
Life Table Study of Fall Armyworm (Spodoptera frugiperda) (Lepidoptera: Noctuidae) on Three Host Plants under Laboratory Conditions



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fall Armyworm Colony
2.2. Host Plants
2.3. Life Table Study
2.4. Life Table Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Developmental Duration, APOP, TPOP, Oviposition, Adult Longevity, and Fecundity of FAW
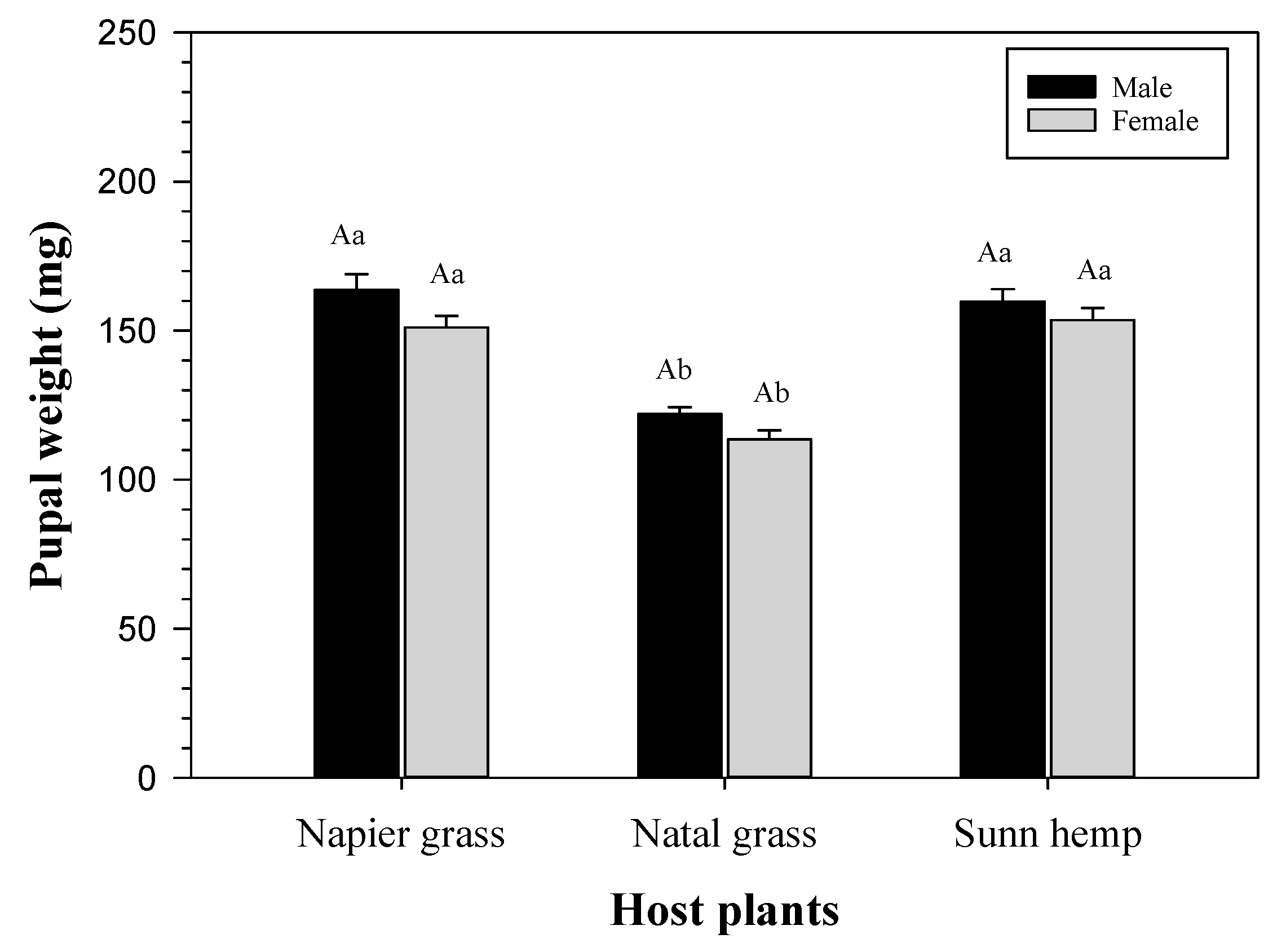
3.2. Pupal Weight of FAW
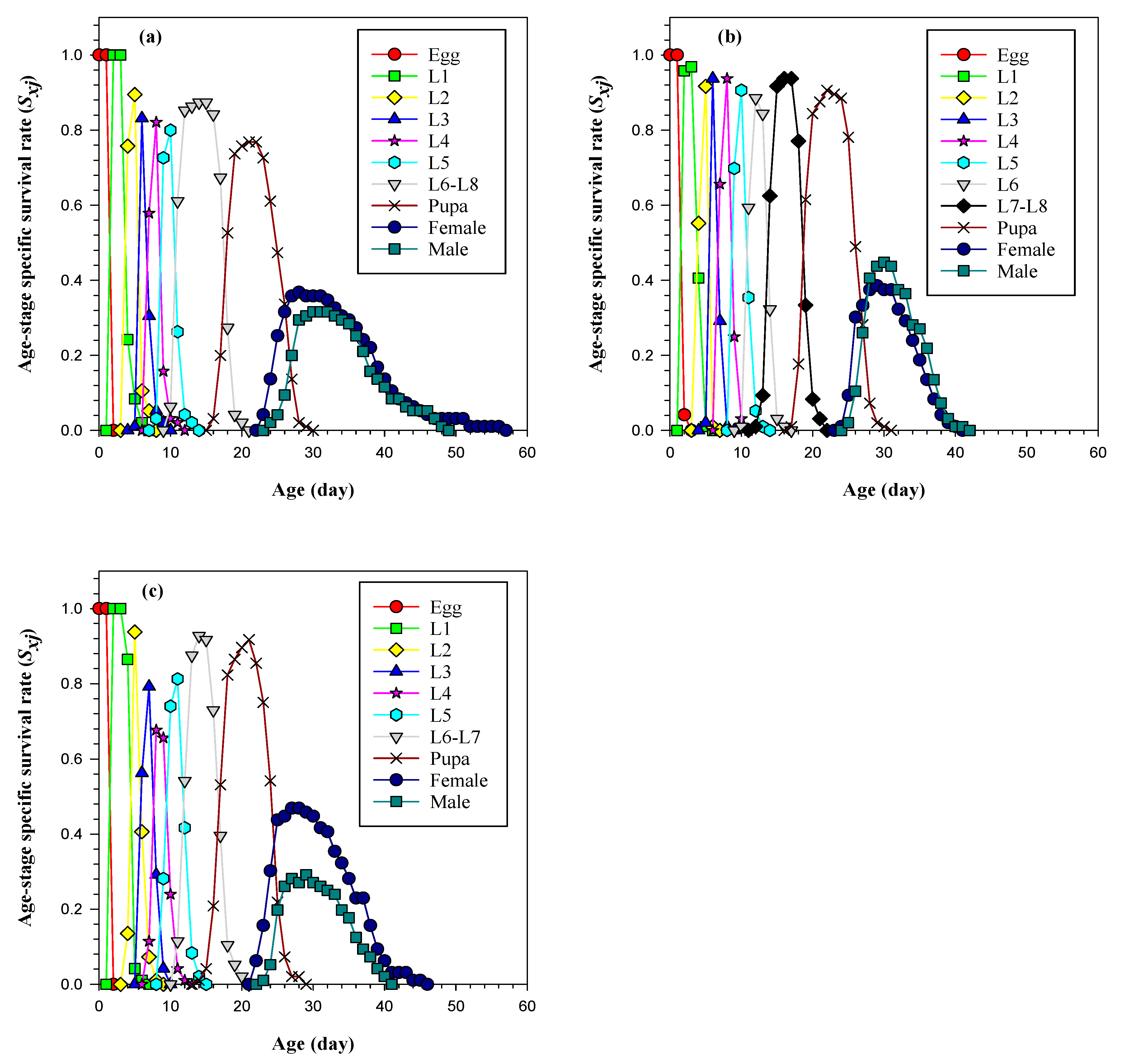
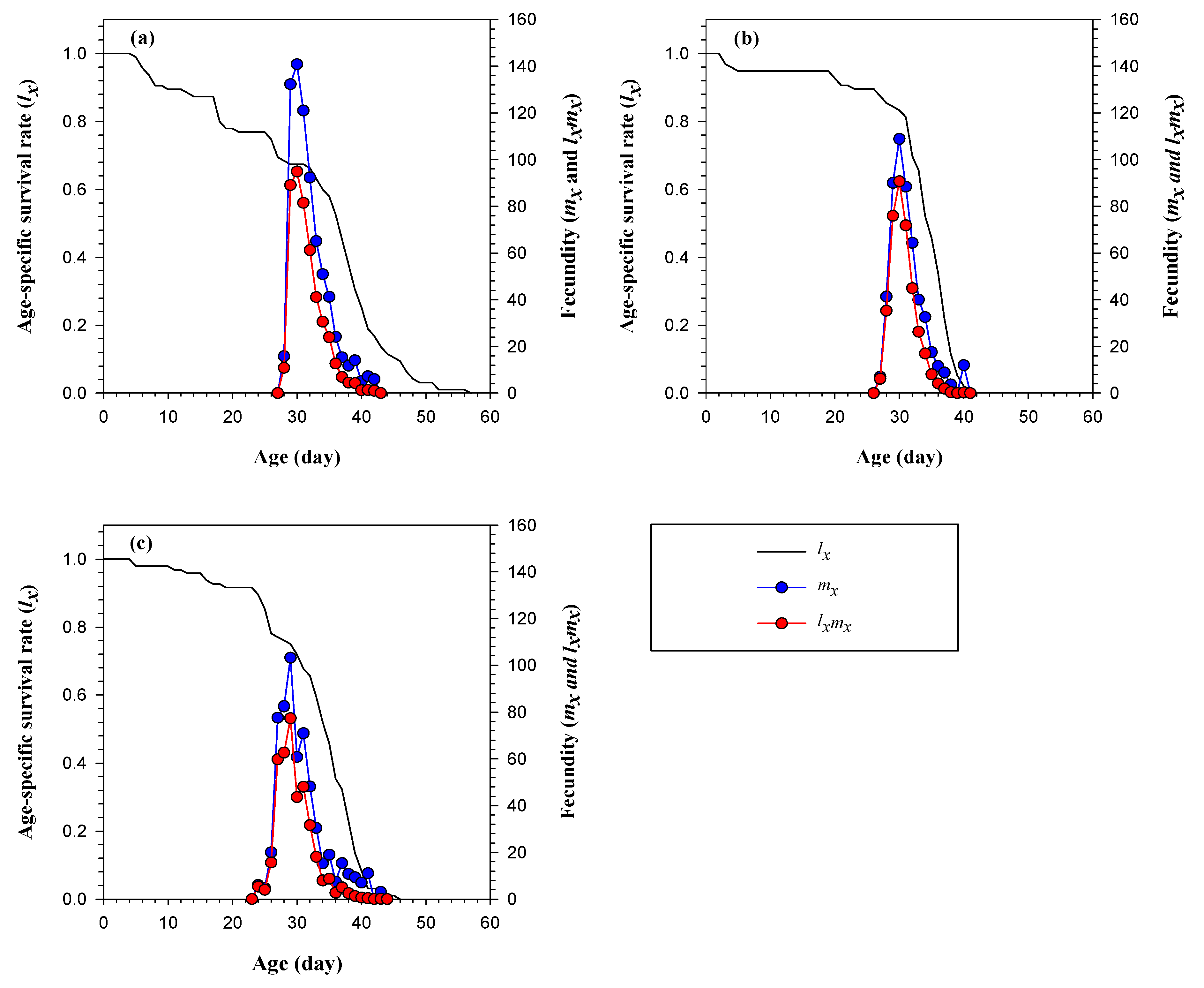
3.3. Life Table and Population Parameters of FAW
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sagar, G.; Aastha, B.; Laxman, K. An introduction of fall armyworm (Spodoptera frugiperda) with management strategies: A review paper. Nippon. J. Environ. Sci. 2020, 1, 1010. [Google Scholar] [CrossRef]
- Maruthadurai, R.; Ramesh, R. Occurrence, damage pattern and biology of fall armyworm, Spodoptera frugiperda (JE smith)(Lepidoptera: Noctuidae) on fodder crops and green amaranth in Goa, India. Phytoparasitica 2020, 48, 15–23. [Google Scholar] [CrossRef]
- FAO. Preparing Countries to Keep Fall Armyworm Away from Their Territories. Available online: https://www.ippc.int/en/news/preparing-countries-to-keep-fall-armyworm-away-from-their-territories/ (accessed on 13 October 2021).
- Khatri, S.; Pakuwal, P.; Khanal, S. Archives of Agriculture and Environmental Science. Arch. Agric. Environ. Sci. 2020, 5, 583–591. [Google Scholar] [CrossRef]
- FAO. Global Action for Fall Armyworm Control. Available online: https://www.fao.org/fall-armyworm/monitoring-tools/faw-map/en/ (accessed on 13 October 2021).
- Everington, K. Fall Armyworms from China Invade Taiwan. Available online: https://www.taiwannews.com.tw/en/news/3721661 (accessed on 13 May 2021).
- Wang, W.; He, P.; Zhang, Y.; Liu, T.; Jing, X.; Zhang, S. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in China. Insects 2020, 11, 639. [Google Scholar] [CrossRef]
- CABI. CABI-Led Study Provides Comprehensive Review of Devastating Fall Armyworm Pest. Available online: https://www.cabi.org/news-article/cabi-led-study-provides-comprehensive-review-of-devastating-fall-armyworm-pest/ (accessed on 15 May 2021).
- Montezano, D.G.; Sosa-Gómez, D.; Specht, A.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.D.; Peterson, J.A.; Hunt, T. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Rwomushana. Spodoptera frugiperda (Fall Armyworm). Available online: https://www.cabidigitallibrary.org/doi/full/10.1079/cabicompendium.29810 (accessed on 14 September 2022).
- Villegas-Mendoza, J.M.; Rosas-García, N.M. Visual and gustatory responses of Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae to artificial food dyes. Fla. Entomol. 2013, 96, 1102–1106. [Google Scholar] [CrossRef]
- Moraes, T.; da Silva, A.F.; Leite, N.A.; Karam, D.; Mendes, S.M. Survival and development of fall armyworm (Lepidoptera: Noctuidae) in weeds during the off-season. Fla. Entomol. 2020, 103, 288–292. [Google Scholar] [CrossRef]
- Chi, H.; You, M.; Atlihan, R.; Smith, C.L.; Kavousi, A.; Özgökçe, M.S.; Güncan, A.; Tuan, S.-J.; Fu, J.-W.; Xu, Y.-Y. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 103–124. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin 1985, 24, 225–240. [Google Scholar]
- Chi, H. TWOSEX-MSChart: Computer Program for Age Stage, Two-Sex Life Table Analysis. Available online: http://140.120.197.173/ecology/products.htm (accessed on 10 December 2021).
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Tibshirani, R.J.; Efron, B. An introduction to the bootstrap. Monogr. Stat. Appl. Probab. 1993, 57, 1–33. [Google Scholar]
- Huang, Y.B.; Chi, H. Life tables of Bactrocera cucurbitae (Diptera: Tephritidae): With an invalidation of the jackknife technique. J. Appl. Entomol. 2013, 137, 327–339. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chen, D.F.; Yang, M.F.; Liu, J.F. The Effect of Temperatures and Hosts on the Life Cycle of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 211. [Google Scholar] [CrossRef]
- Xie, W.; Zhi, J.; Ye, J.; Zhou, Y.; Li, C.; Liang, Y.; Yue, W.; Li, D.; Zeng, G.; Hu, C. Age-stage, two-sex life table analysis of Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae) reared on maize and kidney bean. Chem. Biol. Technol. Agric. 2021, 8, 44. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, J.; Gao, Z.; He, K.; Bai, S.; Zhang, T.; Wang, Z. Performance of Spodoptera frugiperda (Lepidoptera: Noctuidae) Fed on Six Host Plants: Potential Risks to Mid-high Latitude Crops in China. J. Agric. Sci. 2020, 12, 16–27. [Google Scholar] [CrossRef]
- He, L.-M.; Wu, Q.-L.; Gao, X.-W.; Wu, K.-M. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 2021, 20, 745–754. [Google Scholar] [CrossRef]
- Xue, M.; Pang, Y.-H.; Wang, H.-T.; Li, Q.-L.; Liu, T.-X. Effects of four host plants on biology and food utilization of the cutworm, Spodoptera litura. J. Insect Sci. 2010, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- He, L.-M.; Wang, T.-L.; Chen, Y.-C.; Ge, S.-S.; Wyckhuys, K.A.; Wu, K.-M. Larval diet affects development and reproduction of East Asian strain of the fall armyworm, Spodoptera frugiperda. J. Integr. Agric. 2021, 20, 736–744. [Google Scholar] [CrossRef]
- Chen, G.-M.; Chi, H.; Wang, R.-C.; Wang, Y.-P.; Xu, Y.-Y.; Li, X.-D.; Yin, P.; Zheng, F.-Q. Demography and uncertainty of population growth of Conogethes punctiferalis (Lepidoptera: Crambidae) reared on five host plants with discussion on some life history statistics. J. Econ. Entomol. 2018, 111, 2143–2152. [Google Scholar] [CrossRef]
- Du Plessis, H.; Schlemmer, M.-L.; Van den Berg, J. The effect of temperature on the development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2020, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- Pencoe, N.; Martin, P. Development and reproduction of fall armyworms on several wild grasses. Environ. Entomol. 1981, 10, 999–1002. [Google Scholar] [CrossRef]
- Esperk, T.; Tammaru, T.; Nylin, S. Intraspecific variability in number of larval instars in insects. J. Econ. Entomol. 2007, 100, 627–645. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-H.; Cao, Z.; Long, G.-Y.; Yang, X.-B.; Wei, Z.-Y.; Liao, Y.-J.; Hong, Y.; Hu, C.-X. Fitness of fall armyworm, Spodoptera frugiperda to three solanaceous vegetables. J. Integr. Agric. 2021, 20, 755–763. [Google Scholar] [CrossRef]
- Gebretsadik, K.G.; Liu, Y.; Yin, Y.; Zhao, X.; Li, X.; Chen, F.; Zhang, Y.; Chen, J.; Chen, A. Population Growth of Fall Armyworm, Spodoptera frugiperda Fed on Cereal and Pulse Host Plants Cultivated in Yunnan Province, China. Plants 2023, 12, 950. [Google Scholar] [CrossRef]
- Barcelos, L.M.; Fernandes, F.O.; Lopes, C.; Emygdio, B.M.; Valgas, R.; de Carvalho, I.F.; da Rosa, A. Biology and nutritional indexes of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Saccharine sorghum. J. Agric. Sci. 2019, 11, 126–135. [Google Scholar] [CrossRef]
- Farahani, S.; Talebi, A.A.; Fathipour, Y. Life Table of Spodoptera exigua (Lepidoptera: Noctuidae) on Five Soybean Cultivars. Psyche A J. Entomol. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Acharya, R.; Malekera, M.J.; Dhungana, S.K.; Sharma, S.R.; Lee, K.Y. Impact of Rice and Potato Host Plants Is Higher on the Reproduction than Growth of Corn Strain Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 256. [Google Scholar] [CrossRef]
- Guo, J.-W.; Yang, F.; Li, P.; Liu, X.-D.; Wu, Q.-L.; Hu, G.; Zhai, B.-P. Female bias in an immigratory population of Cnaphalocrocis medinalis moths based on field surveys and laboratory tests. Sci. Rep. 2019, 9, 18388. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Stage | Sex (n) | Napier Grass * | Sex (n) | Natal Grass | Sex (n) | Sunn Hemp |
|---|---|---|---|---|---|---|
| Egg | ♀ (38) | 2.00 ± 0.00 Aa | ♀ (37) | 2.00 ± 0.00 Aa | ♀ (45) | 2.00 ± 0.00 Aa |
| ♂ (31) | 2.00 ± 0.00 Aa | ♂ (43) | 2.00 ± 0.00 Aa | ♂ (29) | 2.00 ± 0.00 Aa | |
| 1st instar | ♀ (38) | 2.24 ± 0.08 Ab | ♀ (37) | 2.38 ± 0.08 Ab | ♀ (45) | 2.84 ± 0.07 Aa |
| ♂ (31) | 2.16 ± 0.06 Ab | ♂ (43) | 2.47 ± 0.08 Ab | ♂ (29) | 2.93 ± 0.04 Aa | |
| 2nd instar | ♀ (38) | 1.79 ± 0.06 Aa | ♀ (37) | 1.59 ± 0.08 Aa | ♀ (45) | 1.53 ± 0.07 Aa |
| ♂ (31) | 1.90 ± 0.07 Aa | ♂ (43) | 1.53 ± 0.07 Ab | ♂ (29) | 1.66 ± 0.12 Aab | |
| 3rd instar | ♀ (38) | 1.39 ± 0.08 Ab | ♀ (37) | 1.32 ± 0.07 Ab | ♀ (45) | 1.76 ± 0.06 Aa |
| ♂ (31) | 1.35 ± 0.08 Aab | ♂ (43) | 1.35 ± 0.07 Ab | ♂ (29) | 1.69 ± 0.08 Aa | |
| 4th instar | ♀ (38) | 1.74 ± 0.07 Aa | ♀ (37) | 1.92 ± 0.05 Aa | ♀ (45) | 1.69 ± 0.07 Aa |
| ♂ (31) | 1.71 ± 0.08 Aa | ♂ (43) | 1.98 ± 0.04 Aa | ♂ (29) | 1.69 ± 0.08 Aa | |
| 5th instar | ♀ (38) | 2.08 ± 0.08 Aa | ♀ (37) | 2.11 ± 0.06 Aa | ♀ (45) | 2.36 ± 0.07 Aa |
| ♂ (31) | 2.13 ± 0.07 Ab | ♂ (43) | 2.16 ± 0.05 Ab | ♂ (29) | 2.48 ± 0.09 Aa | |
| 6th instar | ♀ (38) | 2.76 ± 0.19 Ab | ♀ (37) | 2.81 ± 0.06 Ab | ♀ (45) | 4.64 ± 0.10 Aa |
| ♂ (31) | 2.68 ± 0.21 Ab | ♂ (43) | 2.81 ± 0.06 Ab | ♂ (29) | 4.83 ± 0.11 Aa | |
| 7th instar ** | ♀ (31) | 4.74 ± 0.07 Aa | ♀ (37) | 4.35 ± 0.16 Aa | ♀ (2) | 5.00 ± 0.00 |
| ♂ (26) | 4.73 ± 0.16 Aa | ♂ (43) | 4.65 ± 0.10 Aa | ♂ (1) | 6.00 ± 0.00 | |
| 8th instar *** | ♀ (0) | - | ♀ (5) | 4.40 ± 0.24 A | ♀ (0) | - |
| ♂ (2) | 4.50 ± 0.50 a | ♂ (3) | 4.00 ± 0.00 Aa | ♂ (0) | - | |
| Pupa | ♀ (38) | 7.26 ± 0.08 Ba | ♀ (37) | 7.03 ± 0.06 Bab | ♀ (45) | 6.96 ± 0.07 Bb |
| ♂ (31) | 8.71 ± 0.09 Aa | ♂ (43) | 8.02 ± 0.06 Ab | ♂ (29) | 7.93 ± 0.06 Ab | |
| Egg to adult | ♀ (38) | 25.13 ± 0.21 Bb | ♀ (37) | 26.11 ± 0.17 Ba | ♀ (45) | 24.00 ± 0.18 Bc |
| ♂ (31) | 26.90 ± 0.21 Aa | ♂ (43) | 27.26 ± 0.16 Aa | ♂ (29) | 25.41 ± 0.24 Ab | |
| APOP | ♀ (30) | 4.63 ± 0.23 a | ♀ (36) | 3.03 ± 0.15 c | ♀ (32) | 3.84 ± 0.16 b |
| TPOP | ♀ (30) | 29.83 ± 0.20 a | ♀ (36) | 29.11 ± 0.19 a | ♀ (32) | 27.63 ± 0.27 b |
| Oviposition | ♀ (30) | 8.20 ± 0.46 a | ♀ (36) | 6.17 ± 0.40 b | ♀ (32) | 6.78 ± 0.57 ab |
| Adult longevity | ♀ (30) | 14.40 ± 0.88 Aa | ♀ (36) | 9.41 ± 0.39 Ab | ♀ (32) | 11.78 ± 0.56 Aab |
| ♂ (31) | 12.55 ± 0.82 Aa | ♂ (43) | 8.67 ± 0.42 Ab | ♂ (29) | 10.31 ± 0.57 Aab | |
| Fecundity (eggs/♀) | ♀ (30) | 1472.90 ± 108.38 a | ♀ (36) | 1021.19 ± 69.49 b | ♀ (32) | 1185.59 ± 93.76 ab |
| Host Plants | Population Parameters | ||||
|---|---|---|---|---|---|
| r (Day−1) * | λ (Day−1) | R0 | T (Day) | Female Ratio | |
| Napier grass | 0.1918 ± 0.0058 b | 1.2114 ± 0.0070 b | 465.12 ± 77.88 a | 32.02 ± 0.24 a | 0.40 |
| Natal grass | 0.1901 ± 0.0049 c | 1.2094 ± 0.0060 c | 382.94 ± 56.67 c | 31.28 ± 0.18 b | 0.38 |
| Sunn hemp | 0.1993 ± 0.0062 a | 1.2206 ± 0.0076 a | 395.19 ± 64.64 b | 29.98 ± 0.36 c | 0.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.-H.; Itza, B.; Kafle, L.; Chang, T.-Y. Life Table Study of Fall Armyworm (Spodoptera frugiperda) (Lepidoptera: Noctuidae) on Three Host Plants under Laboratory Conditions. Insects 2023, 14, 329. https://doi.org/10.3390/insects14040329
Chen W-H, Itza B, Kafle L, Chang T-Y. Life Table Study of Fall Armyworm (Spodoptera frugiperda) (Lepidoptera: Noctuidae) on Three Host Plants under Laboratory Conditions. Insects. 2023; 14(4):329. https://doi.org/10.3390/insects14040329
Chicago/Turabian StyleChen, Wen-Hua, Brandon Itza, Lekhnath Kafle, and Tsui-Ying Chang. 2023. "Life Table Study of Fall Armyworm (Spodoptera frugiperda) (Lepidoptera: Noctuidae) on Three Host Plants under Laboratory Conditions" Insects 14, no. 4: 329. https://doi.org/10.3390/insects14040329
APA StyleChen, W. -H., Itza, B., Kafle, L., & Chang, T. -Y. (2023). Life Table Study of Fall Armyworm (Spodoptera frugiperda) (Lepidoptera: Noctuidae) on Three Host Plants under Laboratory Conditions. Insects, 14(4), 329. https://doi.org/10.3390/insects14040329