
Landscape Effects on the Cabbage Seedpod Weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), and on Its Parasitoid, Trichomalus perfectus (Hymenoptera: Pteromalidae), in Canola

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Methods: Sweep Nets for the CSW and Pod Collection for T. perfectus
2.2. Emergence Boxes (CSW and Parasitoids)
2.3. Landscape Predictors Measures
2.4. Statistical Analysis
3. Results
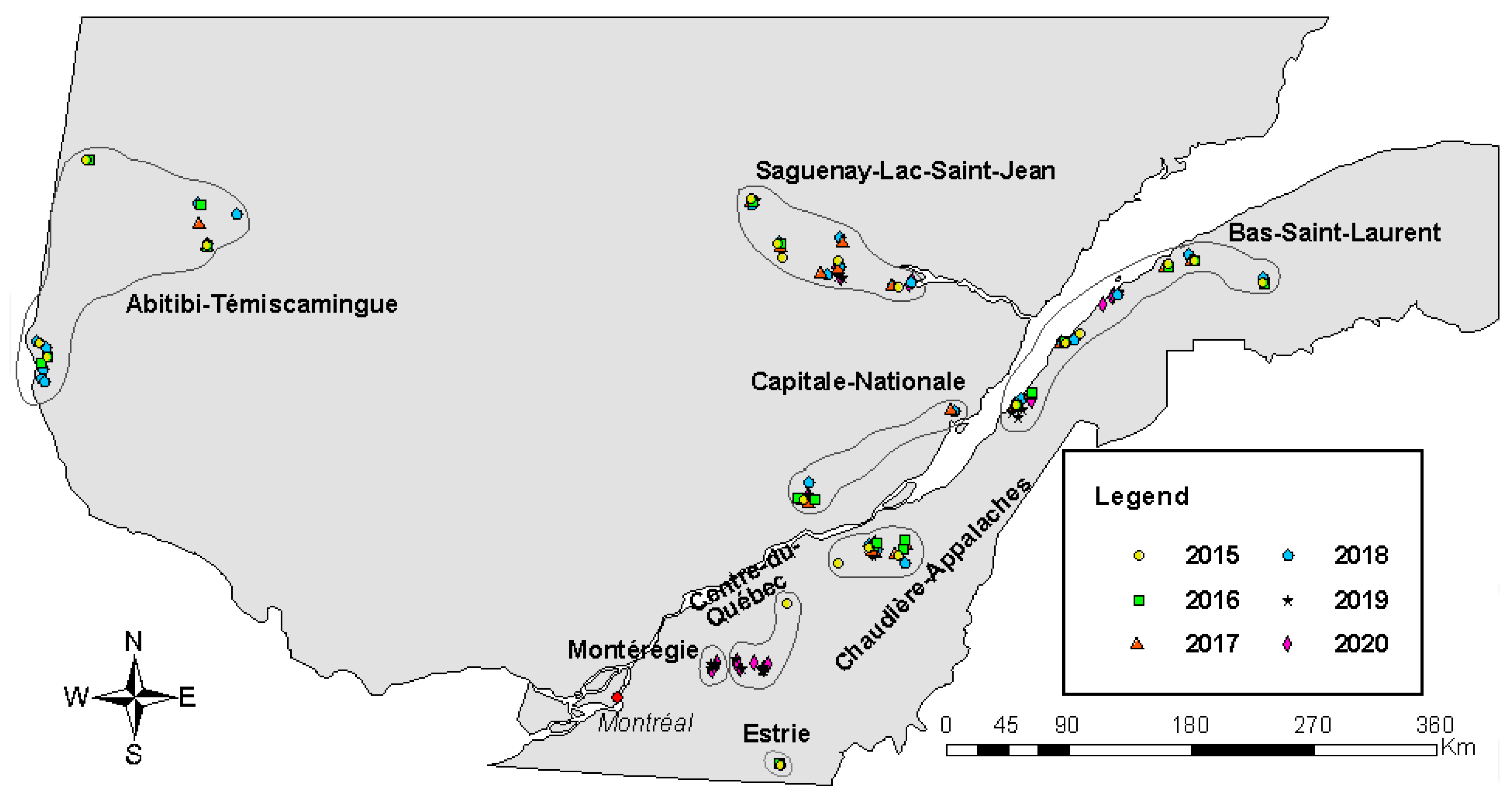
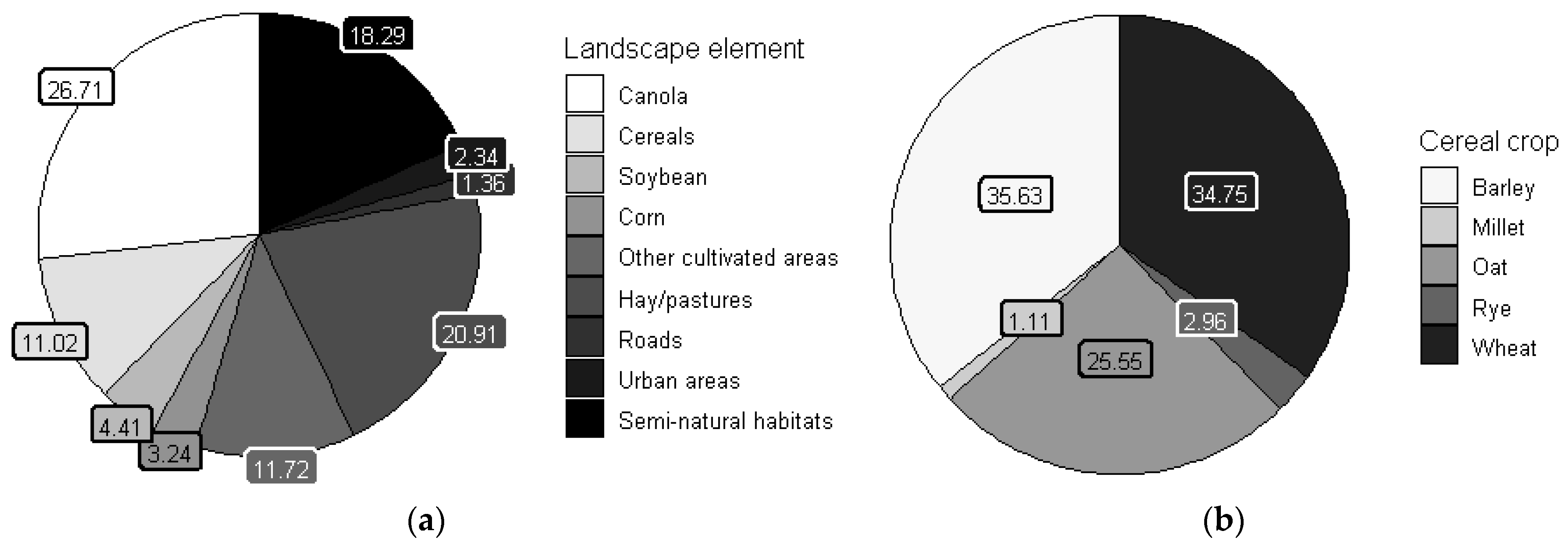
3.1. Overall Situation
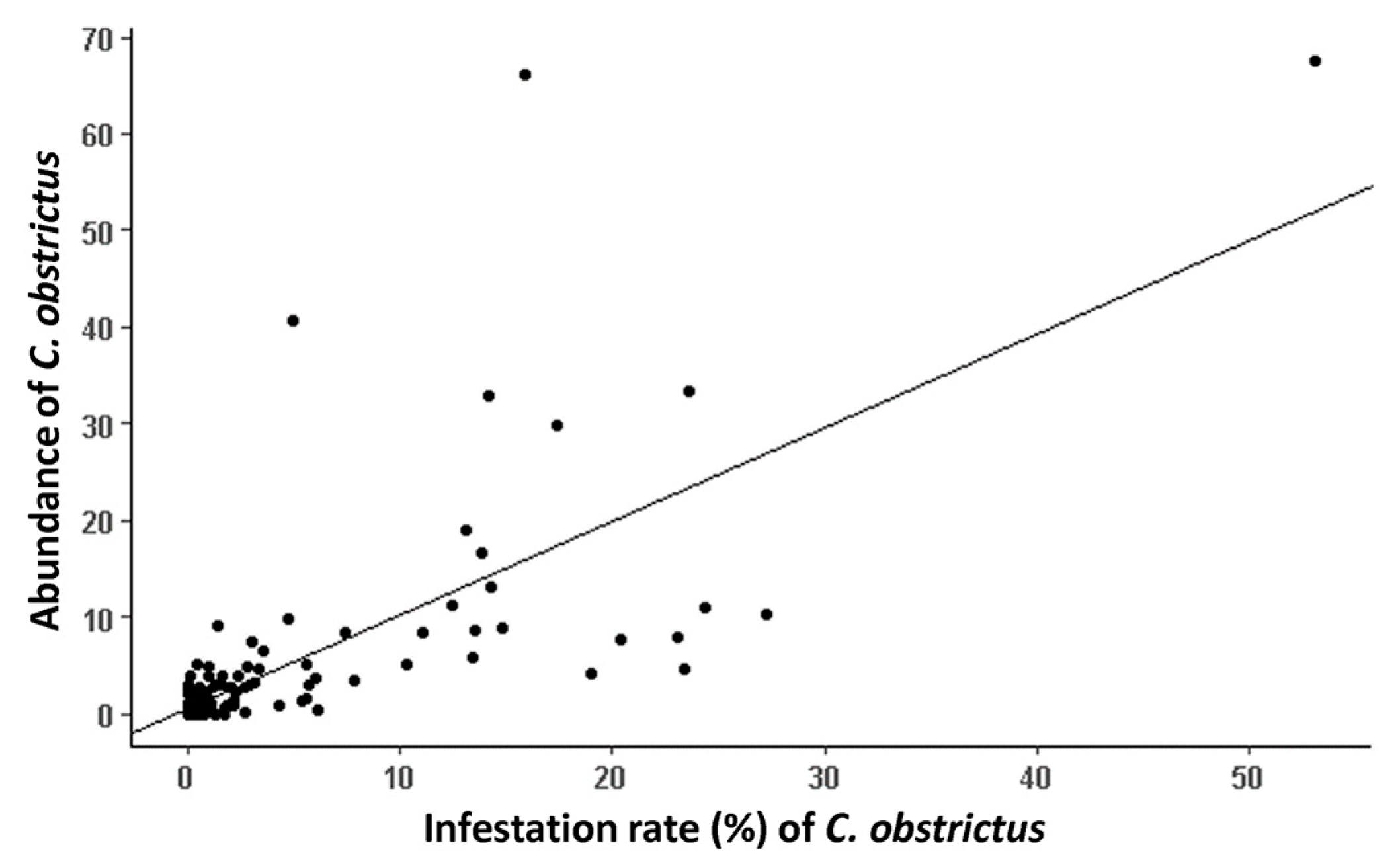
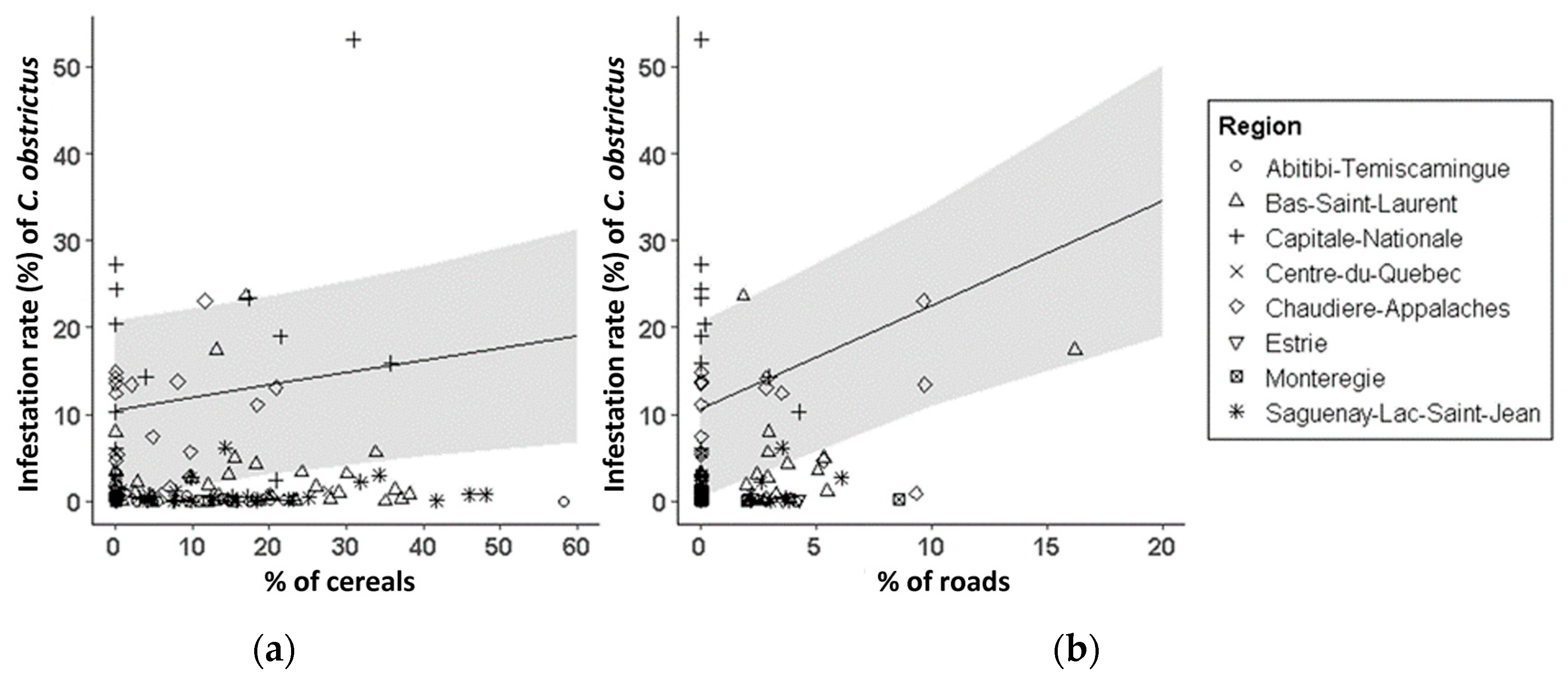
3.2. CSW Abundance and Infestation
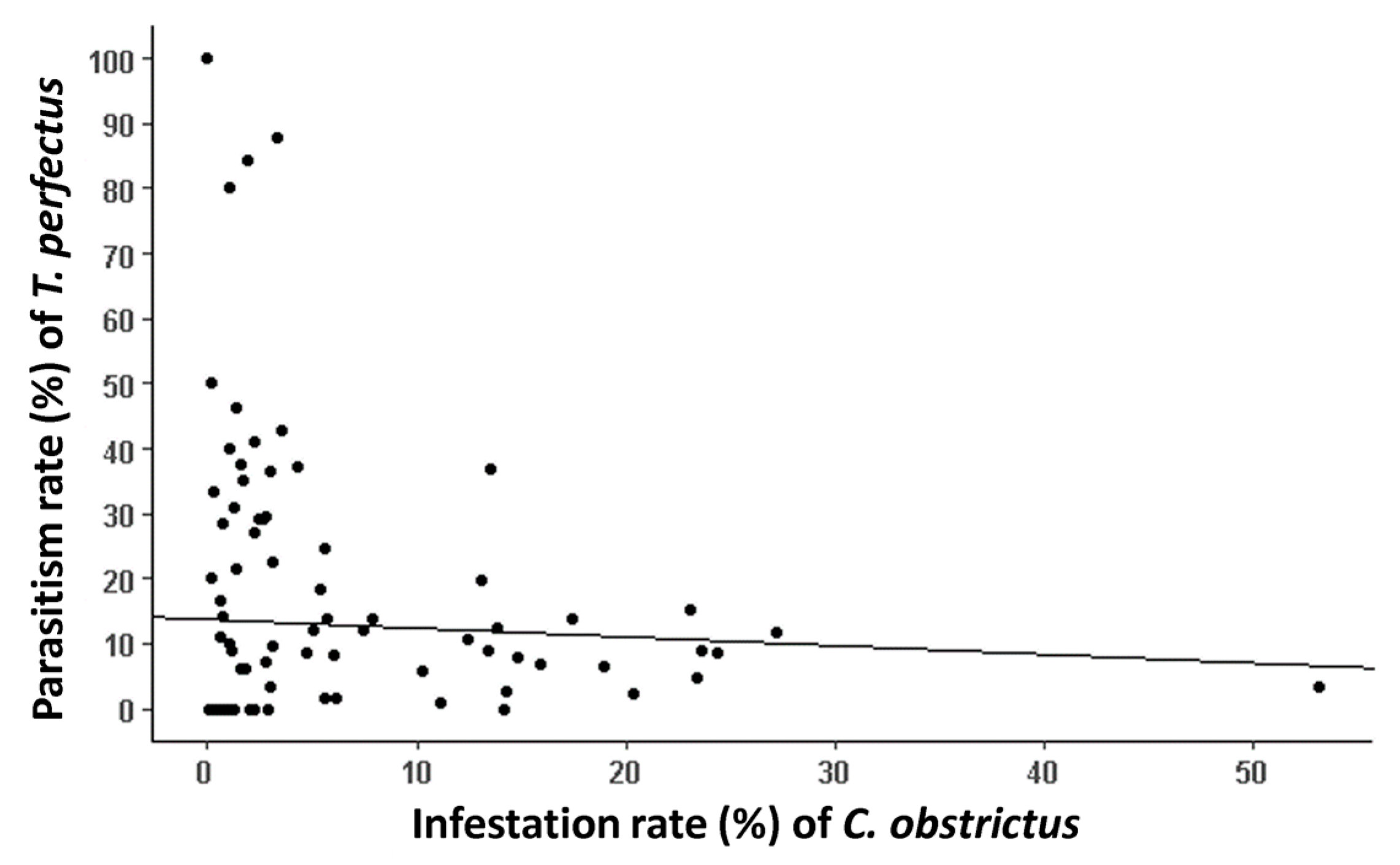
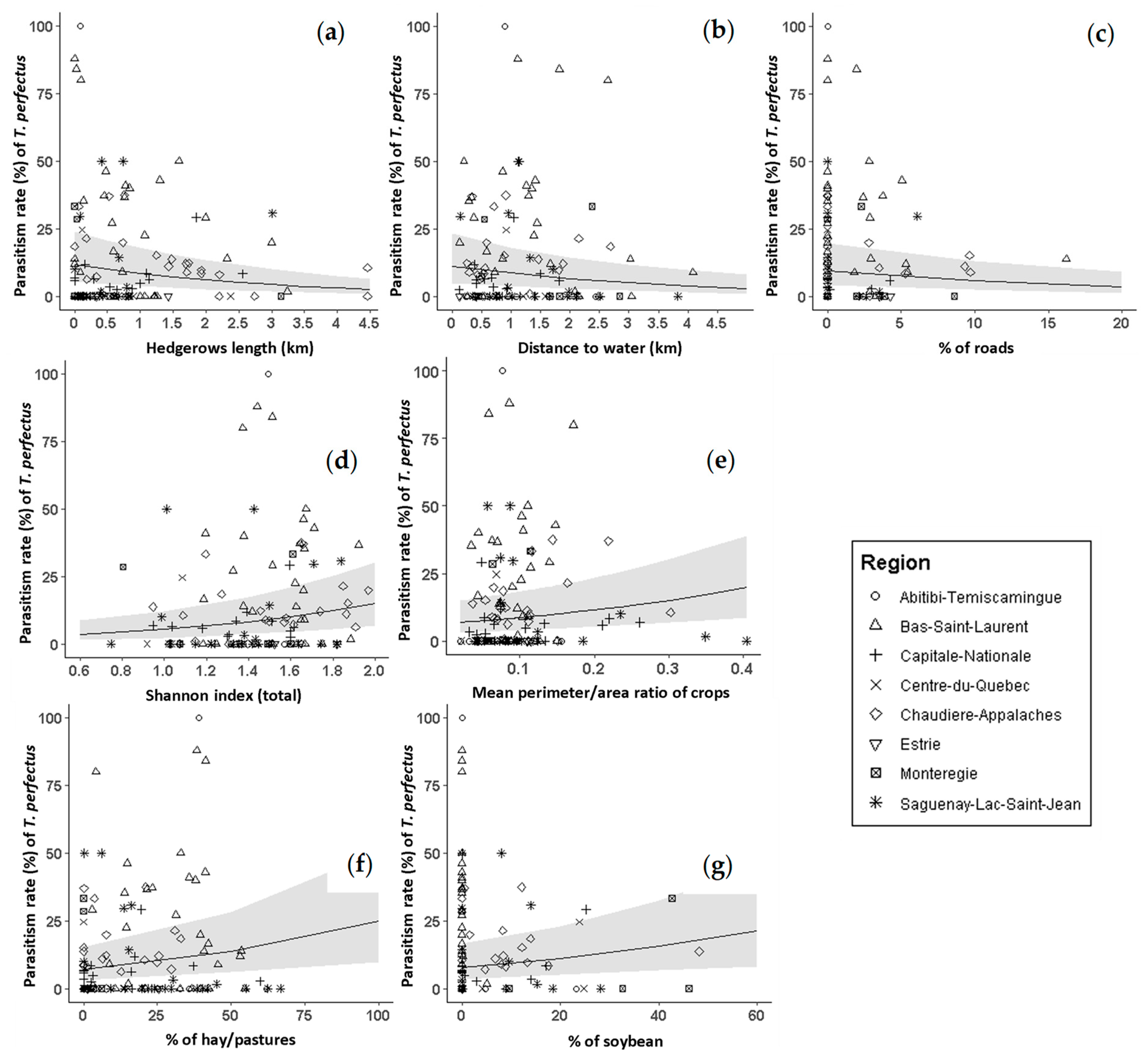
3.3. Trichomalus perfectus Parasitism Rate
4. Discussion
4.1. Landscape Effects on CSW Infestation and Abundance
4.2. Landscape Effects on T. perfectus Parasitism Rate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rieger, M.A.; Lamond, M.; Preston, C.; Powles, S.B.; Roush, R.T. Pollen-mediated movement of herbicide resistance between commercial canola fields. Science 2002, 296, 2386–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statistics Canada. Available online: https://www150.statcan.gc.ca/t1/tbl1/fr/cv.action?pid=3210035901 (accessed on 15 February 2023).
- Statista. Available online: https://www.statista.com/statistics/263930/worldwide-production-of-rapeseed-by-country/ (accessed on 15 February 2023).
- Bonnemaison, L. Le charançon des siliques (Ceuthorrhynchus assimilis Payk.). Ann. Épiphyt. 1957, 8, 387–543. [Google Scholar]
- Dmoch, J. The dynamics of a population of the cabbage seedpod weevil (Ceutorhynchus assimilis Payk.) and the development of winter rape. Part I. Pol. J. Ecol. 1965, 8, 249–287. [Google Scholar]
- Buntin, G.D. Damage loss assessment and control of the cabbage seedpod weevil (Coleoptera: Curculionidae) in winter canola using insecticides. J. Econ. Entomol. 1999, 92, 220–227. [Google Scholar] [CrossRef]
- Dosdall, L.M.; Moisey, D.; Cárcamo, H.; Dunn, R. Cabbage Seedpod Weevil Factsheet; Agdex 622-21; Alberta Agriculture, Food and Rural Development: Edmonton, AB, Canada, 2001; p. 4. [Google Scholar]
- McLeod, J.H. Cabbage seedpod weevil—Ceutorhynchus assimilis (Payk.) Curculionidae. In A Review of the Biological Control Attempts against Insects and Weeds in Canada; McLeod, J.H., McGugan, B.M., Coppel, H.C., Eds.; Commonwealth Agricultural Bureaux (CAB): Farnham Royal, UK, 1962; pp. 5–6. [Google Scholar]
- Cárcamo, H.A.; Dosdall, L.; Dolinski, M.; Olfert, O.; Byers, J.R. The cabbage seedpod weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae)—A review. J. Entomol. Soc. Br. Columbia 2001, 98, 201–210. [Google Scholar]
- Brodeur, J.; Leclerc, L.A.; Fournier, M.; Roy, M. Cabbage seedpod weevil (Coleoptera: Curculionidae): New pest of canola in northeastern North America. Can. Entomol. 2001, 133, 709–711. [Google Scholar] [CrossRef]
- Mason, P.G.; Baute, T.; Olfert, O.; Roy, M. Cabbage seedpod weevil, Ceutorhynchus obstrictus (Marsham) (Coleoptera: Curculionidae) in Ontario and Quebec. J. Entomol. Soc. Ont. 2004, 134, 107–113. [Google Scholar]
- Ni, X.; McCaffrey, J.P.; Stoltz, R.L.; Harmon, B.L. Effects of postdiapause adult diet and temperature on oogenesis of the cabbage seedpod weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 1990, 83, 2246–2251. [Google Scholar] [CrossRef]
- Cárcamo, H.A.; Dosdall, L.M.; Johnson, D.; Olfert, O. Evaluation of foliar and seed treatments for control of the cabbage seedpod weevil (Coleoptera: Curculionidae) in canola. Can. Entomol. 2005, 137, 476–487. [Google Scholar] [CrossRef] [Green Version]
- Williams, I.H. (Ed.) The major insect pests of oilseed rape in Europe and their management—An overview. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Springer: Heidelberg, Germany; London, UK; New York, NY, USA, 2010; pp. 1–44. [Google Scholar]
- Murchie, A.K. Parasitoids of Cabbage Seed Weevil and Brassica Pod Midge in Oilseed Rape. Ph.D. Thesis, University of Keele, Keele, UK, 1996. [Google Scholar]
- Ulber, B.; Williams, I.H.; Klukowski, Z.; Luik, A.; Nilsson, C. Parasitoids of oilseed rape pests in Europe: Key species for conservation biocontrol. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Williams, I.H., Ed.; Springer: Heidelberg, Germany; London, UK; New York, NY, USA, 2010; pp. 45–76. [Google Scholar]
- Kovács, G.; Kaasik, R.; Metspalu, L.; Williams, I.H.; Luik, A.; Veromann, E. Could Brassica rapa, Brassica juncea and Sinapis alba facilitate the control of the cabbage seed weevil in oilseed rape crops? Biol. Control 2013, 65, 124–129. [Google Scholar] [CrossRef]
- Haye, T.; Mason, P.G.; Gillespie, D.R.; Miall, J.H.; Gibson, G.A.P.; Diaconu, A.; Brauner, A.M.; Kuhlmann, U. Determining the host specificity of the biological control agent Trichomalus perfectus (Hymenoptera: Pteromalidae): The importance of ecological host range. Biocontrol Sci. Technol. 2015, 25, 21–47. [Google Scholar] [CrossRef]
- Alford, G.V.; Williams, I.H.; Murchie, A.K.; Walters, K.F.A. The status and potential of parasitoids of seed weevil and pod midge on winter oilseed rape. In HGCA Oilseeds Project Report No. OS14; Home-Grown Cereals Authority: London, UK, 1995; p. 89. [Google Scholar]
- Dmoch, J. Kairomones and searching behavior of Trichomalus perfectus Walker. IOBC/WPRS Bull. 1998, 21, 171–176. [Google Scholar]
- Murchie, A.K.; Williams, I.H. A bibliography of the parasitoids of the cabbage seed weevil (Ceutorhynchus assimilis Payk.). IOBC/WPRS Bull. 1998, 21, 163–169. [Google Scholar]
- Williams, I.H. Parasitoids of cabbage seed weevil. In Biocontrol of Oilseed Rape Pests; Alford, D.V., Ed.; Blackwell Science Ltd.: Oxford, UK, 2003; pp. 97–112. [Google Scholar]
- Mason, P.G.; Miall, J.H.; Bouchard, P.; Gillespie, D.R.; Broadbent, A.B.; Gibson, G.A.P. The parasitoid communities associated with an invasive canola pest, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), in Ontario and Quebec, Canada. Can. Entomol. 2011, 143, 524–537. [Google Scholar] [CrossRef]
- Boquel, S.; D’Ottavio, M.; Labrie, G.; Lucas, E. CÉROM—Centre de Recherche sur les Grains Inc.: Saint-Mathieu-de-Beloeil, QC, Canada, manuscript in preparation.
- Kovács, G.; Kaasik, R.; Treier, K.; Luik, A.; Veromann, E. Do different field bordering elements affect cabbage seed weevil damage and its parasitism rate differently in winter oilseed rape? IOBC/WPRS Bull. 2016, 116, 75–80. [Google Scholar]
- Kovács, G.; Kaasik, R.; Lof, M.E.; van der Werf, W.; Kaart, T.; Holland, J.M.; Luik, A.; Veromann, E. Effects of land use on infestation and parasitism rates of cabbage seed weevil in oilseed rape. Pest Manag. Sci. 2019, 75, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.S.; Birkhofer, K.; Hanson, H.I.; Hedlund, K. Landscape configuration affects herbivore-parasitoid communities in oilseed rape. J. Pest Sci. 2018, 91, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Root, R.B. Organization of a plant-arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Rand, T.A.; Waters, D.K.; Blodgett, S.L.; Knodel, J.J.; Harris, M.O. Increased area of a highly suitable host crop increases herbivore pressure in intensified agricultural landscapes. Agric. Ecosyst. Environ. 2014, 186, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Maisonhaute, J.É.; Labrie, G.; Lucas, E. Direct and indirect effects of the spatial context on the natural biocontrol of an invasive crop pest. Biol. Control 2017, 106, 64–76. [Google Scholar] [CrossRef]
- Ulmer, B.J.; Dosdall, L.M. Spring emergence biology of the cabbage seedpod weevil (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 2006, 99, 64–69. [Google Scholar] [CrossRef]
- Veromann, E.; Williams, I.H.; Kaasik, R.; Luik, A. Potential of parasitoids to control populations of the weevil Ceutorhynchus obstrictus (Marsham) on winter oilseed rape. Int. J. Pest Manag. 2011, 57, 85–92. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. How do landscape composition and configuration, organic farming and fallow strips affect the diversity of bees, wasps and their parasitoids? J. Anim. Ecol. 2010, 79, 491–500. [Google Scholar] [CrossRef]
- Winkler, K.; Wäckers, F.; Bukovinszkine-Kiss, G.; Van Lenteren, J. Sugar resources are vital for Diadegma semiclausum fecundity under field conditions. Basic Appl. Ecol. 2006, 7, 133–140. [Google Scholar] [CrossRef]
- Géneau, C.E.; Wäckers, F.L.; Luka, H.; Daniel, C.; Balmer, O. Selective flowers to enhance biological control of cabbage pests by parasitoids. Basic Appl. Ecol. 2012, 13, 85–93. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Wratten, S.D. Improved fitness of aphid parasitoids receiving resource subsidies. Ecology 2004, 85, 658–666. [Google Scholar] [CrossRef] [Green Version]
- Lavandero, B.; Wratten, S.D.; Didham, R.K.; Gurr, G. Increasing floral diversity for selective enhancement of biological control agents: A double-edged sward? Basic Appl. Ecol. 2006, 7, 236–243. [Google Scholar] [CrossRef]
- Ellers, J.; Van Alphen, J.J.; Sevenster, J.G. A field study of size–fitness relationships in the parasitoid Asobara tabida. J. Anim. Ecol. 1998, 67, 318–324. [Google Scholar] [CrossRef]
- van Nouhuys, S. Effects of habitat fragmentation at different trophic levels in insect communities. Ann. Zool. Fenn. 2005, 42, 433–447. [Google Scholar]
- Dosdall, L.M.; Ulmer, B.J.; Gibson, G.A.P.; Cárcamo, H.A. The spatio-temporal distribution dynamics of the cabbage seedpod weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), and its larval parasitoids in canola in western Canada. Biocontrol Sci. Technol. 2006, 16, 987–1006. [Google Scholar] [CrossRef]
- Muller, F.J.; Baur, H.; Gibson, G.A.; Mason, P.G.; Kuhlmann, U. Review of the species of Trichomalus (Chalcidoidea: Pteromalidae) associated with Ceutorhynchus (Coleoptera: Curculionidae) host species of European origin. Can. Entomol. 2007, 139, 643–657. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 September 2022).
- Sabin, T.E.; Stafford, S.G. Assessing the Need for Transformation of Response Variables; Special Publication 20; Forest Research Laboratory, Oregon State University: Corvallis, OR, USA, 1990; p. 31. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical-Theoretical Approach, 2nd ed.; Springer: Berlin, Germany, 2002; p. 488. [Google Scholar]
- Muñoz, P.T.; Torres, F.P.; Megías, A.G. Effects of roads on insects: A review. Biodivers. Conserv. 2015, 24, 659–682. [Google Scholar] [CrossRef]
- Tansey, J.A.; Dosdall, L.M.; Keddie, A.; Olfert, O. Flight activity and dispersal of the cabbage seedpod weevil (Coleoptera: Curculionidae) are related to atmospheric conditions. Environ. Entomol. 2010, 39, 1092–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desroches, C.; Bouchard, P.; Labrie, G.; Lucas, E. Assemblage of Ceutorhynchinae Weevils Associated with Brassicaceae in Quebec (Canada) Agroecosystems. Environ. Entomol. 2022, 52, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Doucette, C.F. Host plants of the cabbage seedpod weevil. J. Econ. Entomol. 1947, 40, 838–840. [Google Scholar] [CrossRef]
- Dosdall, L.M.; Moisey, D.W. Developmental biology of the cabbage seedpod weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), in spring canola, Brassica napus, in western Canada. Ann. Entomol. Soc. Am. 2004, 97, 458–465. [Google Scholar] [CrossRef]
- Noordijk, J.; Schaffers, A.; Sýkora, K.V. Diversity of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) in roadside verges with grey hair-grass vegetation. Eur. J. Entomol. 2008, 105, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Noordijk, J.; Schaffers, A.P.; Heijerman, T.; Sýkora, K.V. Using movement and habitat corridors to improve the connectivity for heathland carabid beetles. J. Nat. Conserv. 2011, 19, 276–284. [Google Scholar] [CrossRef]
- Knapp, M.; Saska, P.; Knappová, J.; Vonička, P.; Moravec, P.; Kůrka, A.; Anděl, P. The habitat-specific effects of highway proximity on ground-dwelling arthropods: Implications for biodiversity conservation. Biol. Conserv. 2013, 164, 22–29. [Google Scholar] [CrossRef]
- Dániel-Ferreira, J.; Berggren, Å.; Bommarco, R.; Wissman, J.; Öckinger, E. Bumblebee queen mortality along roads increase with traffic. Biol. Conserv. 2022, 272, 109643. [Google Scholar] [CrossRef]
- Anderson, A.B.; Jenkins, C.N. Applying Nature’s Design: Corridors as a Strategy for Biodiversity Conservation; Columbia University Press: New York, NY, USA, 2006; p. 231. [Google Scholar]
- Schaffers, A.P.; Raemakers, I.P.; Sýkora, K.V. Successful overwintering of arthropods in roadside verges. J. Insect Conserv. 2012, 16, 511–522. [Google Scholar] [CrossRef] [Green Version]
- Zoghlami, S.; Bergeron, M.; Moore, H.; Labrie, G.; Pageau, D.; Robert, L.; Tardif, É.; Tremblay, G. Guide de Production du Canola; Fédération des Producteurs de Cultures Commerciales du Québec: Québec, QC, Canada, 2013; p. 11. [Google Scholar]
- Whaley, D.K.; Sowers, K.; Roe, D. Cabbage Seedpod Weevil Management in Canola; Washington State University Extension: Pullman, WA, USA, 2016; pp. 1–8. [Google Scholar]
- Baxter-Gilbert, J.H.; Riley, J.L.; Neufeld, C.J.; Litzgus, J.D.; Lesbarrères, D. Road mortality potentially responsible for billions of pollinating insect deaths annually. J. Insect Conserv. 2015, 19, 1029–1035. [Google Scholar] [CrossRef]
- Martin, A.E.; Graham, S.L.; Henry, M.; Pervin, E.; Fahrig, L. Flying insect abundance declines with increasing road traffic. Insect Conserv. Divers. 2018, 11, 608–613. [Google Scholar] [CrossRef]
- Andersson, P.; Koffman, A.; Sjödin, N.E.; Johansson, V. Roads may act as barriers to flying insects: Species composition of bees and wasps differs on two sides of a large highway. Nat. Conserv. 2017, 18, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Steidle, J.L.; Kimmich, T.; Csader, M.; Betz, O. Negative impact of roadside mowing on arthropod fauna and its reduction with ‘arthropod-friendly’ mowing technique. J. Appl. Entomol. 2022, 146, 465–472. [Google Scholar] [CrossRef]
- Morandin, L.A.; Long, R.F.; Kremen, C. Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2014, 189, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Wratten, S.D.; Bowie, M.H.; Hickman, J.M.; Evans, A.M.; Sedcole, J.R.; Tylianakis, J.M. Field boundaries as barriers to movement of hover flies (Diptera: Syrphidae) in cultivated land. Oecologia 2003, 134, 605–611. [Google Scholar] [CrossRef]
- Bianchi, F.J.; Wäckers, F.L. Effects of flower attractiveness and nectar availability in field margins on biological control by parasitoids. Biol. Control 2008, 46, 400–408. [Google Scholar] [CrossRef]
- Winkler, K.; Wäckers, F.L.; Termorshuizen, A.J.; van Lenteren, J.C. Assessing risks and benefits of floral supplements in conservation biological control. BioControl 2010, 55, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Pollier, A.; Tricault, Y.; Plantegenest, M.; Bischoff, A. Sowing of margin strips rich in floral resources improves herbivore control in adjacent crop fields. Agric. For. Entomol. 2019, 21, 119–129. [Google Scholar] [CrossRef]
- Lee, J.C.; Andow, D.A.; Heimpel, G.E. Influence of floral resources on sugar feeding and nutrient dynamics of a parasitoid in the field. Ecol. Entomol. 2006, 31, 470–480. [Google Scholar] [CrossRef]
- González, E.; Štrobl, M.; Janšta, P.; Hovorka, T.; Kadlec, T.; Knapp, M. Artificial temporary non-crop habitats support parasitoids on arable land. Biol. Conserv. 2022, 265, 109409. [Google Scholar] [CrossRef]
- Pfiffner, L.; Wyss, E. Use of sown wildflower strips to enhance natural enemies of agricultural pests. In Ecological Engineering for Pest Management: Advances in Habitat Manipulation for Arthropods; Gurr, G.M., Wratten, S.D., Altieri, M.A., Eds.; CSIRO Publishing: Collingwood, Australia, 2004; pp. 165–186. [Google Scholar]
- Lavoie, C.; Saint-Louis, A.; Guay, G.; Groeneveld, E. Les plantes vasculaires exotiques naturalisées: Une nouvelle liste pour le Québec. Nat. Can. 2012, 136, 6–32. [Google Scholar] [CrossRef] [Green Version]
- Hatt, S.; Uyttenbroeck, R.; Lopes, T.; Chen, J.L.; Piqueray, J.; Monty, A.; Francis, F. Effect of flower traits and hosts on the abundance of parasitoids in perennial multiple species wildflower strips sown within oilseed rape (Brassica napus) crops. Arthropod-Plant Interact. 2018, 12, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Renault, D.; Vernon, P.; Vannier, G. Critical thermal maximum and body water loss in first instar larvae of three Cetoniidae species (Coleoptera). J. Therm. Biol. 2005, 30, 611–617. [Google Scholar] [CrossRef]
- Tee, H.S.; Lee, C.Y. Water balance profiles, humidity preference and survival of two sympatric cockroach egg parasitoids Evania appendigaster and Aprostocetus hagenowii (Hymenoptera: Evaniidae; Eulophidae). J. Insect Physiol. 2015, 77, 45–54. [Google Scholar] [CrossRef]
- de Pedro, L.; Beitia, F.; Sabater-Muñoz, B.; Harbi, A.; Ferrara, F.; Polidori, C.; Asís, J.D.; Tormos, J. Parasitism of Aganaspis daci against Ceratitis capitata under Mediterranean climate conditions. Entomol. Exp. Appl. 2017, 163, 287–295. [Google Scholar] [CrossRef]
- Yokoyama, V.Y.; Rendón, P.A.; Sivinski, J. Psyttalia cf. concolor (Hymenoptera: Braconidae) for Biological Control of Olive Fruit Fly (Diptera: Tephritidae) in California. Environ. Entomol. 2008, 37, 764–773. [Google Scholar] [CrossRef] [Green Version]
- Moezipour, M.; Kafil, M.; Allahyari, H. Functional response of Trichogramma brassicae at different temperatures and relative humidities. Bull. Insectol. 2008, 61, 245–250. [Google Scholar]
- Bruce, A.Y.; Schulthess, F.; Mueke, J. Host acceptance, suitability, and effects of host deprivation on the West African egg parasitoid Telenomus isis (Hymenoptera: Scelionidae) reared on East African stemborers under varying temperature and relative humidity regimens. Environ. Entomol. 2009, 38, 904–919. [Google Scholar] [CrossRef] [PubMed]
- Duale, A.H. Effect of temperature and relative humidity on the biology of the stem borer parasitoid Pediobius furvus (Gahan) (Hymenoptera: Eulophidae) for the management of stem borers. Environ. Entomol. 2005, 34, 1–5. [Google Scholar] [CrossRef]
- Mainali, B.P.; Kim, S.; Lim, U.T. Interactive influence of temperature and relative humidity on egg parasitoids of Riptortus pedestris (Hemiptera: Alydidae). J. Econ. Entomol. 2012, 105, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Veres, A.; Petit, S.; Conord, C.; Lavigne, C. Does landscape composition affect pest abundance and their control by natural enemies? A review. Agric. Ecosyst. Environ. 2013, 166, 110–117. [Google Scholar] [CrossRef]
- Steckel, J.; Westphal, C.; Peters, M.K.; Bellach, M.; Rothenwoehrer, C.; Erasmi, S.; Scherber, C.; Tscharntke, T.; Steffan-Dewenter, I. Landscape composition and configuration differently affect trap-nesting bees, wasps and their antagonists. Biol. Conserv. 2014, 172, 56–64. [Google Scholar] [CrossRef]
- Haro-Barchin, E.; Scheper, J.; Ganuza, C.; De Groot, G.A.; Colombari, F.; van Kats, R.; Kleijn, D. Landscape-scale forest cover increases the abundance of Drosophila suzukii and parasitoid wasps. Basic Appl. Ecol. 2018, 31, 33–43. [Google Scholar] [CrossRef]
- Lu, Z.X.; Zhu, P.Y.; Gurr, G.M.; Zheng, X.S.; Read, D.M.; Heong, K.L.; Yang, Y.J.; Xu, H.X. Mechanisms for flowering plants to benefit arthropod natural enemies of insect pests: Prospects for enhanced use in agriculture. Insect Sci. 2014, 21, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pellissier, M.E.; Jabbour, R. Herbivore and parasitoid insects respond differently to annual and perennial floral strips in an alfalfa ecosystem. Biol. Control 2018, 123, 28–35. [Google Scholar] [CrossRef]
- Dieckhoff, C.; Theobald, J.C.; Wäckers, F.L.; Heimpel, G.E. Egg load dynamics and the risk of egg and time limitation experienced by an aphid parasitoid in the field. Ecol. Evol. 2014, 4, 1739–1750. [Google Scholar] [CrossRef]
- Maisonhaute, J.É.; Labrie, G.; Lucas, E. Population dynamics of the soybean aphid (Hemiptera: Aphididae) in Quebec (Canada). J. Econ. Entomol. 2016, 109, 1465–1468. [Google Scholar] [CrossRef]
- van Neerbos, F.A.; de Boer, J.G.; Salis, L.; Tollenaar, W.; Kos, M.; Vet, L.E.; Harvey, J.A. Honeydew composition and its effect on life-history parameters of hyperparasitoids. Ecol. Entomol. 2020, 45, 278–289. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landscape Predictor | Details | Landscape Predictor | Details |
|---|---|---|---|
| Area of each crop group (m2) (8) | Canola, cereals, corn, soybean, hay/pastures, other crops, mixed crops, and undifferentiated crops | Total landscape Shannon index (landscape diversity) (1) | 1 |
| Area of each semi-natural habitat (m2) (4) | Forests, aquatic areas (lakes, ponds, rivers…), shrublands, bogs | Crop Shannon index (crop diversity) (1) | 1 |
| Main roads and urban areas (m2) (2) | Urban areas: group of buildings, shops, golf | Number of semi-natural habitats patches, number of crop patches and total number of patches (3) | Distributed across all the landscape |
| Perimeter (m), area (m2) and perimeter-to-area ratio of sampled canola fields (3) | Related to size and field border density of sampled canola field | Distance between the sampled canola field and the nearest forest (m) (1) | Measured from the center of the sampled canola field |
| Average perimeter-to-area ratio of all crops (2) | Field border density index | Distance between the sampled canola field and the nearest aquatic area (m) (1) | Measured from the center of the sampled canola field |
| Landscape richness (1) | Number of different elements | Hedgerows length (m) (1) | Lines of trees/shrubs across all the landscape |
| Total crop area (m2) (1) | Eight crop groups considered altogether | Total semi-natural area (m2) (1) | Four semi-natural habitats considered altogether |
| Year | Canola Fields Number | CSW Abundance (Mean per 10 Sweeps) (±SE) | CSW Infestation Rate (Mean in %) (±SE) | Parasitoids Number (Mean per 1000 Pods) (±SE) | T. perfectus Number (Mean per 1000 Pods) (±SE) | Overall Parasitism Rate (Mean in %) (±SE) | T. perfectus Parasitism Rate (Mean in %) (±SE) | % of Canola Fields above Damage Threshold |
|---|---|---|---|---|---|---|---|---|
| 2015 | 21 | 1.80 (±0.37) | 0.32 (±0.13) | 2.29 (±0.93) | 1.71 (±0.78) | 48.39 (±11.54) 2 | 24.76 (±9.10) 2 | 4.76 |
| 2016 | 19 | 3.63 (±1.59) | 3.45 (±1.48) | 6.40 (±2.53) | 4.00 (±1.71) | 12.88 (±4.00) | 5.40 (±2.01) | 21.05 |
| 2017 | 20 | 7.15 (±2.88) | 3.69 (±1.34) | 3.33 (±1.36) | 2.78 (±1.17) | 9.81 (±2.54) | 6.33 (±1.60) | 35.00 |
| 2018 | 26 | 1.09 (±0.37) | 0.86 (±0.26) | 4.95 (±2.07) | 3.53 (±1.64) | 23.09 (±7.27) | 14.93 (±5.32) | 7.69 |
| 2019 | 28 1 | 9.24 (±3.37) | 6.91 (±2.15) | 6.44 (±2.06) | 4.88 (±1.52) | 9.88 (±3.21) | 6.10 (±2.37) | 39.29 |
| 2020 | 26 1 | 2.83 (±0.75) | 6.43 (±1.66) | 14.00 (±3.23) | 10.72 (±2.52) | 26.82 (±4.53) | 21.76 (±3.95) | 30.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Ottavio, M.; Boquel, S.; Labrie, G.; Lucas, E. Landscape Effects on the Cabbage Seedpod Weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), and on Its Parasitoid, Trichomalus perfectus (Hymenoptera: Pteromalidae), in Canola. Insects 2023, 14, 327. https://doi.org/10.3390/insects14040327
D’Ottavio M, Boquel S, Labrie G, Lucas E. Landscape Effects on the Cabbage Seedpod Weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), and on Its Parasitoid, Trichomalus perfectus (Hymenoptera: Pteromalidae), in Canola. Insects. 2023; 14(4):327. https://doi.org/10.3390/insects14040327
Chicago/Turabian StyleD’Ottavio, Marie, Sébastien Boquel, Geneviève Labrie, and Eric Lucas. 2023. "Landscape Effects on the Cabbage Seedpod Weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), and on Its Parasitoid, Trichomalus perfectus (Hymenoptera: Pteromalidae), in Canola" Insects 14, no. 4: 327. https://doi.org/10.3390/insects14040327
APA StyleD’Ottavio, M., Boquel, S., Labrie, G., & Lucas, E. (2023). Landscape Effects on the Cabbage Seedpod Weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), and on Its Parasitoid, Trichomalus perfectus (Hymenoptera: Pteromalidae), in Canola. Insects, 14(4), 327. https://doi.org/10.3390/insects14040327