
Surveys of Drosophila suzukii (Diptera: Drosophilidae) and Its Host Fruits and Associated Parasitoids in Northeastern China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
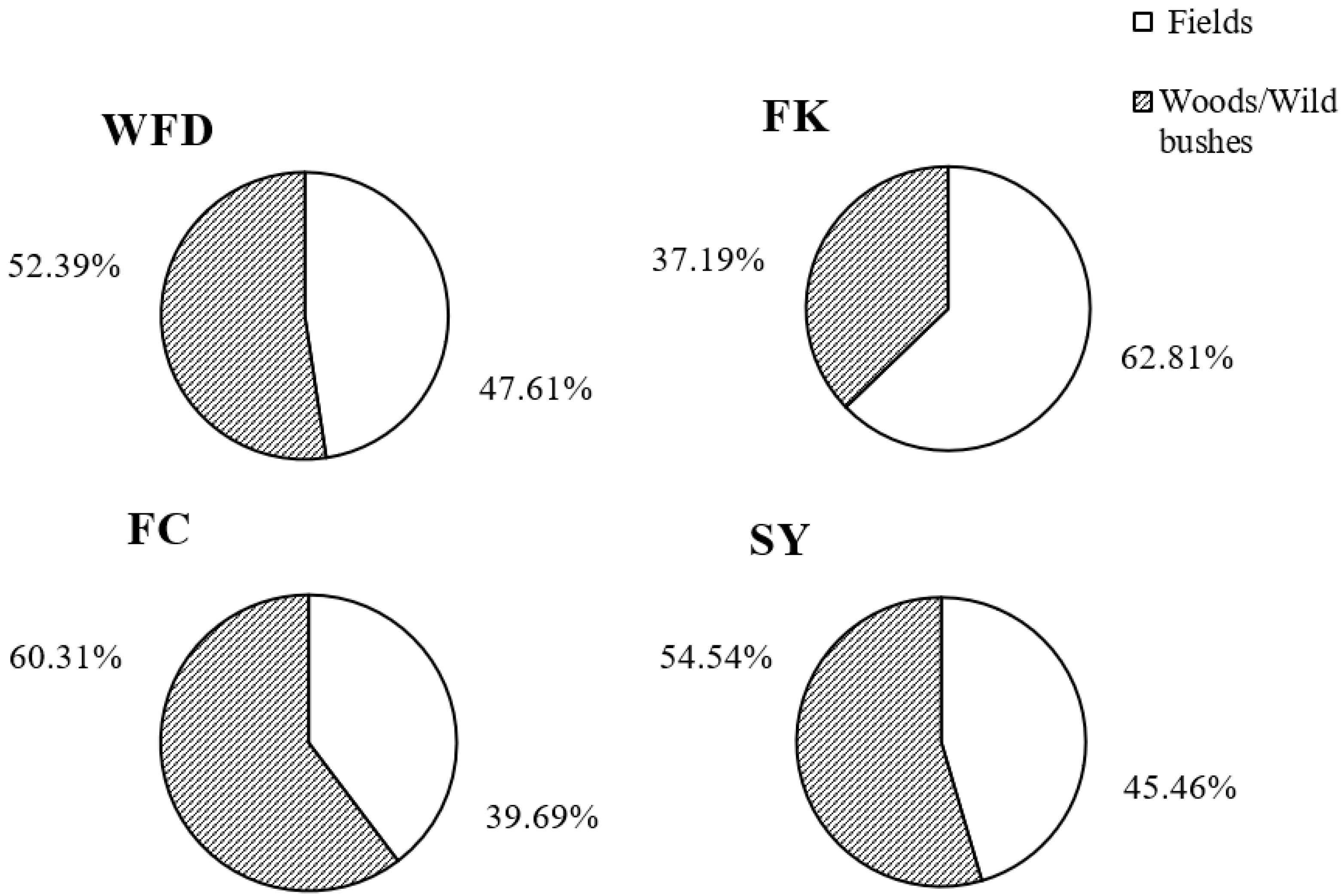
2.1. Survey Locations
2.2. Collections of D. suzukii and Parasitoids
2.3. Morphological Identification of D. suzukii and Parasitoids
2.4. Statistical Analysis
3. Results
3.1. Morphological Characteristics of D. suzukii and Parasitoids
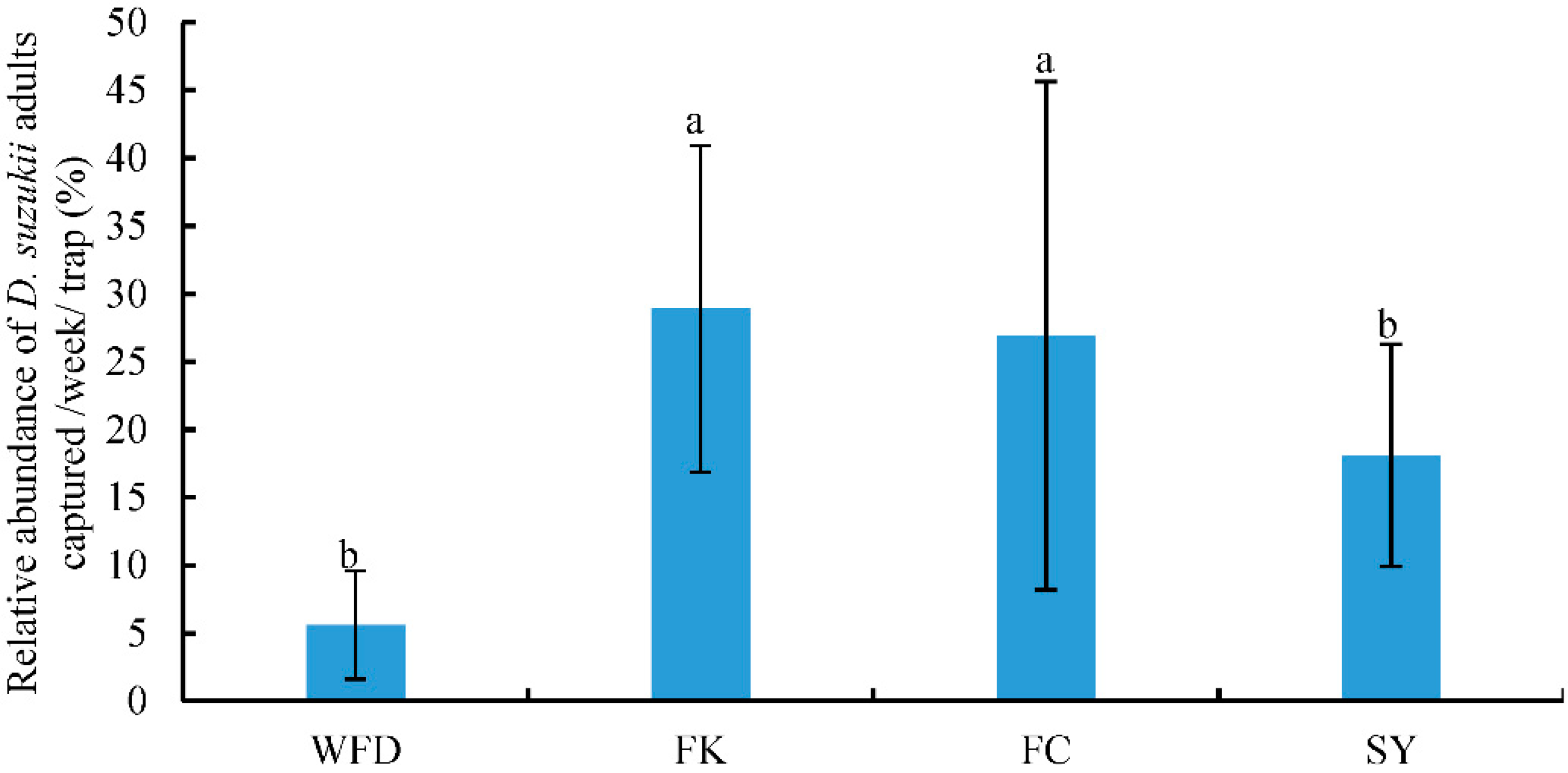
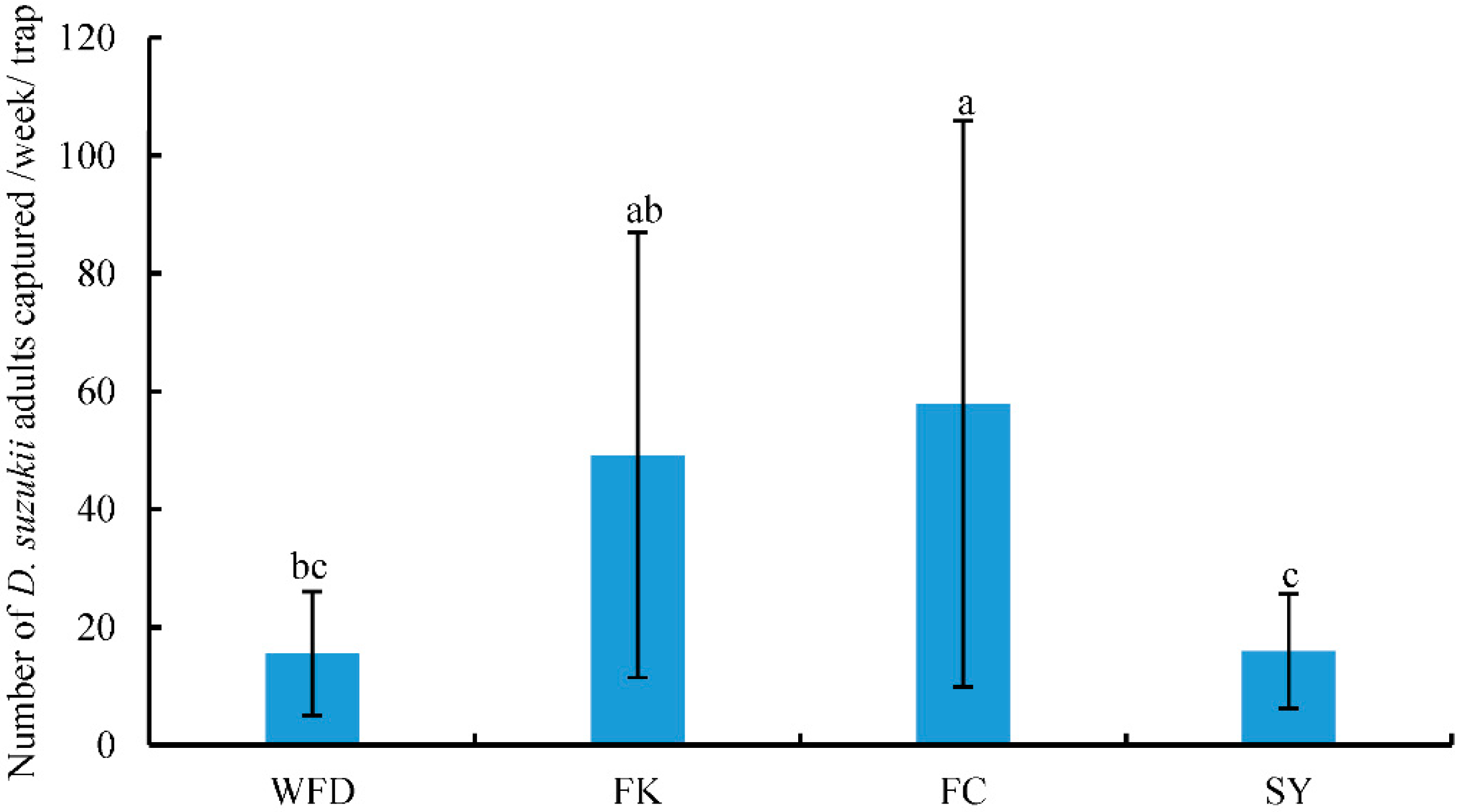
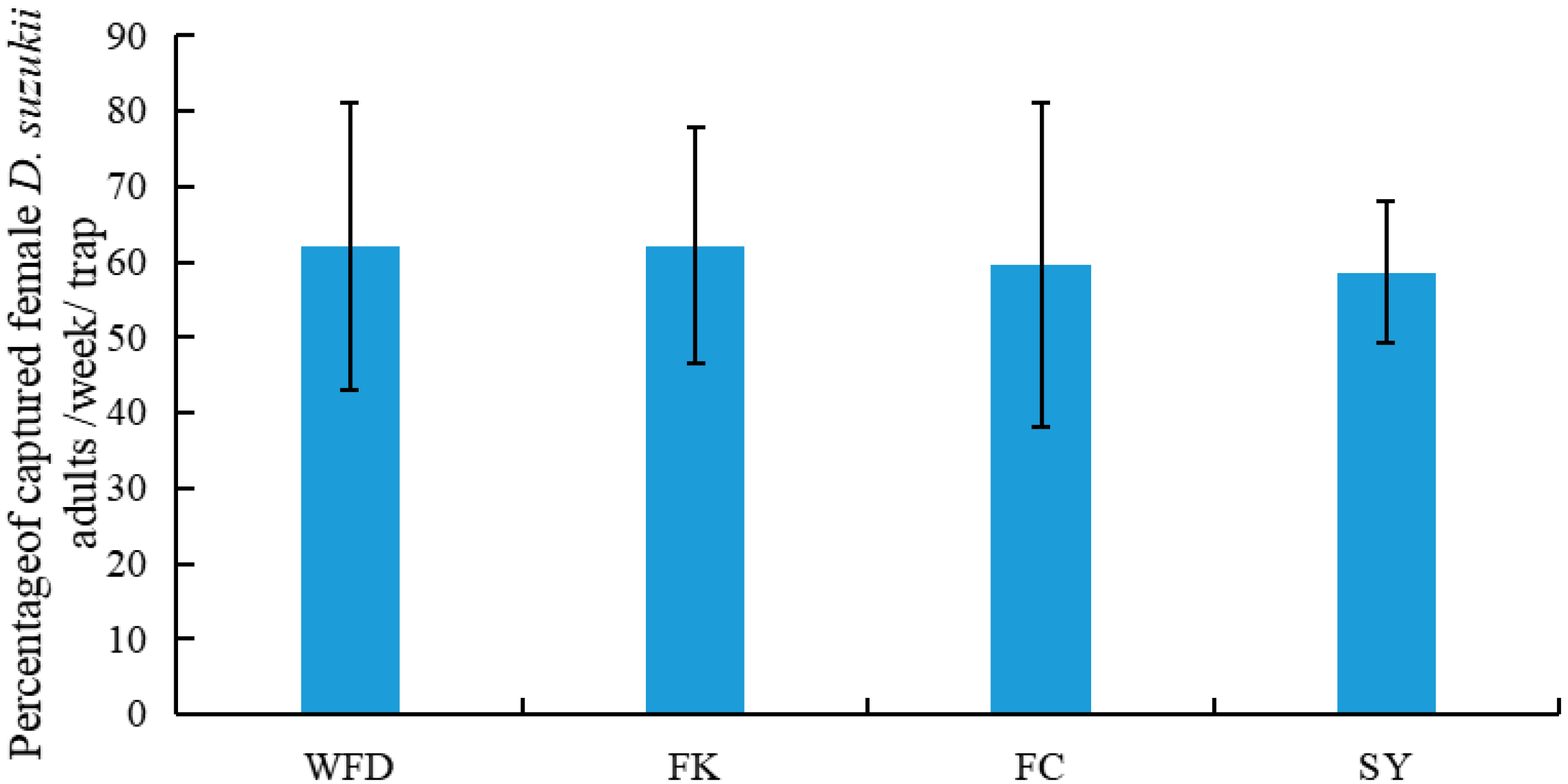
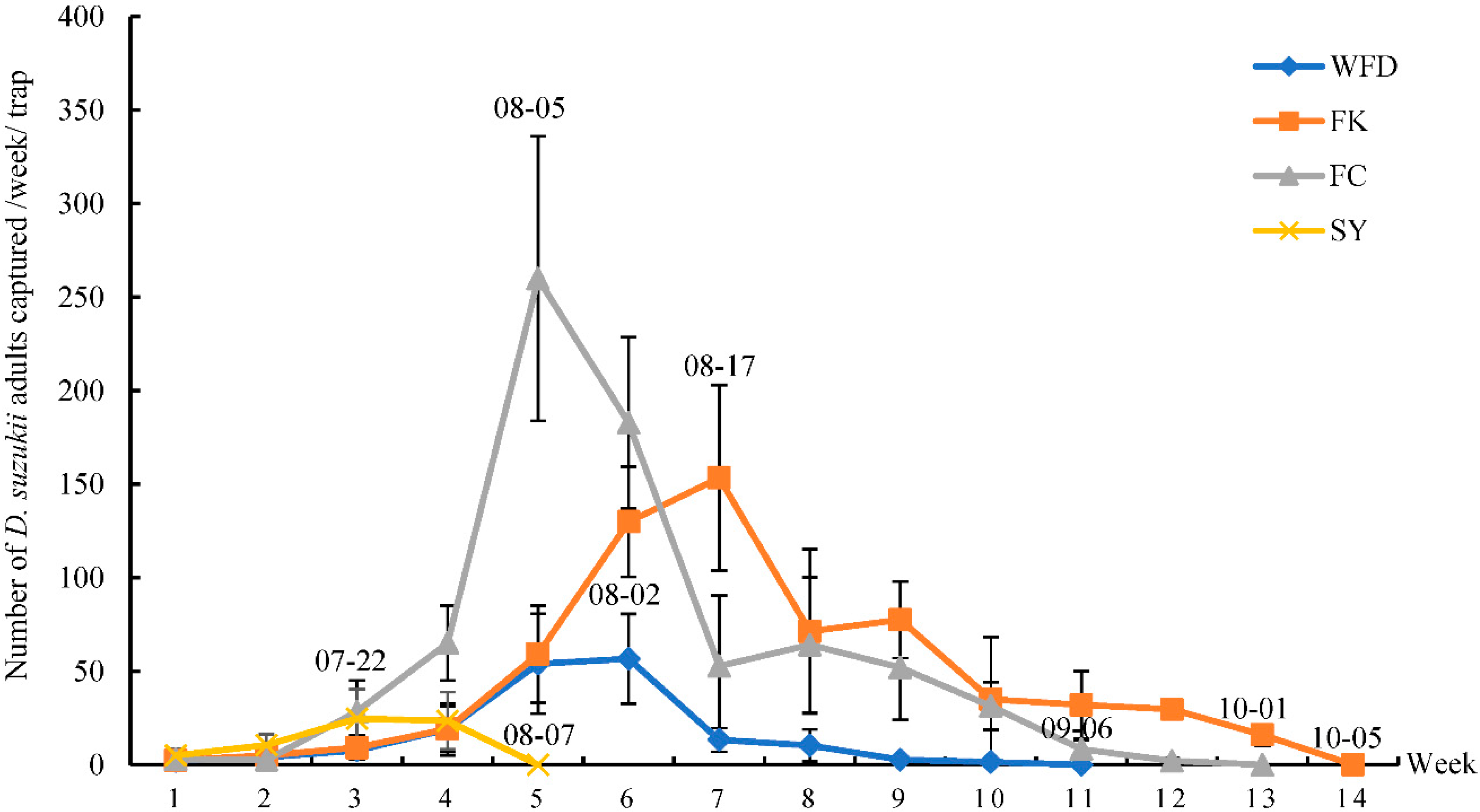
3.2. Captures of Adult D. suzukii in Sentinel Traps
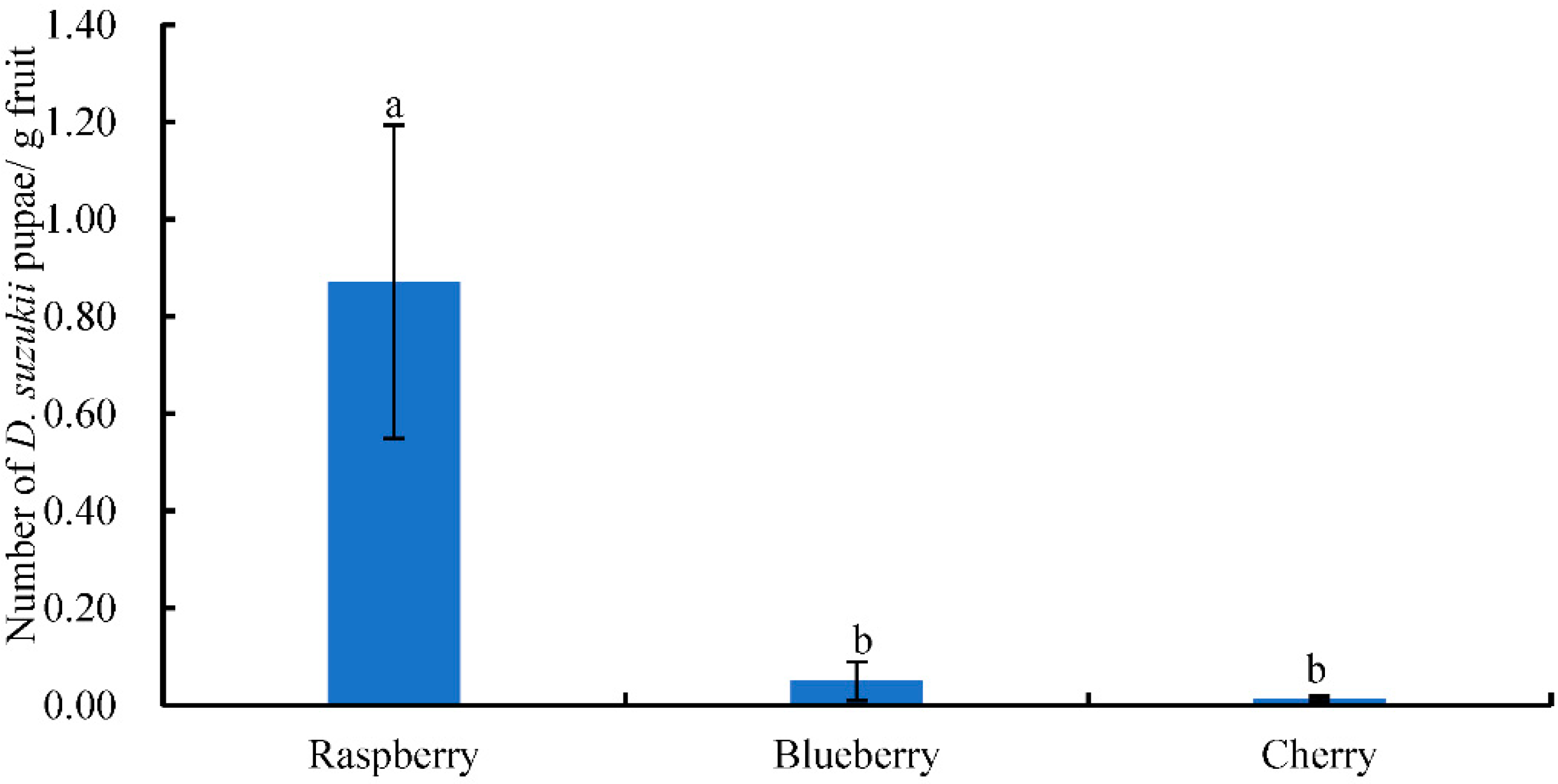
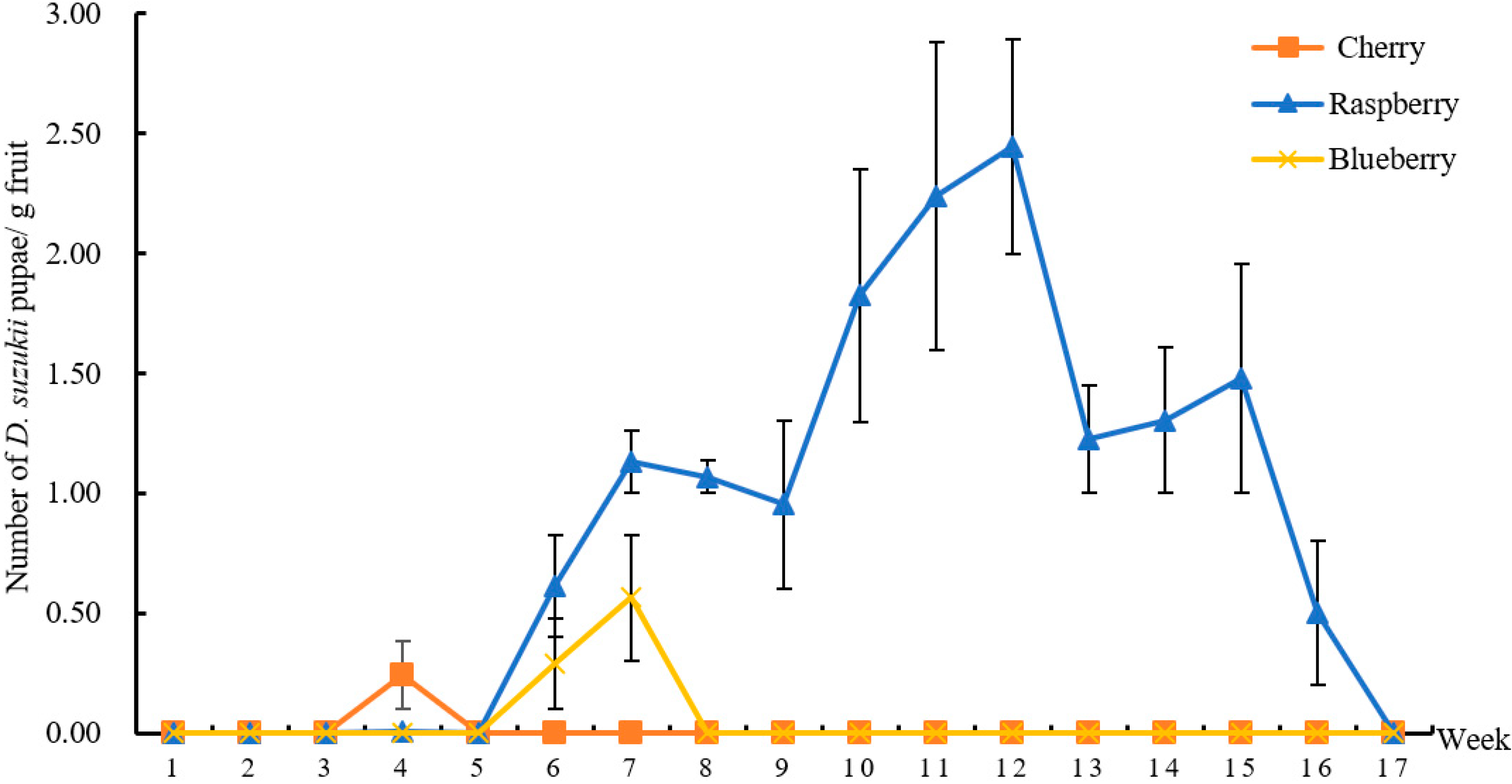
3.3. D. suzukii Larvae Numbers in Suspected Fruits
3.4. D. suzukii Surveys in Wild Host Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kanzawa, T. Studies on Drosophila suzukii Mats; Yamanashi Agricultural Experimental Station: Kofu, Japan, 1939; p. 49. [Google Scholar]
- Asplen, M.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 681, 469–494. [Google Scholar] [CrossRef]
- Cini, A.; Anfora, G.; Escudero-Colomar, L.A.; Grassi, A.; Santosuosso, U.; Seljak, G.; Papini, A. Tracking the invasion of the alien fruit pest Drosophila suzukii in Europe. J. Pest Sci. 2014, 87, 559–566. [Google Scholar] [CrossRef]
- Winkler, A.; Jung, J.; Kleinhenz, B.; Racca, P. Estimating temperature effects on Drosophila suzukii life cycle parameters. Agric. For. Entomol. 2021, 23, 361–377. [Google Scholar] [CrossRef]
- Peng, F.T. On some species of Drosophila from China. Annot. Zool. 1937, 16, 20–27. Available online: https://dl.ndl.go.jp/info:ndljp/pid/10853470 (accessed on 1 January 2020).
- Chung, Y.J. Collection of wild Drosophila on Quelpart Island, Korea. Drosoph. Inf. Serv. 1955, 29, 111. [Google Scholar]
- Kang, Y.S.; Moon, K.W. Drosophilid fauna of six regions near the demilitarized zone in Korea. Korean J. Zool. 1968, 11, 65–68. Available online: https://www.dbpia.co.kr/journal/articleDetail (accessed on 13 April 2022).
- Toda, M.J. Drosophilidae (Diptera) in Myanmar (Burma) VII. The Drosophila melanogaster species-group, excepting the D. montiumspecies-subgroup. Orient. Insects 1991, 25, 69–94. [Google Scholar] [CrossRef]
- Okada, T. New distribution records of the Drosophilids in the oriental region. Makunagi 1976, 8, 1–8. (In Japanese) [Google Scholar]
- Sidorenko, V.S. New and unrecorded species of Drosophilidae from Soviet Far East (Diptera, Brachycera). Spixiana 1992, 15, 93–95. Available online: https://www.biodiversitylibrary.org/part/66137 (accessed on 13 April 2022).
- Parshad, R.; Duggal, K.K. Drosophilidae of Kashmir, India. Drosoph. Inf. Serv. 1965, 40, 44. [Google Scholar]
- Zhao, C.; Li, P.; Xie, D.S.; Hu, C.H.; Xiong, Y.; He, L.Y. The seasonal abundance of Drosophila suzukii in orchards and seasonal variation in fruit damage caused by this pest. Chin. J. Appl. Entomol. 2017, 54, 724–729. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-KCZS201705004.htm (accessed on 13 April 2022).
- Liu, S.; Gao, H.H.; Lv, Z.Y.; Zhai, Y.F.; Dang, H.Y.; Li, Q. Oviposition Selection of Drosophila suzukii (Diptera: Drosophilidae) on Six Nectarine Cultivars and Its Correlation to the Physiological Characteristics of Nectarine Cultivars. North. Hortic. 2018, 21, 60–65. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-BFYY201821011.htm (accessed on 13 April 2022).
- Hauser, M.; Gaimari, S.; Damus, M. Drosophila suzukii new to North America. J. Fly Times 2009, 43, 12–15. Available online: https://www.researchgate.net/publication/292585271 (accessed on 13 April 2022).
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Beers, E.H.; Steenwyk, R.A.V.; Shearer, P.W.; Coates, W.W.; Grant, J.A. Developing Drosophila suzukii management programs for sweet cherry in the western United States. Pest Manag. Sci. 2011, 67, 1386–1395. [Google Scholar] [CrossRef]
- Calabria, G.; Máca, J.; Bächli, G.; Serra, L.; Pascual, M. First records of the potential pest species Drosophila suzukii, (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Anonymous. Normes OEPP-Evaluation biologique des produits phytosanitaires. Bulletion OEPP/EPPO Bulletin. 2013, 43, 386–388. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/epp.12261 (accessed on 20 December 2021).
- Liu, M.P.; Li, P.P.; Chen, P.; Wu, X.Y.; Zhao, M.; Huang, Y. Changes of EPPO (2019) quarantine pest list. Plant Quar. 2019, 33, 20–25. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-ZWJY201906005.htm (accessed on 13 April 2022).
- Depra, M.; Poppe, J.L.; Schmitz, H.J.; Toni, D.C.; Valente, V.L.S. The first records of the invasive pest Drosophila suzukii in the South American continent. J. Pest Sci. 2014, 87, 379–383. [Google Scholar] [CrossRef]
- Boughdad, A.; Haddi, K.; El Bouazzati, A.; Nassiri, A.; Tahiri, A.; El Anbri, C.; Eddaya, T.; Zaid, A.; Biondi, A. First record of the invasive spotted wing Drosophila infesting berry crops in Africa. J. Pest Sci. 2021, 94, 261–271. [Google Scholar] [CrossRef]
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The making of a pest: The evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proc. Biol. Sci. 2014, 281, 2013–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, A.; Tanaka, K.M.; Yew, J.Y.; Takahashi, A. Drosophila suzukii avoidance of microbes in oviposition choice. R. Soc. Open sci. 2021, 8, 201601. [Google Scholar] [CrossRef] [PubMed]
- Ioriatti, C.; Walton, V.; Dalton, D.; Anfora, G.; Grassi, A.; Maistri, S.; Mazzoni, V. Drosophila suzukii (Diptera: Drosophilidae) and its potential impact to wine grapes during harvest in two cool climate wine grape production regions. J. Econ. Entomol. 2015, 108, 1148–1155. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Dreves, A.J.; Cave, A.M.; Kawai, S.; Isaacs, R.; Miller, J.C.; Van Timmeren, S.; Bruck, D.J. Infestation of Wild and Ornamental Noncrop Fruits by Drosophila suzukii (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2015, 108, 117–129. [Google Scholar] [CrossRef]
- Little, C.M.; Chapman, T.W.; Kirk, H.N. Plasticity Is Key to Success of Drosophila suzukii (Diptera: Drosophilidae) Invasion. J. Insect Sci. 2020, 20, 1049–1055. [Google Scholar] [CrossRef]
- Schneberg, T.; Arsenault-Benoit, A.L.; Taylor, C.M.; Butler, B.R.; Hamby, K.A. Pruning of small fruit crops can affect habitat suitability for Drosophila suzukii. Agric. Ecosyst. Environ. 2020, 294, 106860. [Google Scholar] [CrossRef]
- Stewart, T.J.; Wang, X.G.; Andrew, M.; Daane, M.D. Factors limiting peach as a potential host for Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2014, 107, 1771–1779. [Google Scholar] [CrossRef] [Green Version]
- Bruck, D.J.; Bolda, M.; Tanigoshi, L.; Klick, J.; Kleiber, J.; Defrancesco, J.; Gerdeman, B.; Spitler, H. Laboratory and field comparisons of insecticides to reduce infestation of Drosophila suzukii in berry crops. Pest Manag. Sci. 2011, 67, 1375–1385. [Google Scholar] [CrossRef]
- Emiljanowicz, L.M.; Ryan, G.D.; Langille, A.; Newman, J. Development, reproductive output and population growth of the fruit fly pest Drosophila suzukii (Diptera: Drosophilidae) on artificial diet. J. Econ. Entomol. 2014, 107, 1392–1398. [Google Scholar] [CrossRef] [Green Version]
- Hannah, J.B.; Powell, S.J.; Douglas, G.P.; Glen, K.; Joseph, L. Using Volunteer-Based Networks to Track Drosophila suzukii (Diptera: Drosophilidae) an Invasive Pest of Fruit Crops. J. Integr. Pest Manag. 2012, 4, B1–B5. [Google Scholar] [CrossRef] [Green Version]
- Samantha, T.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Wiman, N.G.; Walton, V.M.; Dalton, D.T.; Anfora, G.; Burrack, H.J.; Chiu, J.C.; Daane, K.M.; Grassi, A.; Miller, B.; Tochen, S.; et al. Integrating temperature-dependent life table data into a matrix projection model for Drosophila suzukii population estimation. PLoS ONE 2014, 9, e106909. [Google Scholar] [CrossRef] [PubMed]
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zalom, F.G. Spotted wing drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef]
- Gress, B.E.; Zalom, F.G. Identifcation and risk assessment of spinosad resistance in a California population of Drosophila suzukii. Pest Manag. Sci. 2019, 75, 1270–1276. [Google Scholar] [CrossRef]
- Whitehouse, T.S.; Sial, A.A.; Schmidt, J.M. Natural enemy abundance in southeastern blueberry agroecosystems: Distance to edge and impact of management practices. Environ. Entomol. 2018, 47, 32–38. [Google Scholar] [CrossRef]
- Girod, P.; Borowiec, N.; Buffngton, M.; Chen, G.H.; Fang, Y.; Kimura, M.T.; Peris-Felipo, F.J.; Ris, N.; Wu, H.; Xiao, C.; et al. The parasitoid complex of D. suzukii and other fruit feeding Drosophila species in Asia. Sci. Rep. 2018, 8, 11839. [Google Scholar] [CrossRef]
- Bock, I.N.; Wheeler, M.R. The Drosophila melanogaster species group. Univ Tex Publ. 1972, 7213, 1–102. [Google Scholar]
- Abram, P.; Wang, X.G.; Hueppelsheuser, T.; Franklin, M.F.; Daane, K.M.; Lee, J.C.; Lue, C.-H.; Girod, P.; Carrillo, J.; Wong, W.H.L.; et al. A coordinated sampling and identification methodology for larval parasitoids of spotted-wing drosophila. J. Econ. Entomol. 2022, 115, toab237. [Google Scholar] [CrossRef]
- Guerrieri, E.; Giorgini, M.; Cascone, P.; Carpenito, S.; Achterberg, C. Species diversity in the parasitoid genus Asobara (Hymenoptera: Braconidae) from the native area of the fruit fly pest Drosophila suzukii (Diptera: Drosophilidae). PLoS ONE 2016, 11, e0147382. [Google Scholar] [CrossRef] [Green Version]
- Zerulla, F.N.; Schmidt, S.; Streitberger, M.; Zebitz, C.P.W.; Zelger, R. On the overwintering ability of Drosophila suzukii in South Tyrol. J. Berry. Res. 2015, 5, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Mazzetto, F.; Marchetti, E.; Amiresmaeili, N.; Sacco, D.; Tavella, L. Drosophila parasitoids in northern Italy and their potential to attack the exotic pest Drosophila suzukii. J. Pest Sci. 2015, 89, 1–14. [Google Scholar] [CrossRef]
- Tait, G.; Mermer, S.; Stockton, D.; Lee, J.; Avosani, S.; Abrieux, A.; Anfora, G.; Beers, E.; Biondi, A.; Burrack, H.; et al. Drosophila suzukii (Diptera: Drosophilidae): A Decade of Research Towards a Sustainable Integrated Pest Management Program. J. Econ. Entomol. 2021, 114, 1950–1974. [Google Scholar] [CrossRef] [PubMed]
- Thistlewood, H.M.A.; Gill, P.; Beers, E.H.; Shearer, P.W.; Walsh, D.B.; Rozema, B.M.; Acheampong, S.; Castagnoli, S.; Yee, W.L.; Smytheman, P.; et al. Spatial analysis of seasonal dynamics and overwintering of Drosophila suzukii (Diptera: Drosophilidae) in the Okanagan-Columbia Basin, 2010–2014. Environ. Entomol. 2018, 47, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, H.; Hagler, J.R.; Machtley, S.A.; Isaacs, R. Spotted wing drosophila (Drosophila suzukii) utilization and dispersal from the wild host Asian bush honeysuckle (Lonicera spp.). Agric. For. Entomol. 2019, 21, 149–158. [Google Scholar] [CrossRef]
- Biondi, A.; Wang, X.G.; Daane, K.M. Host preference of three Asian larval parasitoids to closely related Drosophila species: Implications for biological control of Drosophila suzukii. J. Pest Sci. 2021, 94, 273–283. [Google Scholar] [CrossRef]
- Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Lee, J.C.T. Biological Control of Spotted-Wing Drosophila: An update on promising agents. In Drosophila suzukii Management; Garcia, F.R.M., Ed.; Springer: Cham, Switzerland, 2020; pp. 143–168. [Google Scholar] [CrossRef]
- Carton, Y.; Bouletreau, M.; Alphen, J.J.M.V.; Lenteren, J.C.V. The Drosophila parasitic wasps. Genet. Biol. Drosoph. 1986, 3, 347–394. [Google Scholar]
- Lewald, K.M.; Abrieux, A.; Wilson, D.A.; Lee, Y.; Andreazza, F.; Beers, E.H. Population genomics of Drosophila suzukii reveal longitudinal population structure and signals of migrations in and out of the continental United States. G3-Genes. Genom. Genet. 2021, 343, 1–11. [Google Scholar] [CrossRef]
- Adrion, J.R.; Athanasios, K.; Marta, P.; Burrack, H.J.; Haddad, N.M.; Bergland, A.O.; Machado, H.; Sackton, T.B.; Schlenke, T.A.; Watada, M.; et al. Drosophila suzukii: The Genetic Footprint of a Recent, Worldwide Invasion. Mol. Biol. Evol. 2014, 31, 3148–3163. [Google Scholar] [CrossRef] [Green Version]
- Daane, K.M.; Wang, X.G.; Biondi, A.; Miller, B.; Miller, J.C.; Riedl, H.; Shearer, P.W.; Guerrieri, E.; Giorgini, M.; Buffington, M.; et al. First exploration of parasitoids of Drosophila suzukii in South Korea as potential classical biological agents. J. Pest Sci. 2016, 89, 823–835. [Google Scholar] [CrossRef]
- Giorgini, M.; Wang, X.G.; Wang, Y.; Chen, F.S.; Hougardy, E.; Zhang, H.M.; Chen, Z.-Q.; Chen, H.-Y.; Liu, C.-X.; Cascone, P.; et al. Exploration for native parasitoids of Drosophila suzukii in China reveals a diversity of parasitoid species and narrow host range of the dominant parasitoid. J. Pest Sci. 2019, 92, 509–522. [Google Scholar] [CrossRef]
- Daane, K.M.; Wang, X.G.; Hogg, B.N.; Biondi, A. Potential host ranges of three Asian larval parasitoids of Drosophila suzukii. J. Pest Sci. 2021, 94, 1171–1182. [Google Scholar] [CrossRef]
- Puppato, S.; Grassi, A.; Pedrazzoli, F.; Cristofaro, A.D.; Ioriatti, C. First Report of Leptopilina japonica in Europe. Insects 2020, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Abram, P.K.; McPherson, A.E.; Kula, R.; Hueppelsheuser, T.; Thiessen, J.; Perlman, S.J.; Curtis, C.I.; Fraser, J.L.; Tam, J.; Carrillo, J.; et al. New records of Leptopilina, Ganaspis, and Asobara species associated with Drosophila suzukii in North America, including detections of L. japonica and G. brasiliensis. J. Hymenopt. Res. 2020, 78, 1–17. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survey Location | Collection Site | Host Plant Habitat | Coordinates | Collection Date | Mean Temperature (°C) | Mean Humidity (%) | Mean Rainfall (mm) |
|---|---|---|---|---|---|---|---|
| Wafangdian | Delisi Orchard | Commercial cherry farm | N 39°47′ E 122°03′ | Jun 21–Oct 31 | 20.51 | 75.14 | 2.91 |
| Fengcheng | Enhue Orchard | Commercial blueberry farm | N 40°24′ E 123°57′ | Jun 22–Oct 31 | 19.06 | 77.54 | 4.64 |
| Fengcheng | Fenghuangshan | Natural forest | N 40°24′ E 124°4′ | Aug 2–Sep 24 | 21.53 | 81.15 | 3.96 |
| Shenyang | Shenyang Agricultural University | Research raspberry farm | N 41°49′ E 123°34′ | Jul 2–Aug 26 | 24.97 | 63.74 | 6.93 |
| Faku | Maanshan Orchard | Commercial raspberry farm | N 42°26′ E 122°52′ | Jun 26–Oct 31 | 18.67 | 74.59 | 4.21 |
| Fushun | Huangqi City | Natural forest (wild raspberry) | N 41°51′ E 123°54′ | Jul 18 | 22.54 | 76.16 | 0.08 |
| Fushun | Wendao Forest | State owned forest | N 41°8′ E 124°2′ | Sep 7 | 19.85 | 88.11 | 5.68 |
| Collection Location 1 | Collection Date | Wild Berry Species | Was D. suzukii Present? |
|---|---|---|---|
| FC | July 8–October 1 | Actinidia arguta (Sieb. & Zucc) Planch. Ex Miq. | Yes |
| Rubus crataegifolius Bunge | Yes | ||
| Lonicera maackii (Rupr.) Maxim. | No | ||
| SY | July 9–August 26 | Bothrocaryum controversum (Hemsl.) Pojark | No |
| Cerasus tomentosa (Thunb.) Wall. | No | ||
| Padus racemosa (L.) Gilib. | No | ||
| Rhamnus davurica Pall. | No | ||
| Sambucus williamsii Hance | No | ||
| Viburnum dilatatum Thunberg | No | ||
| FS | July 18 | Rubus idaeus L. | Yes |
| September 7 | Bothrocaryum controversum (Hemsl.) Pojark | No | |
| Cerasus tomentosa (Thunb.) Wall. | No | ||
| Hippophae rhamnoides L. | No | ||
| Padus racemosa (L.) Gilib. | No | ||
| Rhamnus davurica Pall. | No | ||
| Rubus crataegifolius Bunge | Yes | ||
| Viburnum dilatatum Thunberg | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zheng, Y.; Fan, L.; Wang, W. Surveys of Drosophila suzukii (Diptera: Drosophilidae) and Its Host Fruits and Associated Parasitoids in Northeastern China. Insects 2022, 13, 390. https://doi.org/10.3390/insects13040390
Wang J, Zheng Y, Fan L, Wang W. Surveys of Drosophila suzukii (Diptera: Drosophilidae) and Its Host Fruits and Associated Parasitoids in Northeastern China. Insects. 2022; 13(4):390. https://doi.org/10.3390/insects13040390
Chicago/Turabian StyleWang, Jue, Yanan Zheng, Lichun Fan, and Weitao Wang. 2022. "Surveys of Drosophila suzukii (Diptera: Drosophilidae) and Its Host Fruits and Associated Parasitoids in Northeastern China" Insects 13, no. 4: 390. https://doi.org/10.3390/insects13040390
APA StyleWang, J., Zheng, Y., Fan, L., & Wang, W. (2022). Surveys of Drosophila suzukii (Diptera: Drosophilidae) and Its Host Fruits and Associated Parasitoids in Northeastern China. Insects, 13(4), 390. https://doi.org/10.3390/insects13040390