
Paternity Analyses for the Planning of SIT Projects against the Red Palm Weevil


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Simulated Paternity Assignments
- The number of candidate fathers (i.e., the number of males the females mated with) in each simulation was 2, 4, 6, 8 and 10;
- The number of offspring produced was 150 larvae (which represented a true estimate of the average successful reproductive rate of laboratory-reared females [16]);
- The proportion of candidate fathers sampled was 1.00 (which was assumed to genotype all males involved in the experiment);
- The proportion of typed loci was 0.99 (based on the almost total absence of missing data from previous experiments due to the previous optimization of conditions for DNA extraction and amplification);
- The proportion of genotyping errors was 0.01 (i.e., the standard estimated error rate in the production of genotypic microsatellite data [44]).
2.2. Laboratory Mating Experiments
2.2.1. The Creation of Genetically Diverse Stocks of Individuals
2.2.2. Mating and Rearing
2.2.3. Genotyping and Paternity Analysis
3. Results
3.1. Simulations
3.2. Laboratory MATING Experiments
3.2.1. Mating and Rearing
3.2.2. Genotyping
3.2.3. Paternity Analysis
4. Discussion
4.1. Simulations
4.2. Paternity Analyses of Mating Experiments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murphy, S.T.; Briscoe, A.B.R. The Red Palm Weevil as an Alien Invasive: Biology and the Prospects for Biological Control as a Component of IPM. Biocontrol News Inf. 1999, 20, 35N–46N. [Google Scholar]
- Milosavljević, I.; El-Shafie, H.A.F.; Faleiro, J.R.; Hoddle, C.D.; Lewis, M.; Hoddle, M.S. Palmageddon: The Wasting of Ornamental Palms by Invasive Palm Weevils, Rhynchophorus spp. J. Pest Sci. 2019, 92, 143–156. [Google Scholar] [CrossRef]
- Faleiro, J.R. A Review of the Issues and Management of the Red Palm Weevil Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in Coconut and Date Palm during the Last One Hundred Years. Int. J. Trop. Insect Sci. 2006, 26, 135–154. [Google Scholar] [CrossRef]
- El-Mergawy, R.; Al-Ajlan, A.M. Red Palm Weevil, Rhynchophorus ferrugineus (Olivier): Economic Importance, Biology, Biogeography and Integrated Pest Management. J. Agric. Sci. Technol. 2011, 1, 1–23. [Google Scholar]
- Dembilio, Ó.; Jaques, J.A. Biology and Management of Red Palm Weevil. In Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges; Wakil, W., Romeno Faleiro, J., Miller, T.A., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 13–36. ISBN 9783319243979. [Google Scholar]
- Faghih, A.A. Others The Biology of Red Palm Weevil, Rhynchophorus ferrugineus Oliv (Coleoptera, Curculionidae) in Saravan Region (Sistan & Balouchistan Province, Iran). Appl. Entomol. Phytopathol. 1996, 63, 16–18. [Google Scholar]
- Kehat, M. Threat to Date Palms in Israel, Jordan and the Palestinian Authority, by the Red Palm Weevil, Rhynchophorus ferrugineus. Phytoparasitica 1999, 27, 241–242. [Google Scholar] [CrossRef]
- El-Shafie, H.A.F.; Faleiro, J.R. Red Palm Weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae): Global Invasion, Current Management Options, Challenges and Future Prospects. In Invasive Species-Introduction Pathways, Economic Impact, and Possible Management Options; IntechOpen: London, UK, 2020; pp. 1–30. [Google Scholar]
- Rugman-Jones, P.F.; Hoddle, C.D.; Hoddle, M.S.; Stouthamer, R. The Lesser of Two Weevils: Molecular-Genetics of Pest Palm Weevil Populations Confirm Rhynchophorus vulneratus (Panzer 1798) as a Valid Species Distinct from R. ferrugineus (Olivier 1790), and Reveal the Global Extent of Both. PLoS ONE 2013, 8, e78379. [Google Scholar] [CrossRef]
- Rochat, D.; Dembilio, O.; Jaques, J.A.; Suma, P.; Pergola, A.L.; Hamidi, R.; Kontodimas, D.; Soroker, V. Rhynchophorus ferrugineus: Taxonomy, Distribution, Biology, and Life Cycle. In Handbook of Major Palm Pests; Soroker, V., Colazza, S., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; Volume 12, pp. 69–104. ISBN 9781119057468. [Google Scholar]
- CABI. Invasive Species Compendium—Rhynchophorus ferrugineus (Red Palm Weevil). 2022. Available online: https://www.cabi.org/isc/datasheet/47472 (accessed on 19 October 2022).
- EPPO. EPPO Global Database—Rhynchophorus ferrugineus (RHYCFE). 2022. Available online: https://gd.eppo.int/taxon/RHYCFE (accessed on 19 October 2022).
- EPPO. EPPO A2 List of Pests Recommended for Regulation as Quarantine Pests—Version 2022-09. 2022. Available online: http://www.eppo.int/QUARANTINE/listA2.htm (accessed on 19 October 2022).
- Al-Dosary, N.M.; Al-Dobai, S.; Faleiro, J.R. Review on the management of Red Palm Weevil Rhynchophorus ferrugineus Olivier in date palm Phoenix dactylifera L. Emir. J. Food Agric. 2016, 28, 34–44. [Google Scholar] [CrossRef]
- Esteban Duran, J.; Yela, J.L.; Beitia Crespo, F.; Jimenez Alvarez, A. Biology of Red Palm Weevil Rhynchophorus ferrugineus (Olivier) in Laboratory and Field: Cycle in Captivity, Biological Characteristics in Its Introduction Zone in Spain and Biological Detection Methods and Possible Control (Coleoptera: Curculionidae: Rhynchophorinae). Bol. Sanid. Veg. Plagas 1998, 24, 737–748. [Google Scholar]
- Kaakeh, W. Longevity, fecundity, and fertility of the red palm weevil, Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae) on natural and artificial diets. Emir. J. Food Agric. 2005, 17, 23–33. [Google Scholar] [CrossRef]
- Prabhu, S.T.; Patil, R.S. Studies on the Biological Aspects of Red Palm Weevil, Rhynchophorus ferrugineus (Oliv.). Karnataka J. Agric. Sci. 2009, 22, 732–733. [Google Scholar]
- Ju, R.-T.; Wang, F.; Wan, F.-H.; Li, B. Effect of Host Plants on Development and Reproduction of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). J. Pest Sci. 2011, 84, 33–39. [Google Scholar] [CrossRef]
- Dembilio, O.; Jacas, J.A. Basic Bio-Ecological Parameters of the Invasive Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in Phoenix canariensis under Mediterranean Climate. Bull. Entomol. Res. 2011, 101, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Francardi, V.; Inghilesi, A.F.; Stasolla, G.; Benvenuti, C.; Cini, A.; Barzanti, G.P.; Cito, A.; Arnone, S.; Cristofaro, M.; et al. Analisi del comportamento sociale e riproduttivo dei maschi sterili di Rhynchophorus ferrugineus: Implicazioni per il controllo biologico con agenti entomopatogeni. In Proceedings of the XXIV Congresso Nazionale Italiano di Entomologia, Orosei, NU, Italy, 9–14 June 2014. [Google Scholar]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, MA, USA, 2021; ISBN 9781003035572. [Google Scholar]
- Cristofaro, M.; Sforza, R.F.H.; Roselli, G.; Paolini, A.; Cemmi, A.; Musmeci, S.; Anfora, G.; Mazzoni, V.; Grodowitz, M. Effects of Gamma Irradiation on the Fecundity, Fertility, and Longevity of the Invasive Stink Bug Pest Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae). Insects 2022, 13, 787. [Google Scholar] [CrossRef]
- Paoli, F.; Dallai, R.; Cristofaro, M.; Arnone, S.; Francardi, V.; Roversi, P.F. Morphology of the Male Reproductive System, Sperm Ultrastructure and γ-Irradiation of the Red Palm Weevil Rhynchophorus ferrugineus Oliv. (Coleoptera: Dryophthoridae). Tissue Cell 2014, 46, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, I. Mechanisms of Sperm Competition in Insects. Ann. Zool. Fenn. 1998, 35, 241–257. [Google Scholar]
- Shuker, D.M.; Simmons, L.W. The Evolution of Insect Mating Systems; Oxford University Press: Oxford, UK, 2014; ISBN 9780199678020. [Google Scholar]
- Eberhard, W. Female Control: Sexual Selection by Cryptic Female Choice; Princeton University Press: Princeton, NJ, USA, 1996; ISBN 9780691010847. [Google Scholar]
- Córdoba-Aguilar, A.; Uhía, E.; Cordero Rivera, A. Sperm Competition in Odonata (Insecta): The Evolution of Female Sperm Storage and Rivals’ Sperm Displacement. J. Zool. 2003, 261, 381–398. [Google Scholar] [CrossRef] [Green Version]
- Siva-Jothy, M.T.; Blake, D.E.; Thompson, J.; Ryder, J.J. Short-and Long-Term Sperm Precedence in the Beetle Tenebrio molitor: A Test of the “adaptive Sperm Removal” Hypothesis. Physiol. Entomol. 1996, 21, 313–316. [Google Scholar] [CrossRef]
- Gregory, G.E. The formation and fate of the spermatophore in the African migratory locust, Locusta migratoria migratorioides Reiche and Fairmaire. Trans. R. Entomol. Soc. Lond. 1965, 117, 33–66. [Google Scholar] [CrossRef]
- Ono, T.; Siva-Jothy, M.T.; Kato, A. Removal and Subsequent Ingestion of Rivals’ Semen during Copulation in a Tree Cricket. Physiol. Entomol. 1989, 14, 195–202. [Google Scholar] [CrossRef]
- Harshman, L.G.; Prout, T. Sperm displacement without sperm transfer in Drosophila melanogaster. Evolution 1994, 48, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Otronen, M.; Siva-Jothy, M.T. The Effect of Postcopulatory Male Behaviour on Ejaculate Distribution within the Female Sperm Storage Organs of the Fly, Dryomyza anilis (Diptera: Dryomyzidae). Behav. Ecol. Sociobiol. 1991, 29, 33–37. [Google Scholar] [CrossRef]
- Parker, G.A. Sperm Competition and Its Evolutionary Consequences in The Insects. Biol. Rev. Camb. Philos. Soc. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Parker, G.A. Sperm Competition and the Evolution of Animal Mating Strategies. In Sperm Competition and the Evolution of Animal Mating Systems; Academic Press: Cambridge, MA, USA, 1984; pp. 1–60. [Google Scholar]
- Gwynne, D.T. Male mating effort, confidence of paternity, and insect sperm competition. In Sperm Competition and the Evolution of Animal Mating Systems; Smith, R.L., Ed.; Academic Press: New York, NY, USA, 1984; pp. 117–149. [Google Scholar]
- Ridley, M. The Incidence of Sperm Displacement in Insects: Four Conjectures, One Corroboration. Biol. J. Linn. Soc. Lond. 1989, 38, 349–367. [Google Scholar] [CrossRef]
- Boorman, E.; Parker, G.A. Sperm (ejaculate) Competition in Drosophila melanogaster, and the Reproductive Value of Females to Males in Relation to Female Age and Mating Status. Ecol. Entomol. 1976, 1, 145–155. [Google Scholar] [CrossRef]
- Musmeci, S.; Cristofaro, M.; Arnone, S.; Sasso, R.; Baccaro, S.; Pasquali, A.; Catarci, S. Controllo del Punteruolo rosso mediante la tecnica del maschio sterile (SIT): Utopia o realtà? Il punteruolo rosso delle palme: Nuove acquisizioni e possibilità di controllo demografico. Atti Accad. Naz. Ital. Di Entomol. 2014, 61, 239–246. [Google Scholar]
- Musmeci, S.; Belvedere, S.; Sasso, R.; Arnone, S.; Cristofaro, M.; Nobili, P.; La Marca, A.; De Biase, A. Last-Male Sperm Precedence in Rhynchophorus ferrugineus (Olivier): Observations in Laboratory Mating Experiments with Irradiated Males. Bull. Entomol. Res. 2018, 108, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.G.; Small, C.M.; Paczolt, K.A.; Ratterman, N.L. A Practical Guide to Methods of Parentage Analysis. Mol. Ecol. Resour. 2010, 10, 6–30. [Google Scholar] [CrossRef]
- Belvedere, S.; La Marca, A.; Marcari, V.; Senia, G.; De Biase, A. In Silico Mining of Microsatellite Markers for the Red Palm Weevil as Contribution to Its Invasion Management. Conserv. Genet. Resour. 2014, 6, 947–948. [Google Scholar] [CrossRef]
- Molecular Ecology Resources Primer Development Consortium; A’Hara, S.W.; Amouroux, P.; Argo, E.E.; Avand-Faghih, A.; Barat, A.; Barbieri, L.; Bert, T.M.; Blatrix, R.; Blin, A.; et al. Permanent Genetic Resources Added to Molecular Ecology Resources Database 1 August 2011-30 September 2011. Mol. Ecol. Resour. 2012, 12, 185–189. [Google Scholar] [CrossRef]
- De Biase, A.; Belvedere, S.; Senia, G.; Marcari, V.; La Marca, A.; Vu, L.V.; Cristofaro, M. Preliminary analysis of genetic variability and differentiation of Asian and Mediterranean populations of Red Palm Weevil. Atti Accad. Naz. Ital. Entomol. 2014, 61, 193–200. [Google Scholar]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising How the Computer Program CERVUS Accommodates Genotyping Error Increases Success in Paternity Assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Cristofaro, M.; Colazza, S.; De Biase, A.; Belvedere, S.; Arnone, S.; Di Ilio, V.; Isidoro, N.; Musmeci, S.; Sasso, R.; Barlattani, M. Preliminary studies of the biological cycle of the Red Palm Weevil (Rhynchophorus ferrugineus) in Italy, aimed to the application of Sterile Insect Technique. In Proceedings of the ESA 58th Annual Meeting, San Diego, CA, USA, 12–15 December 2010. Abstract poster D0487. [Google Scholar]
- Mack, P.D.; Hammock, B.A.; Promislow, D.E.L. Sperm Competitive Ability and Genetic Relatedness in Drosophila melanogaster: Similarity Breeds Contempt. Evolution 2002, 56, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Gaskin, J.F.; Bon, M.-C.; Cock, M.J.W.; Cristofaro, M.; Biase, A.D.; De Clerck-Floate, R.; Ellison, C.A.; Hinz, H.L.; Hufbauer, R.A.; Julien, M.H.; et al. Applying Molecular-Based Approaches to Classical Biological Control of Weeds. Biol. Control 2011, 58, 1–21. [Google Scholar] [CrossRef]
- Cristofaro, M.; De Biase, A.; Smith, L. Field Release of a Prospective Biological Control Agent of Weeds, Ceratapion basicorne, to Evaluate Potential Risk to a Nontarget Crop. Biol. Control 2013, 64, 305–314. [Google Scholar] [CrossRef]
- Palero, F.; González-Candelas, F.; Pascual, M. MICROSATELIGHT—Pipeline to Expedite Microsatellite Analysis. J. Hered. 2011, 102, 247–249. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F. Micro-checker: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Mol. Ecol. 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Dodds, K.G.; Tate, M.L.; McEwan, J.C.; Crawford, A.M. Exclusion Probabilities for Pedigree Testing Farm Animals. Theor. Appl. Genet. 1996, 92, 966–975. [Google Scholar] [CrossRef]
- Marshall, T.C.; Slate, J.; Kruuk, L.E.; Pemberton, J.M. Statistical Confidence for Likelihood-Based Paternity Inference in Natural Populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [Green Version]
- Signorovitch, J.; Nielsen, R. PATRI-Paternity Inference Using Genetic Data. Bioinformatics 2002, 18, 341–342. [Google Scholar] [CrossRef] [Green Version]
- Margeta, P.; Skorput, D.; Salamon, D.; Mencik, S.; Gvozdanović, K.; Karolyi, D.; Lukovic, Z.; Salajpal, K. 12-Plex Highly Polymorphic Microsatellite Marker Set for Parentage Analysis in Banija Spotted Pigs. J. Cent. Eur. Agric. 2019, 20, 50–54. [Google Scholar] [CrossRef]
- Satyanarayana, Y.; Babu, P.G.; Jahageerdar, S. Parentage Determination in the Freshwater Prawn Macrobrachium Rosenbergii De Man, 1879 Using Microsatellite Markers. Indian J. Fish. 2015, 62, 29–32. [Google Scholar]
- Hallett, R.H.; Gries, G.; Gries, R.; Borden, J.H.; Czyzewska, E.; Oehlschlager, A.C.; Pierce, H.D.; Angerilli, N.P.D.; Rauf, A. Aggregation Pheromones of Two Asian Palm Weevils, Rhynchophorus ferrugineus and R. vulneratus. Naturwissenschaften 1993, 80, 328–331. [Google Scholar] [CrossRef]
- Inghilesi, A.F.; Mazza, G.; Cervo, R.; Cini, A. A Network of Sex and Competition: The Promiscuous Mating System of an Invasive Weevil. Curr. Zool. 2015, 61, 85–97. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Common Parameters | |
|---|---|
| Number of offspring/female | 150 |
| Proportion of sampled candidate fathers | 1.00 |
| Proportion of typed loci | 0.99 |
| Proportion of genotyping errors | 0.01 |
| Likelihood error rate | 0.01 (default) |
| Minimum number of typed loci/individual | 8.00 |
| Variable Parameters | |
| Number of candidate fathers | 2; 4; 6; 8; 10 |
| Common Parameters | |
|---|---|
| Number of offspring/female | 150 |
| Proportion of typed loci | 0.99 |
| Proportion of genotyping errors | 0.01 |
| Likelihood error rate | 0.01 (default) |
| Minimum number of typed loci/individuals | 8.00 |
| Variable Parameters | |
| Number of candidate fathers | 5; 10; 20; 30; 40 |
| Proportion of sampled candidate fathers | 0.25; 0.5; 0.75; 1.00 |
| Locus | NA 1 | N 2 | PIC 3 | NE-2P 4 |
|---|---|---|---|---|
| RPW02 | 3 | 22 | 0.360 | 0.802 |
| RPW03 | 4 | 22 | 0.405 | 0.758 |
| RPW06 | 7 | 21 | 0.712 | 0.467 |
| RPW11 | 2 | 22 | 0.208 | 0.896 |
| RPW16 | 2 | 22 | 0.348 | 0.826 |
| RPW20 | 2 | 22 | 0.370 | 0.815 |
| RPW22 | 2 | 22 | 0.208 | 0.896 |
| RPW25 | 2 | 22 | 0.318 | 0.841 |
| RPW26 | 2 | 22 | 0.305 | 0.848 |
| RPW32 | 2 | 22 | 0.330 | 0.835 |
| RPW36 | 2 | 22 | 0.305 | 0.848 |
| P4C2 | 4 | 22 | 0.617 | 0.593 |
| P1A3 | 5 | 22 | 0.473 | 0.698 |
| Number of Candidate Fathers | |||||
|---|---|---|---|---|---|
| 2 | 4 | 6 | 8 | 10 | |
| Assignment rate | 100% | 100% | 100% | 99% | 89% |
| Number of Candidate Fathers | ||||||
|---|---|---|---|---|---|---|
| 5 | 10 | 20 | 30 | 40 | ||
| Proportion of Sampled Candidate Fathers | 0.25 | 33% | 19% | 18% | 15% | 5% |
| 0.50 | 55% | 33% | 29% | 24% | 13% | |
| 0.75 | 75% | 59% | 57% | 49% | 28% | |
| 1.00 | 100% | 89% | 67% | 49% | 32% | |
| Experiment 1 | Experiment 2 | |
|---|---|---|
| Oviposition Period (days) | 29 | 28 |
| Number of Oviposition Events | 14 | 17 |
| Number of Eggs Laid | 175 | 192 |
| Number of Eggs Hatched (%) | 127 (72.57%) | 132 (68.75%) |
| Number of Larvae that Survived | 81 | 125 |
| Experiment 1 | Experiment 2 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Locus | Female | Male 1 | Male 2 | Female | Male 1 | Male 2 | ||||||
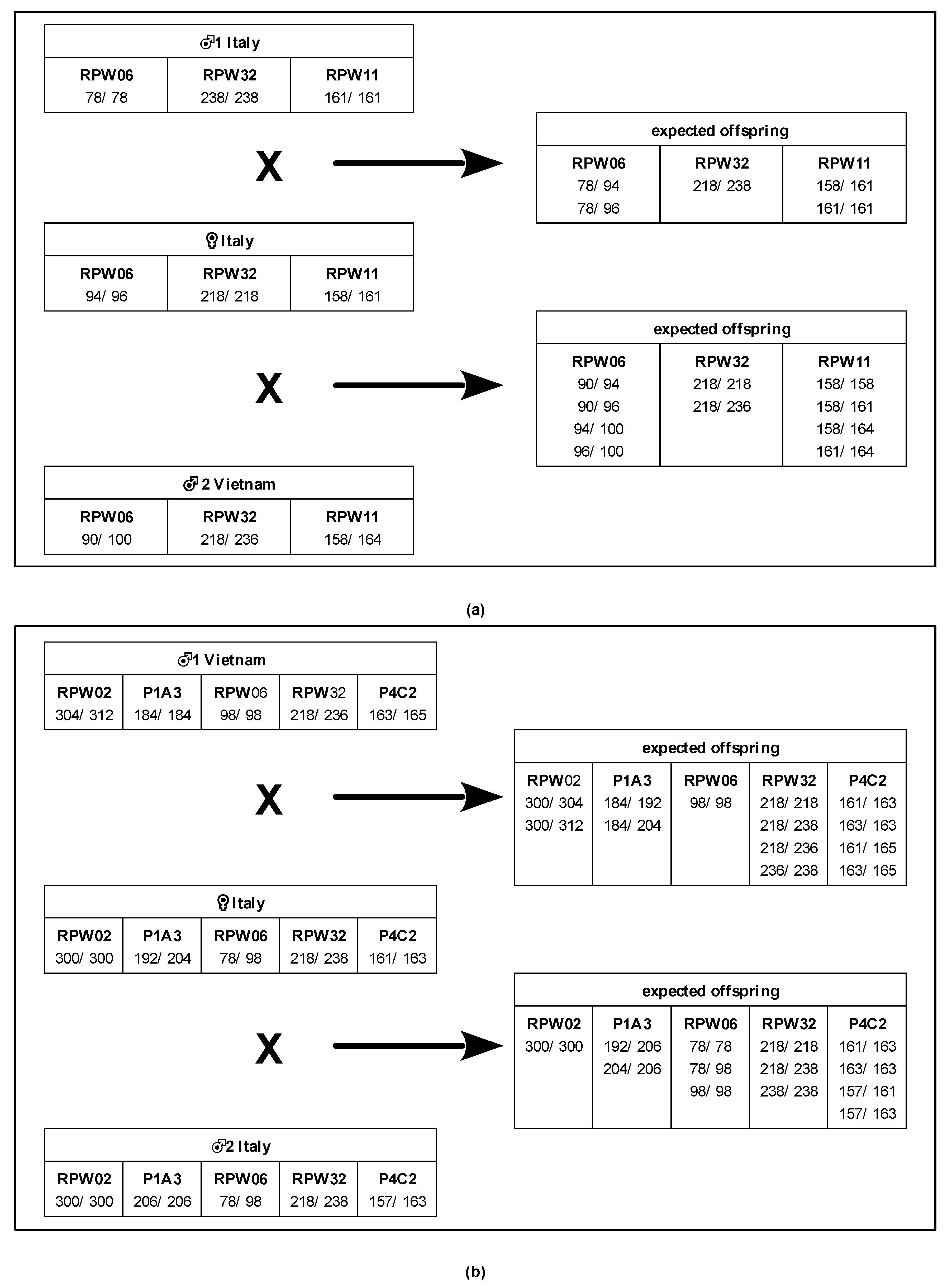
| RPW02 | 300 | 300 | 300 | 300 | 292 | 292 | 300 | 300 | 304 | 312 | 300 | 300 |
| RPW03 | - | - | 214 | 214 | 214 | 214 | - | - | 212 | 214 | 212 | 214 |
| RPW06 | 94 | 96 | 78 | 78 | 90 | 100 | 78 | 98 | 98 | 98 | 78 | 98 |
| RPW11 | 158 | 161 | 161 | 161 | 158 | 164 | 161 | 161 | 158 | 158 | 158 | 161 |
| RPW13 | - | - | 172 | 172 | 172 | 172 | - | - | 172 | 172 | 172 | 172 |
| RPW16 | - | - | 226 | 229 | 226 | 226 | 226 | 226 | 226 | 226 | 226 | 229 |
| RPW17 | - | - | 222 | 222 | 219 | 222 | 222 | 222 | 219 | 222 | 222 | 222 |
| RPW20 | - | - | 82 | 86 | 86 | 86 | 82 | 86 | 86 | 86 | 82 | 86 |
| RPW22 | - | - | 159 | 163 | 159 | 159 | 163 | 163 | 159 | 163 | 163 | 163 |
| RPW24 | - | - | 95 | 95 | 95 | 95 | - | - | 95 | 95 | 95 | 95 |
| RPW25 | - | - | 99 | 99 | 99 | 99 | - | - | 99 | 99 | 99 | 99 |
| RPW26 | - | - | 140 | 140 | 132 | 140 | - | - | 140 | 140 | 140 | 140 |
| RPW32 | 218 | 218 | 238 | 238 | 218 | 236 | 218 | 238 | 218 | 236 | 218 | 238 |
| RPW36 | - | - | 143 | 145 | 145 | 145 | 143 | 143 | 143 | 145 | 145 | 145 |
| P1A3 | 184 | 204 | 192 | 204 | 184 | 184 | 192 | 204 | 184 | 184 | 206 | 206 |
| P4C2 | - | - | 157 | 163 | 157 | 157 | 161 | 163 | 163 | 165 | 157 | 163 |
 Totally diagnostic loci: No shared alleles between the males or between the female and either male.
Totally diagnostic loci: No shared alleles between the males or between the female and either male.  Partially diagnostic loci: Males had one shared allele and/or the female shared an allele with at least one of the males.
Partially diagnostic loci: Males had one shared allele and/or the female shared an allele with at least one of the males.  Not diagnostic loci: Males had the same genotypes. Not genotyped because the locus was excluded from the subsequent analyses on the basis of the male genotypes. The numbers refer to the allele dimensions.
Not diagnostic loci: Males had the same genotypes. Not genotyped because the locus was excluded from the subsequent analyses on the basis of the male genotypes. The numbers refer to the allele dimensions.| Locus | k | N | Ho | He | PIC | NE-1P | NE-2P | NE-PP | NE-I | NE-SI |
|---|---|---|---|---|---|---|---|---|---|---|
| Experiment 1 | ||||||||||
| RPW06 | 13 | 38 | 0.553 | 0.850 | 0.823 | 0.472 | 0.307 | 0.130 | 0.042 | 0.341 |
| RPW11 | 5 | 40 | 0.375 | 0.599 | 0.514 | 0.816 | 0.684 | 0.530 | 0.245 | 0.515 |
| RPW32 | 3 | 40 | 0.325 | 0.391 | 0.349 | 0.925 | 0.803 | 0.676 | 0.414 | 0.660 |
| Mean/Combined | 0.562 | 0.357 | 0.168 | 0.047 | 0.004 | 0.116 | ||||
| Experiment 2 | ||||||||||
| RPW02 | 7 | 38 | 0.368 | 0.711 | 0.665 | 0.696 | 0.516 | 0.318 | 0.125 | 0.431 |
| P1A3 | 13 | 40 | 0.450 | 0.816 | 0.789 | 0.525 | 0.351 | 0.157 | 0.055 | 0.361 |
| RPW06 | 13 | 38 | 0.553 | 0.847 | 0.820 | 0.478 | 0.311 | 0.132 | 0.043 | 0.343 |
| RPW32 | 3 | 40 | 0.350 | 0.406 | 0.360 | 0.919 | 0.796 | 0.668 | 0.400 | 0.649 |
| P4C2 | 9 | 38 | 0.500 | 0.772 | 0.726 | 0.628 | 0.450 | 0.263 | 0.093 | 0.392 |
| Mean/Combined | 0.672 | 0.101 | 0.020 | 0.001 | 0.000 | 0.014 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belvedere, S.; Arnone, S.; Cristofaro, M.; La Marca, A.; De Biase, A. Paternity Analyses for the Planning of SIT Projects against the Red Palm Weevil. Insects 2023, 14, 326. https://doi.org/10.3390/insects14040326
Belvedere S, Arnone S, Cristofaro M, La Marca A, De Biase A. Paternity Analyses for the Planning of SIT Projects against the Red Palm Weevil. Insects. 2023; 14(4):326. https://doi.org/10.3390/insects14040326
Chicago/Turabian StyleBelvedere, Silvia, Silvia Arnone, Massimo Cristofaro, Alessandra La Marca, and Alessio De Biase. 2023. "Paternity Analyses for the Planning of SIT Projects against the Red Palm Weevil" Insects 14, no. 4: 326. https://doi.org/10.3390/insects14040326
APA StyleBelvedere, S., Arnone, S., Cristofaro, M., La Marca, A., & De Biase, A. (2023). Paternity Analyses for the Planning of SIT Projects against the Red Palm Weevil. Insects, 14(4), 326. https://doi.org/10.3390/insects14040326