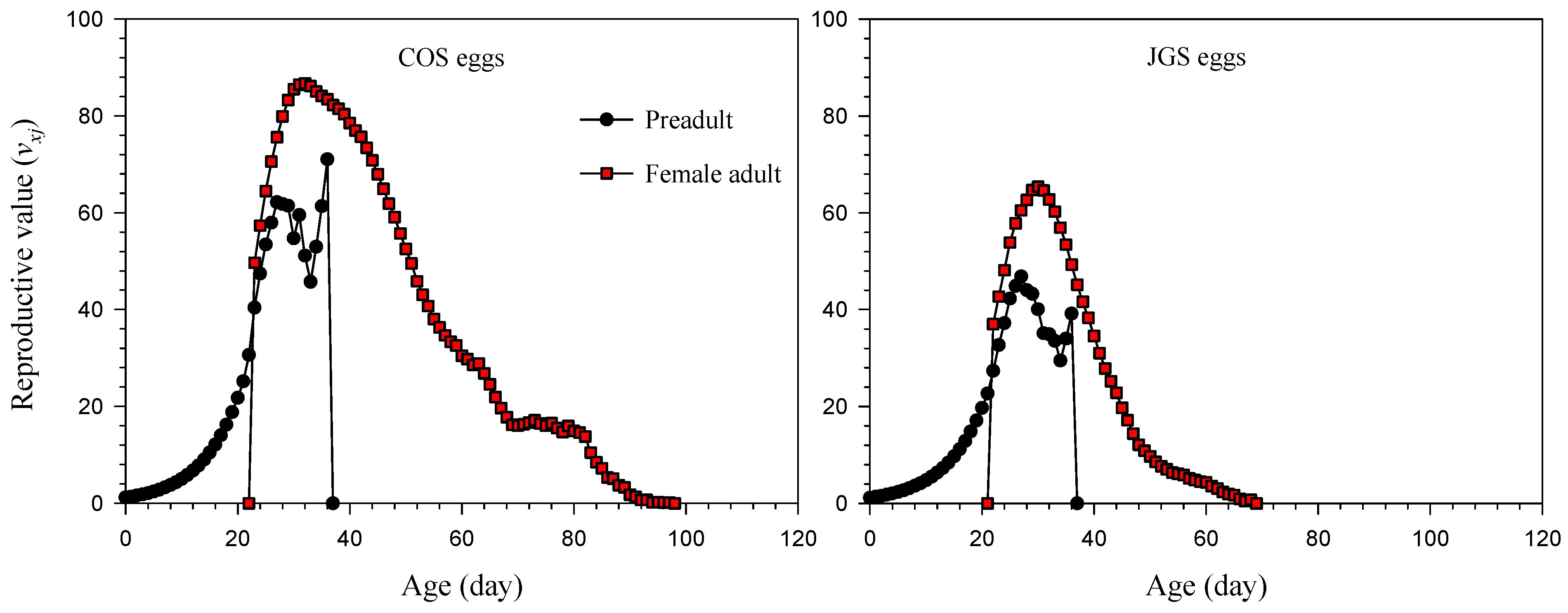
3.3. Life Expectancy and Reproductive Value of A. japonicus Parasitizing COS and JGS Eggs
3.4. Population Parameters of A. japonicus Parasitizing COS and JGS Eggs
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.M.; Gibson, G.A.P.; Peng, L.F.; Iqbal, A.; Zang, L.S. Anastatus Motschulsky (Hymenoptera, Eupelmidae): Egg parasitoids of Caligula japonica Moore (Lepidoptera, Saturniidae) in China. ZooKeys 2019, 881, 109–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.H.; Ge, W.G.; Wang, Y.F. Occurrence regularity and control measures of Ginkgo biloba. Jilin For. Sci. Technol. 1997, 6, 19–20. [Google Scholar]
- Huang, Y.Z. Prevention and control of walnut tree by Ginkgo biloba. Gansu Sci. Technol. 2014, 30, 161–162. [Google Scholar]
- Jiang, D.A. Study on the occurrence and control of Dictypoloca japonica. For. Sci. Technol. 2003, 1, 25–27. [Google Scholar]
- Qiao, X.; Wang, Y.C.; Wu, G.; Wang, S.C.; Hu, J.Z.; Liu, T.X.; Feng, S.Q. Occurrence reasons and control measures of Caligula Japonica in Longnan City of China. Plant Dis. Pests 2014, 5, 38–41. [Google Scholar] [CrossRef]
- Wu, N.M.; Yang, S.Z.; Chen, J.; Du, Y.B.; Cai, J.X.; Fu, C.S. A study on the Anastatus spp. released in forest for control of Dictyoploca japonica Moore. Nat. Enemies Insects 2001, 23, 26–29. [Google Scholar]
- Dong, D.J. Toxicity determination and control effect of several pesticides against Dictyoploca japonica Butler. For. Pest Dis. 2013, 32, 38–40. [Google Scholar]
- Wang, Y.X.; Qiao, X.; Wang, Z.K. Pharmacodynamic study on different insecticide for Caligula japonica Moore. Mod. Agric. Sci. Technol. 2012, 14, 95–96. [Google Scholar]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Huang, N.X.; Jaworski, C.C.; Desneux, N.; Zhang, F.; Yang, P.Y.; Wang, S. Long-term, large-scale releases of Trichogramma promotes pesticide decrease in maize in Jilin province, Northern China. Entomol. Gen. 2020, 40, 331–335. [Google Scholar] [CrossRef]
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Zapponi, L.; Tortorici, F.; Anfora, G.; Bardella, S.; Bariselli, M.; Benvenuto, L.; Bernardinelli, I.; Butturini, A.; Caruso, S.; Colla, R.; et al. Assessing the distribution of exotic egg parasitoids of Halyomorpha halys in Europe with a large-scale monitoring program. Insects 2021, 12, 316. [Google Scholar] [CrossRef] [PubMed]
- Stahl, J.M.; Babendreier, D.; Marazzi, C.; Caruso, S.; Costi, E.; Maistrello, L.; Haye, T. Can Anastatus bifasciatus be used for augmentative biological control of the brown marmorated stink bug in fruit orchards? Insects 2019, 10, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, J.M.; Babendreier, D.; Haye, T. Using the egg parasitoid Anastatus bifasciatus against the invasive brown marmorated stink bug in Europe: Can non-target effects be ruled out? J. Pest Sci. 2018, 91, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef]
- Lalitha, Y.; Ballal, C.R.; Gupta, A. Interaction between Anastatus acherontiae and Anastatus bangalorensis (Hymenoptera: Chalcidoidea) two potential parasitoids of Litchi stink bug Tessaratoma javanica Thunberg. In Conference on National Priorities in Plant Health; SV Agricultural College: Tirupati, India, 2016; pp. 4–5. [Google Scholar]
- Choudhary, J.S.; Prabhakar, C.S.; Moanaro; Das, B.; Kumar, S. Litchi stink bug (Tessaratoma javanica) outbreak in Jharkhand, India, on litchi. Phytoparasitica 2013, 41, 73–77. [Google Scholar] [CrossRef]
- Li, D.S.; Liao, C.; Zhang, B.X.; Song, Z.W. Biological control of insect pests in litchi orchards in China. Biol. Control 2014, 68, 23–36. [Google Scholar] [CrossRef]
- Han, S.C.; Liu, W.H.; Chen, Q.X.; Zeng, B.K.; Chen, N.R.; Lin, J.X.; Yu, F.X. Mass releasing Anastatus japonicus to control Tessaratoma papillosa in Hong Kong. J. Biol. Control 1999, 15, 54–56. [Google Scholar]
- Chen, Y.M.; Qu, X.R.; Li, T.H.; Iqbal, A.; Wang, X.G.; Ren, Z.Y.; Desneux, N.; Zang, L.S. Performances of six eupelmid egg parasitoids from China on Japanese giant silkworm Caligula japonica with different host age regimes. J. Pest Sci. 2020, 94, 309–319. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhang, X.; Zang, L.S.; Du, W.M.; Hou, Y.Y.; Ruan, C.C.; Desneux, N. Advantages of diapause in Trichogramma dendrolimi mass production via eggs of the Chinese silkworm, Antheraea pernyi. Pest Manag. Sci. 2018, 74, 959–965. [Google Scholar] [CrossRef]
- Li, T.H.; Tian, C.Y.; Zang, L.S.; Hou, Y.Y.; Ruan, C.C.; Yang, X.B.; Monticelli, L.; Desneux, N. Multiparasitism with Trichogramma dendrolimi on egg of Chinese oak silkworm, Antheraea pernyi, enhances emergence of Trichogramma ostriniae. J. Pest Sci. 2019, 92, 707–713. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Mitra, S.; Mobarak, S.H.; Barik, A. Age-stage, two-sex life table of the biocontrol agent, Altica cyanea on three Ludwigia species. Biologia 2021, 76, 101–112. [Google Scholar] [CrossRef]
- del Pino, M.; Gallego, J.R.; Suárez, E.H.; Cabello, T. Effect of temperature on life history and parasitization behavior of Trichogramma achaeae Nagaraja and Nagarkatti (Hym.: Trichogrammatidae). Insects 2020, 11, 482. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, C.L.; Yang, X.B.; Chi, H.; Dai, P.; Desneux, N.; Benelli, G.; Zang, L.S. Yacon as an alternative host plant for Encarsia formosa mass-rearing: Validating a multinomial theorem for bootstrap technique in life table research. Pest Manag. Sci. 2021, 77, 2324–2336. [Google Scholar] [CrossRef]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2020, 28, 602–610. [Google Scholar] [CrossRef]
- Ali, S.; Li, S.; Jaleel, W.; Khan, M.M.; Wang, J.T.; Zhou, X.M. Using a two-sex life table tool to calculate the fitness of Orius strigicollis as a predator of Pectinophora gossypiella. Insects 2020, 11, 275. [Google Scholar] [CrossRef]
- Farooq, M.; Shakeel, M.; Iftikhar, A.; Shahid, M.R.; Zhu, X. Age-stage, two-sex life tables of the lady beetle (Coleoptera: Coccinellidae) feeding on different aphid species. J. Econ. Entomol. 2018, 111, 575–585. [Google Scholar] [CrossRef]
- Bussaman, P.; Sa-uth, C.; Chandrapatya, A.; Atlihan, R.; Gokce, A.; Saska, P.; Chi, H. Fast population growth in physogastry reproduction of Luciaphorus perniciosus rack (Acari: Pygmephordae) at different temperatures. J. Econ. Entomol. 2017, 110, 1397–1403. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chi, H.; Liu, T.X. Demography and parasitic effectiveness of Aphelinus asychis reared from Sitobion avenae as a biological control agent of Myzus persicae reared on chili pepper and cabbage. Biol. Control 2016, 92, 111–119. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Chi, H. Demographic comparison of sweet potato weevil reared on a major host, Ipomoea batatas, and an alternative host, I. triloba. Sci. Rep. 2015, 5, 11871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2021. Available online: http://140.120.197.173/Ecology (accessed on 9 February 2022).
- Chi, H. CONSUME-MSChart: A Computer Program for Consumption Rate Analysis Based on the Age Stage, Two-Sex Life Table. 2021. Available online: http://140.120.197.173/ecology (accessed on 9 February 2022).
- Chi, H.; You, M.S.; Atlıhan, R.; Smith, C.L.; Kavousi, A.; Özgökçe, M.S.; Güncan, A.; Tuan, S.J.; Fu, J.W.; Xu, Y.Y.; et al. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2019, 40, 103–124. [Google Scholar] [CrossRef]
- Chen, Y.M.; Iqbal, A.; Lv, R.E.; Wang, X.G.; Desneux, N.; Zang, L.S. Chinese oak silkworm Antherae pernyi egg, a suitable factitious host for rearing eupelmid egg parasitoids. Pest Manag. Sci. 2022, 78. [Google Scholar] [CrossRef]
- Chen, Y.M.; Sun, J.W.; Iqbal, A.; Lu, R.; Wang, H.; Zang, L.S. An investigation of Caligula japonica (Lepidoptera: Saturniidae) egg distribution and associated parasitoids on walnut trees (Juglans regia L.) in northwestern China. Int. J. Pest Manag. 2020, 1–8. [Google Scholar] [CrossRef]
- Gibson, G.A.P. Revision of the Old world genus Mesocomys Cameron (Hymenoptera: Eupelmidae). Zootaxa 2021, 4901, 1–92. [Google Scholar] [CrossRef]
- Amir-Maafi, M.; Chi, H.; Chen, Z.Z.; Xu, Y.Y. Innovative bootstrap-match technique for life table set up. Entomol. Gen. 2022, 42. [Google Scholar] [CrossRef]
- Mou, D.F.; Lee, C.C.; Smith, C.L.; Chi, H. Using viable eggs to accurately determine the demographic and predation potential of Harmonia dimidiata (Coleoptera: Coccinellidae). J. Appl. Entomol. 2015, 139, 579–591. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 1936. [Google Scholar] [CrossRef]
- Wei, M.F.; Chi, H.; Guo, Y.F.; Li, X.W.; Zhao, L.L.; Ma, R.Y. Demography of Cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of Pyrus bretschneideri and P. communis (Rosales: Rosaceae) pears with estimations of confidence intervals of specific life table statistics. J. Econ. Entomol. 2020, 113, 2343–2353. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Y.M.; Yang, X.B.; Lv, R.E.; Desneux, N.; Zang, L.S. Parasitism and suitability of Aprostocetus brevipedicellus on Chinese oak silkworm, Antheraea pernyi, a dominant factitious host. Insects 2021, 12, 694. [Google Scholar] [CrossRef]
- Mochiah, M.B.; Ngi-Song, A.J.; Overholt, W.A.; Botchey, M. Host suitability of four cereal stem borers (Lepidoptera: Crambidae, Noctuidae) for different geographic populations of Cotesia sesamiae (Cameron) (Hymenoptera: Braconidae) in Kenya. Biol. Control 2001, 21, 285–292. [Google Scholar] [CrossRef]
- Le Masurier, A.D. Effect of host size on clutch size in Cotesia glomerata. J. Anim. Ecol. 1991, 60, 107–118. [Google Scholar] [CrossRef]
- Jiang, X.J.; Zhou, Z.J. Effects of substitute host pupal stages on feeding and oviposition of Scleroderma sichuanensis (Hymenoptera: Bethylidae). Guangxi For. Sci. 2005, 34, 179–180. [Google Scholar]
- Wang, X.Y.; Yang, Z.Q. Host adaptations of the generalist parasitoids and some factors influencing the choice of hosts. Acta Ecol. Sin. 2010, 30, 1615–1627. [Google Scholar]
- Vinson, S.B. The general host selection behavior of parasitoid Hymenoptera and a comparison of initial strategies utilized by larvaphagous and oophagous species. Biol. Control 1998, 11, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M. The influence of the thickness of host egg chorion on the longevity and fecundity of Trichogramma confusum. Nat. Enemies Insects 1999, 21, 150–151. [Google Scholar]
- Pennacchio, F.; Digilio, M.C.; Tremblay, E.; Tranfaglia, A. Host recognition and acceptance behaviour in two aphid parasitoid species: Aphidius ervi and Aphidius microlophii (Hymenoptera: Braconidae). Bull. Entomol. Res. 1994, 84, 57–64. [Google Scholar] [CrossRef]
- Shin, S.S.; Chun, Y.S.; Ryoo, M.I. Functional and numerical responses of Anisopteromalus calandrae and Lariophagus distinguendus (Hymenoptera: Pteromalidae) to the various densities of an alternative host, Callosobruchus chinensis. Korean J. Entomol. 1994, 24, 199–206. [Google Scholar]
- Xu, W.; Wen, X.Y.; Hou, Y.Y.; Desneux, N.; Ali, A.; Zang, L.S. Suitability of Chinese oak silkworm eggs for the multigenerational rearing of the parasitoid Trichogramma leucaniae. PLoS ONE 2020, 15, e0231098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giunti, G.; Canale, A.; Messing, R.H.; Donati, E.; Stefanini, C.; Michaud, J.P.; Benelli, G. Parasitoid learning: Current knowledge and implications for biological control. Biol. Control 2015, 90, 208–219. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.B.; Lin, Y.; Zang, L.S.; Tian, C.Y.; Ruan, C.C. Effect of fertilized, unfertilized, and UV-irradiated hosts on parasitism and suitability for Trichogramma parasitoids. Entomol. Exp. Appl. 2016, 161, 50–56. [Google Scholar] [CrossRef]
- Lu, A.P. Sexual differentiation of parasitic Hymenoptera through nutrition 3. The changes of sexual ratio after the transplantation of egg. Suppl. J. Sun YatSen Univ. 1995, 3, 80–83. [Google Scholar]
- Yang, X.; Qu, Y.L.; Wu, Z.Y.; Lin, Y.; Ruan, C.C.; Desneux, N.; Zang, L.S. Parasitism and suitability of fertilized and nonfertilized eggs of the rice striped stem borer, Chilo suppressalis (Lepidoptera: Crambidae), for Trichogramma parasitoids. J. Econ. Entomol. 2016, 109, 1524–1528. [Google Scholar] [CrossRef]
- Krugner, R. Suitability of non-fertilized eggs of Homalodisca vitripennis for the egg parasitoid Gonatocerus morrilli. BioControl 2014, 59, 167–174. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | COS Eggs 1 | JGS Eggs 1 |
|---|---|---|
| Initial cohort size (female: male) | 20:20 | 24:24 |
| Matched cohort size (female: male) | 259:77 | 175:36 |
| Female adult longevity (days) | 40.50 ± 0.84 a | 29.43 ± 0.54 b |
| Female total longevity (days) | 66.64 ± 0.87 a | 55.49 ± 0.57 b |
| Male adult longevity (days) | 10.19 ± 0.50 a | 6.47 ± 0.37 b |
| Male total longevity (days) | 33.27 ± 0.51 a | 29.69 ± 0.42 b |
| Adult preoviposition period (days) | 0.85 ± 0.03 a | 0.92 ± 0.03 a |
| Total preoviposition period (days) | 26.99 ± 0.14 a | 26.98 ± 0.19 a |
| Oviposition days | 35.88 ± 0.71 a | 24.02 ± 0.46 b |
| Eggs laid per oviposition day | 10.30 ± 0.05 a | 7.53 ± 0.06 b |
| Total fecundity (eggs/female) | 369.7 ± 7.0 a | 180.9 ± 3.8 b |
| Age at 0.5 lx (days) | 61.00 ± 0.85 a | 53.00 ± 0.89 b |
| Population Parameter | COS Eggs 1 | JGS Eggs 1 |
|---|---|---|
| Net reproduction rate (R0) (offspring) | 284.9 ± 10.0 a | 150.0 ± 5.6 b |
| R0, F (female offspring) | 205.3 ± 6.9 a | 118.7 ± 4.3 b |
| R0, M (male offspring) | 79.7 ± 3.4 a | 31.4 ± 1.8 b |
| Intrinsic rate of increase (r) (day−1) | 0.1466 ± 0.0011 a | 0.1419 ± 0.0012 b |
| Finite rate of increase (λ) (day−1) | 1.1579 ± 0.0012 a | 1.1525 ± 0.0014 b |
| Mean generation time (T) (days) | 38.55 ± 0.19 a | 35.31 ± 0.20 b |
| Net non-effective parasitism rate (G0) | 0.00 b | 9.60 ± 0.61 a |
| Net parasitism rate (P0) (eggs/parasitoid) | 284.9 ± 10.0 a | 159.6 ± 6.0 b |
| Transformation rate (Qp) | 1.00 ± 0.00 b | 1.0640 ± 0.0033 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.-Y.; Chen, Y.-M.; Wang, X.; Lv, R.-E.; Zang, L.-S. Demography and Fitness of Anastatus japonicus Reared from Antheraea pernyi as a Biological Control Agent of Caligula japonica. Insects 2022, 13, 349. https://doi.org/10.3390/insects13040349
Wei X-Y, Chen Y-M, Wang X, Lv R-E, Zang L-S. Demography and Fitness of Anastatus japonicus Reared from Antheraea pernyi as a Biological Control Agent of Caligula japonica. Insects. 2022; 13(4):349. https://doi.org/10.3390/insects13040349
Chicago/Turabian StyleWei, Xiao-Yan, Yong-Ming Chen, Xingeng Wang, Rui-E Lv, and Lian-Sheng Zang. 2022. "Demography and Fitness of Anastatus japonicus Reared from Antheraea pernyi as a Biological Control Agent of Caligula japonica" Insects 13, no. 4: 349. https://doi.org/10.3390/insects13040349
APA StyleWei, X. -Y., Chen, Y. -M., Wang, X., Lv, R. -E., & Zang, L. -S. (2022). Demography and Fitness of Anastatus japonicus Reared from Antheraea pernyi as a Biological Control Agent of Caligula japonica. Insects, 13(4), 349. https://doi.org/10.3390/insects13040349