
A Novel Model of Pathogenesis of Metarhizium anisopliae Propagules through the Midguts of Aedes aegypti Larvae


 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture and Preparation of M. anisopliae Conidia
2.2. Influence of Shaker Rotations and the Time of Incubation on Blastospore Production
2.3. Insects
2.4. Survival Analyses
2.5. Scanning Electron Microscopy and Histopathological Analysis
2.6. Statistical Analyses
3. Results
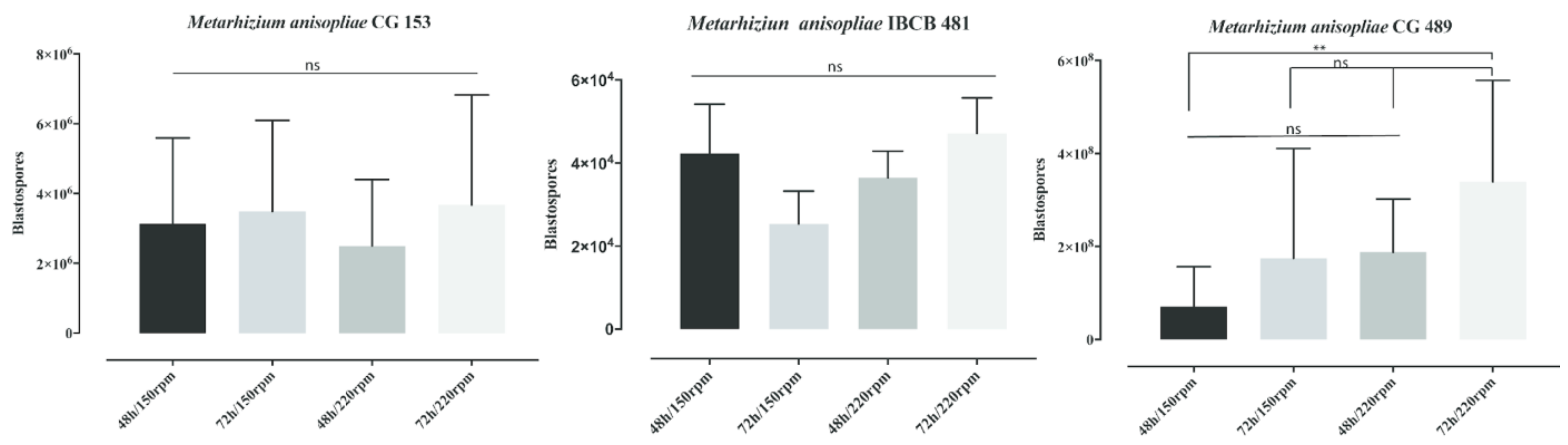
3.1. Influence of Shaker Rotations and the Time of Incubation on Blastospore Production
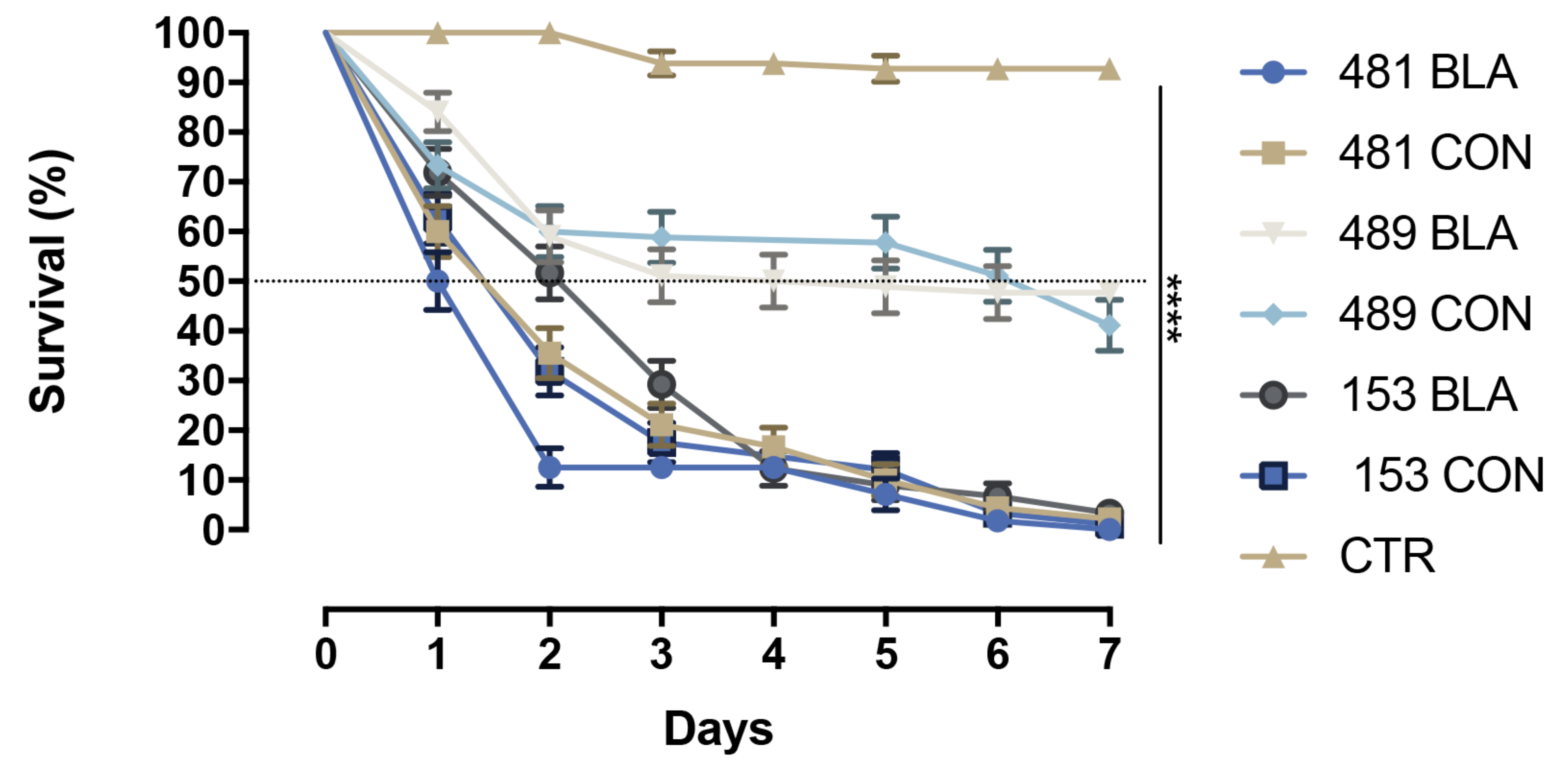
3.2. Survival Analyzes
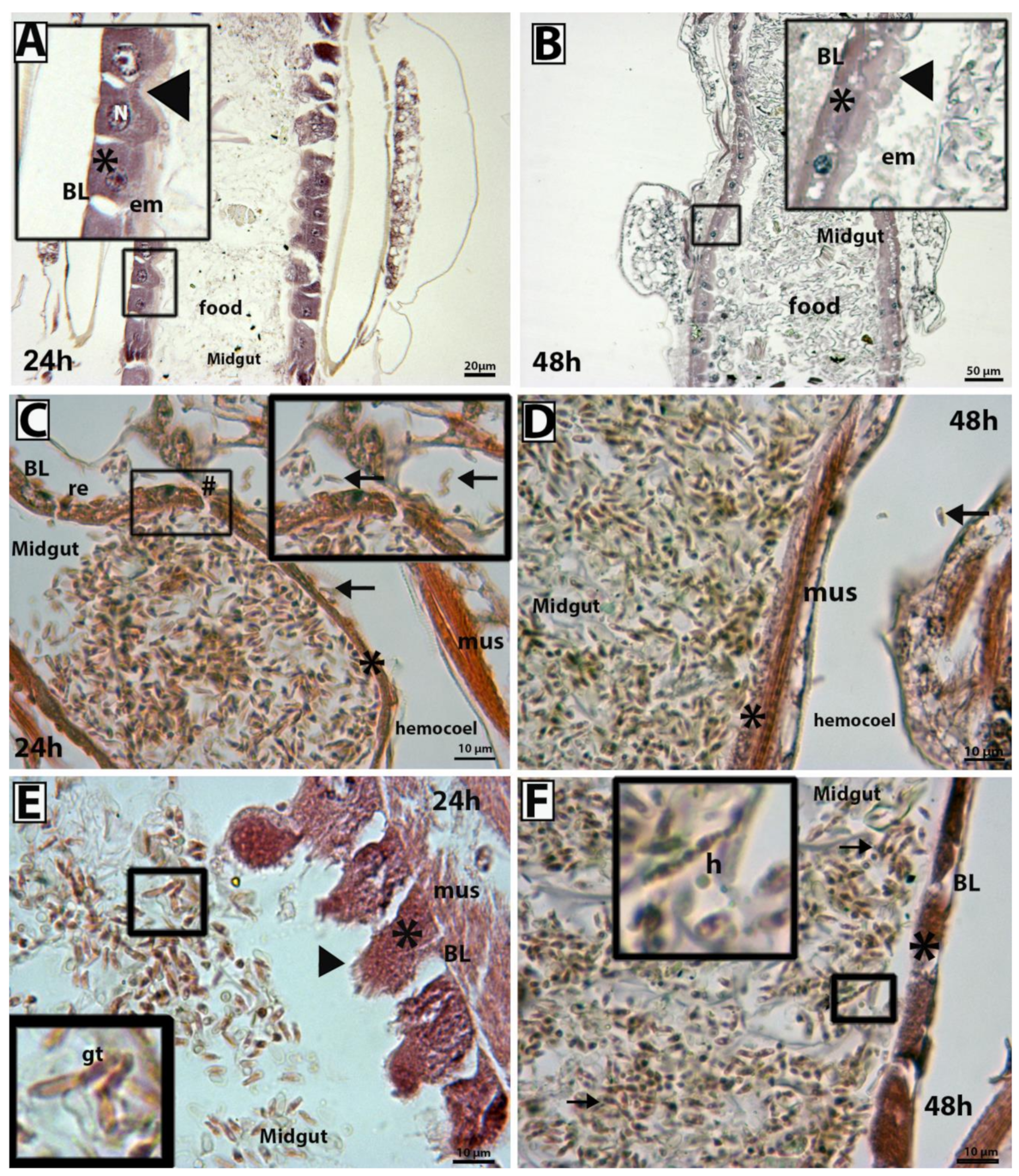
3.3. Scanning Electron Microscopy and Histopathological Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schrank, A.; Vainstein, M.H. Metarhizium anisopliae enzymes and toxins. Toxicon 2010, 56, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Alkhaibari, A.M.; Wood, M.J.; Yavasoglu, S.I.; Bull, J.C.; Butt, T.M. Optimizing the application timing and dosage of Metarhizium brunneum (Hypocreales: Clavicipitaceae) as a biological control agent of Aedes aegypti (Diptera: Culicidae) larvae. J. Med. Entomol. 2022, 20, tjac186. [Google Scholar] [CrossRef]
- De Oliveira Barbosa Bitencourt, R.; Mallet, J.R.S.; Mesquita, E.; Gôlo, P.S.; Fiorotti, J.; Bittencourt, V.R.E.P.; Pontes, E.G.; Angelo, I.C. Larvicidal activity, route of interaction and ultrastructural changes in Aedes aegypti exposed to entomopathogenic fungi. Acta Trop. 2021, 213, 1–11. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Barbosa Bitencourt, R.; de Souza Faria, F.; Marchesini, P.; Reis Dos Santos-Mallet, J.; Guedes Camargo, M.; Rita Elias Pinheiro Bittencourt, V.; Guedes Pontes, E.; Baptista Pereira, D.; Siqueira de Almeida Chaves, D.; da Costa Angelo, I. Entomopathogenic fungi and Schinus molle essential oil: The combination of two eco-friendly agents against Aedes aegypti larvae. J. Invertebr. Pathol. 2022, 194, 107827. [Google Scholar] [CrossRef] [PubMed]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Thomas, M.B.; Read, A.F. Fungal bioinsecticide with a sting. Nat. Biotechnol. 2007, 25, 1367–1368. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Carolino, A.T.; Yavasoglu, S.I.; Maffeis, T.; Mattoso, T.C.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Metarhizium brunneum blastospore pathogenesis in Aedes aegypti larvae: Attack on several fronts accelerates mortality. PLoS Pathog. 2016, 12, e1005715. [Google Scholar] [CrossRef] [Green Version]
- Alkhaibari, A.M.; Carolino, A.T.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Differential pathogenicity of Metarhizium blastospores and conidia against larvae of three mosquito species. J. Med. Entomol. 2017, 54, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Alkhaibari, A.M.; Lord, A.M.; Maffeis, T.; Bull, J.C.; Olivares, F.L.; Samuels, R.I.; Butt, T.M. Highly specific host-pathogen interactions influence Metarhizium brunneum blastospore virulence against Culex quinquefasciatus larvae. Virulence 2018, 9, 1449–1467. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Barbosa Bitencourt, R.; Farias, F.S.; Freitas, M.C.; Balduino, C.J.R.; Mesquita, E.S.; Corval, A.R.C.; Corval, P.S.; Pontes, E.G.; Bittencourt, V.R.E.P.; Angelo, I.C. In vitro control of Aedes aegypti larvae using Beauveria bassiana. Int. J. Bioeng. Life Sci. 2018, 12, 400–404. [Google Scholar] [CrossRef]
- Carolino, A.T.; Gomes, S.A.; Pontes Teodoro, T.B.P.; Mattoso, T.C.; Samuels, R.I. Aedes aegypti pupae are highly susceptible to infection by Metarhizium anisopliae blastospores. J. Pure Appl. Microbiol. 2019, 13, 1629–1634. [Google Scholar] [CrossRef] [Green Version]
- De Paula, A.R.; Silva, L.E.I.; Ribeiro, A.; da Silva, G.A.; Silva, C.P.; Butt, T.M.; Samuels, R.I. Metarhizium anisopliae blastospores are highly virulent to adult Aedes aegypti, an important arbovirus vector. Parasit. Vectors. 2021, 14, 2–10. [Google Scholar] [CrossRef]
- Noskov, Y.A.; Polenogova, O.V.; Yaroslavtseva, O.N.; Belevich, O.E.; Yurchenko, Y.A.; Chertkova, E.A.; Kryukova, N.A.; Kryukov, V.Y.; Glupov, V.V. Combined effect of the entomopathogenic fungus Metarhizium robertsii and avermectins on the survival and immune response of Aedes aegypti larvae. PeerJ 2019, 7, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Gomes, S.A.; Paula, A.R.; Ribeiro, A.; Moraes, C.O.; Santos, J.W.; Silva, C.P.; Samuels, R.I. Neem oil increases the efficiency of the entomopathogenic fungus Metarhizium anisopliae for the control of Aedes aegypti (Diptera: Culicidae) larvae. Parasit. Vectors 2015, 8, 669. [Google Scholar] [CrossRef] [Green Version]
- Butt, T.M.; Greenfield, B.P.J.; Greig, C.; Maffeis, T.G.G.; Taylor, J.W.D.; Piasecka, J.; Dudley, E.; Abdulla, A.; Dubovskiy, I.M.; Garrido-Jurado, I.; et al. Metarhizium anisopliae pathogenesis of mosquito larvae: A verdict of accidental death. PLoS ONE 2013, 8, e0081686. [Google Scholar] [CrossRef] [Green Version]
- Kalha, C.S.; Singh, P.P.; Kang, S.S.; Hunjan, M.S.; Gupta, V.; Sharma, R. Chapter 12 - Entomopathogenic viruses and bacteria for insect-pest control. J. Integr. Pest Manag. 2014, 1, 225–244. [Google Scholar] [CrossRef]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-fungal-interactions: A detailed review on entomopathogenic fungi pathogenicity to combat insect pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-pathogen interactions, advances in genetics. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [CrossRef]
- De Oliveira Barbosa Bitencourt, R.; Salcedo-Porras, N.; Umanã-Diaz, C.; Angelo, I.C.; Lowenberger, C. Antifungal immune responses in mosquitoes (Diptera: Culicidae): A review. J. Invertebr. Pathol. 2021, 178, 107505. [Google Scholar] [CrossRef] [PubMed]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? Insights into alternative modes of action of entomopathogenic fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran, K.; Akutse, K.S.; Sivaramakrishnan, S.; Wang, L. Determination and characterization of destruxin production in Metarhizium anisopliae Tk6 and formulations for Aedes aegypti mosquitoes control at the field level. Toxicon 2016, 15, 89–96. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef]
- Lacey, C.M.; Lacey, L.A.; Roberts, D.R. Route of invasion and histopathology of Metarhizium anisopliae in Culex quinquefasciatus. J. Invertebr. Pathol. 1988, 52, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, B.P.; Lord, A.M.; Dudley, E.; Butt, T.M. Conidia of the insect pathogenic fungus, Metarhizium anisopliae, fail to adhere to mosquito larval cuticle. R. Soc. Open Sci. 2014, 1, 140193. [Google Scholar] [CrossRef] [Green Version]
- Da Cunha, L.P.; Casciatori, F.P.; de Cenço Lopes, I.; Thoméo, J.C. Production of conidia of the entomopathogenic fungus Metarhizium anisopliae ICB 425 in a tray bioreactor. Bioprocess. Biosyst. Eng. 2019, 42, 1757–1768. [Google Scholar] [CrossRef]
- Kruger, R.D.; Posadas, J.B.; Lewylle, M.A.; Mini, J.I.; Lecuona, R.E. Solid substrate production and formulation of an isolate of Metarhizium anisopliae for biological control of stem bug Tibraca limbativentrisa. World Appl. Sci. J. 2014, 32, 1242–1251. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jackson, M.A.; Kobori, N.N.; Behle, R.W.; Dunlap, C.A.; Júnior, I.D. Glucose concentration alters dissolved oxygen levels in liquid cultures of Beauveria bassiana and affects formation and bioefficacy of blastospores. Appl. Microbiol. Biotechnol. 2015, 99, 6653–6665. [Google Scholar] [CrossRef]
- Holder, D.J.; Kirkland, B.H.; Lewis, M.W.; Keyhani, N.O. Surface characteristics of the entomopathogenic fungus Beauveria (Cordyceps) bassiana. Microbiology 2007, 153, 3448–3457. [Google Scholar] [CrossRef] [Green Version]
- Mascarin, G.M.; Jackson, M.A.; Kobori, N.N.; Behle, R.W.; Delalibera Júnior, Í. Liquid culture fermentation for rapid production of desiccation tolerant blastospores of Beauveria bassiana and Isaria fumosorosea strains. J. Invertebr. Pathol. 2015, 127, 11–20. [Google Scholar] [CrossRef]
- Miranpuri, G.S.; Khachatourians, G.G. Larvicidal activity of blastospores and conidiospores of Beauveria bassiana (strain GK 2016) against age groups of Aedes aegypti. Vet. Parasitol. 1990, 37, 155–162. [Google Scholar] [CrossRef]
- Lemos, A.B.; Adam, F.C.; Moura, K.R.S.; Moraes, L.B.; Silva, O.S. Histological and histochemical characterization of the midgut of healthy Aedes aegypti larvae. ARRB 2018, 22, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Alves, S.B. Controle Microbiano de Insetos, 2nd ed.; Alves, S.B., Ed.; FEALQ: Piracicaba, Brazil, 1998; pp. 289–382. [Google Scholar]
- Duarte, G.F. Viabilidade de Massas Ovígeras de Biomphalaria glabrata em filme de Água e Susceptibilidade a Metarhizium anisopliae e Beauveria bassiana. Master’s Thesis, Universidade Federal de Goiás, Goiás, Brazil, 2014. Available online: https://repositorio.bc.ufg.br/tede/handle/tede/4200 (accessed on 10 October 2022).
- Bernardo, C.C.; Barreto, L.P.; E Silva, C.S.R.; Luz, C.; Arruda, W.; Fernandes, É.K.K. Conidia and blastospores of Metarhizium spp. and Beauveria bassiana s.l.: Their development during the infection process and virulence against the tick Rhipicephalus microplus. Ticks Tick Borne Dis. 2018, 9, 1334–1342. [Google Scholar] [CrossRef]
- Gregory, G.E. Alcoholic Bouin fixation of insect nervous systems for bodian silver staining. III. A shortened, single impregnation method. Stain Technol. 1980, 55, 161–165. [Google Scholar] [CrossRef]
- Latifian, M.; Rad, B. Effects of photoperiod, incubation time and shaker incubation rotation speed on the production of Beauveria bassiana and Metarhizium anisopliae in liquid fermentation. Biocontrol Plant Prot. 2020, 7, 139–152. [Google Scholar] [CrossRef]
- Kleespies, R.G.; Zimmermann, G. Production of blastospores by three strains of Metarhizium anisopliae (metch.) sorokin in submerged culture. Biocontrol Sci. Technol. 1992, 2, 127–135. [Google Scholar] [CrossRef]
- Issaly, N.; Chauveau, H.; Aglevor, F.; Fargues, J.; Durand, A. Influence of nutrient, pH and dissolved oxygen on the production of Metarhizium flavoviride Mf189 blastospores in submerged batch culture. Process. Biochem. 2005, 40, 1425–1431. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Mascarin, G.M. Mass Production of Fungal Entomopathogens, in Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 141–155. [Google Scholar] [CrossRef]
- Miranpuri, G.S.; Khachatourians, G.G. Infection sites of the entomopathogenic fungus Beauveria bassiana in the larvae of the mosquito Aedes aegypti. Entomo. Exp. Appl. 1991, 59, 19–27. [Google Scholar] [CrossRef]
- Silva, C.; Pinheiro, N.L.; Scherer, P.O.; Falcão, S.S.; Ribeiro, V.R.; Mendes, R.M.M.; Chaves, R.; Cardozo-De-Almeida, M.; Dos Santos-Mallet, J.R. Histology and ultrastructure of Aedes albopictus larval midgut infected with Bacillus thuringiensis var. israelensis. Microsc. Res. Tech. 2008, 71, 663–668. [Google Scholar] [CrossRef]
- Jiraungkoorskul, W. Efficiency of Tinospora crispa against Culex quinquefasciatus larva. Environ. Sci. Pollut. Res. 2019, 26, 14712–14716. [Google Scholar] [CrossRef] [PubMed]
- Al-mehmadi, R.M.; Al-khalaf, A.A. Larvicidal and histological effects of Melia azedarach extract on Culex quinquefasciatus say larvae (Diptera: Culicidae). J. King Saud Univ. Sci. 2010, 22, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.R.; Raikhel, A.S.; Lea, A.O. Ultrastructure of midgut endocrine cells in the adult mosquito, Aedes aegypti. Tissue Cell. 1985, 17, 709–721. [Google Scholar] [CrossRef]
- Fernandes, K.M.; Neves, C.A.; Serrão, J.E.; Martins, G.F. Aedes aegypti midgut remodeling during metamorphosis. Parasitol. Int. 2014, 63, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Taracena, M.L.; Bottino-Rojas, V.; Talyuli, O.A.C.; Walter-Nuno, A.B.; Oliveira, J.H.M.; Angleró-Rodriguez, Y.I.; Wells, M.B.; Dimopoulos, G.; Oliveira, P.L.; Paiva-Silva, G.O. Regulation of midgut cell proliferation impacts Aedes aegypti susceptibility to dengue virus. PLoS Negl. Trop. Dis. 2018, 12, e0006498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hixson, B.; Taracena, M.L.; Buchon, N. Midgut epithelial dynamics are central to mosquitoes’ physiology and fitness, and to the transmission of vector-borne disease. Front. Cell Infect. Microbiol. 2021, 11, 653156. [Google Scholar] [CrossRef]
- Buchon, N.; Broderick, N.A.; Chakrabarti, S.; Lemaitre, B. Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes. Dev. 2009, 23, 2333–2344. [Google Scholar] [CrossRef] [Green Version]
- Buchon, N.; Broderick, N.A.; Poidevin, M.; Pradervand, S.; Lemaitre, B. Drosophila intestinal response to bacterial infection: Activation of host defense and stem cell proliferation. Cell Host Microbe 2009, 5, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Dinglasan, R.R.; Devenport, M.; Florens, L.; Johnson, J.R.; McHugh, C.A.; Donnelly-Doman, M.; Carucci, D.J.; Yates, J.R.; Jacobs-Lorena, M. The Anopheles gambiae adult midgut peritrophic matrix proteome. Insect. Biochem. Mol. Biol. 2009, 39, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, R.G.; Kang, S.; Simões, M.L.; Anglero-Rodríguez, Y.I.; Dimopoulos, G. Mosquito gut antiparasitic and antiviral immunity. Dev. Comp. Immunol. 2016, 64, 53–64. [Google Scholar] [CrossRef]
- Pedrini, N. The entomopathogenic fungus Beauveria bassiana shows its toxic side within insects: Expression of genes encoding secondary metabolites during pathogenesis. J. Fungi 2022, 8, 488. [Google Scholar] [CrossRef]
- Samuels, R.I.; Reynolds, S.E.; Charnley, A.K. Calcium channel activation of insect muscle by destruxins, insecticidal compounds produced by the entomopathogenic fungus, Metarhizium anisopliae. Comp. Biochem. Physiol. 1988, 90, 403–412. [Google Scholar] [CrossRef]
- Dumas, C.; Ravallec, M.; Matha, V.; Vey, A. Comparative study of the cytological aspects of the mode of action of destruxins and other peptidic fungal metabolites on target epithelial cells. J. Invert. Pathol. 1996, 67, 137–146. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | 1 × 107 Propagules mL−1 | S50 | ||||||
|---|---|---|---|---|---|---|---|---|
| CTR | 481 BLA | 481 CON | 489 BLA | 489 CON | 153 BLA | 153 CON | ||
| CTR | - | χ2 = 192.3; p < 0.0001 | χ2 = 243.3; p < 0.0001 | χ2 = 48.10; p < 0.0001 | χ2 = 61.35; p < 0.0001 | χ2 = 178.1; p < 0.0001 | χ2 = 197.0; p < 0.0001 | ND |
| 481 BLA | - | - | χ2 = 5.461; p = 0.0195 | χ2 = 61.27; p < 0.0001 | χ2 = 53.26; p < 0.0001 | χ2 = 14.89; p = 0.0001 | χ2 = 5.900; p = 0.0151 | 1 |
| 481 CON | - | - | - | χ2 = 47.50; p < 0.0001 | χ2 = 46.53; p < 0.0001 | χ2 = 46.53; p < 0.0001 | χ2 = 1.574; p = 0.2096 | 2 |
| 489 BLA | - | - | - | - | χ2 = 0.887; p = 0.3463 | χ2 = 34.24; p < 0.0001 | χ2 = 50.33; p < 0.0001 | 5.5 |
| 489 CON | - | - | - | - | - | χ2 = 35.43; p < 0.0001 | χ2 = 48.25; p < 0.0001 | 7 |
| 153 BLA | - | - | - | - | - | - | χ2 = 2.264; p = 0.1324 | 3 |
| 153 CON | - | - | - | - | - | - | - | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bitencourt, R.d.O.B.; Santos-Mallet, J.R.d.; Lowenberger, C.; Ventura, A.; Gôlo, P.S.; Bittencourt, V.R.E.P.; Angelo, I.d.C. A Novel Model of Pathogenesis of Metarhizium anisopliae Propagules through the Midguts of Aedes aegypti Larvae. Insects 2023, 14, 328. https://doi.org/10.3390/insects14040328
Bitencourt RdOB, Santos-Mallet JRd, Lowenberger C, Ventura A, Gôlo PS, Bittencourt VREP, Angelo IdC. A Novel Model of Pathogenesis of Metarhizium anisopliae Propagules through the Midguts of Aedes aegypti Larvae. Insects. 2023; 14(4):328. https://doi.org/10.3390/insects14040328
Chicago/Turabian StyleBitencourt, Ricardo de Oliveira Barbosa, Jacenir Reis dos Santos-Mallet, Carl Lowenberger, Adriana Ventura, Patrícia Silva Gôlo, Vânia Rita Elias Pinheiro Bittencourt, and Isabele da Costa Angelo. 2023. "A Novel Model of Pathogenesis of Metarhizium anisopliae Propagules through the Midguts of Aedes aegypti Larvae" Insects 14, no. 4: 328. https://doi.org/10.3390/insects14040328
APA StyleBitencourt, R. d. O. B., Santos-Mallet, J. R. d., Lowenberger, C., Ventura, A., Gôlo, P. S., Bittencourt, V. R. E. P., & Angelo, I. d. C. (2023). A Novel Model of Pathogenesis of Metarhizium anisopliae Propagules through the Midguts of Aedes aegypti Larvae. Insects, 14(4), 328. https://doi.org/10.3390/insects14040328