


Suitability of Raycell MK2 Blood X-ray Irradiator for the Use in the Sterile Insect Technique: Dose Response in Fruit Flies, Tsetse Flies and Mosquitoes

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Irradiation Set-Up
2.2. Dosimetry
2.3. Dose Response of Ceratitis capitata and Anastrepha ludens Pupae under Hypoxic Conditions
2.3.1. Strains and Rearing
2.3.2. Irradiation Procedure and Assessment of Sterility
2.4. Dose Response of Glossina palpalis gambiensis Pupae
2.4.1. Strain and Rearing
2.4.2. Irradiation Procedure and Assessment of Sterility
2.5. Dose Response of Aedes aegypti and Anopheles arabiensis Pupae and Adults
2.5.1. Strains and Rearing
2.5.2. Irradiation Procedure and Assessment of Sterility
2.6. Statistical Analyses
3. Results
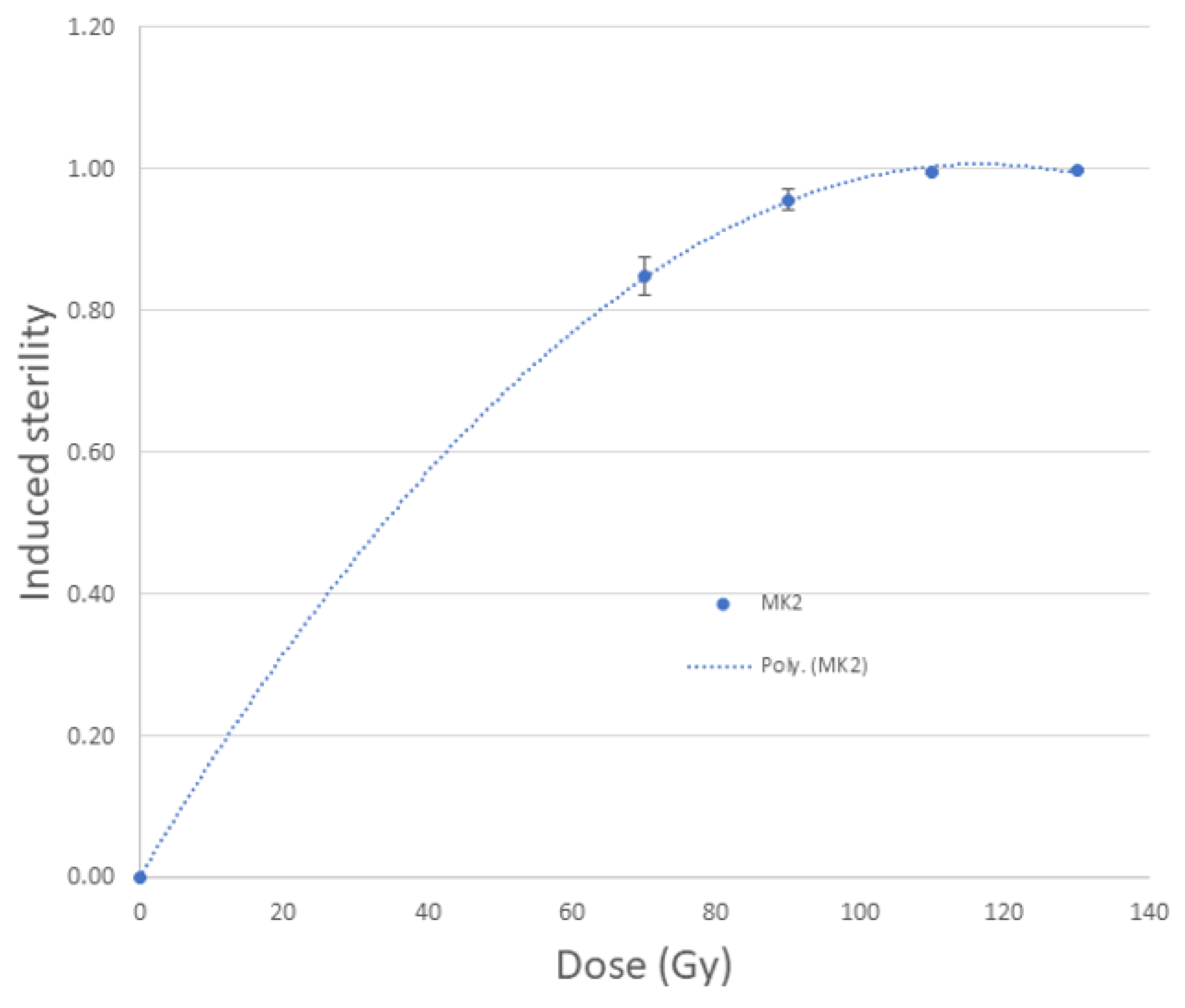
3.1. Dosimetry
3.2. Sterilization Efficiency of Raycell MK2
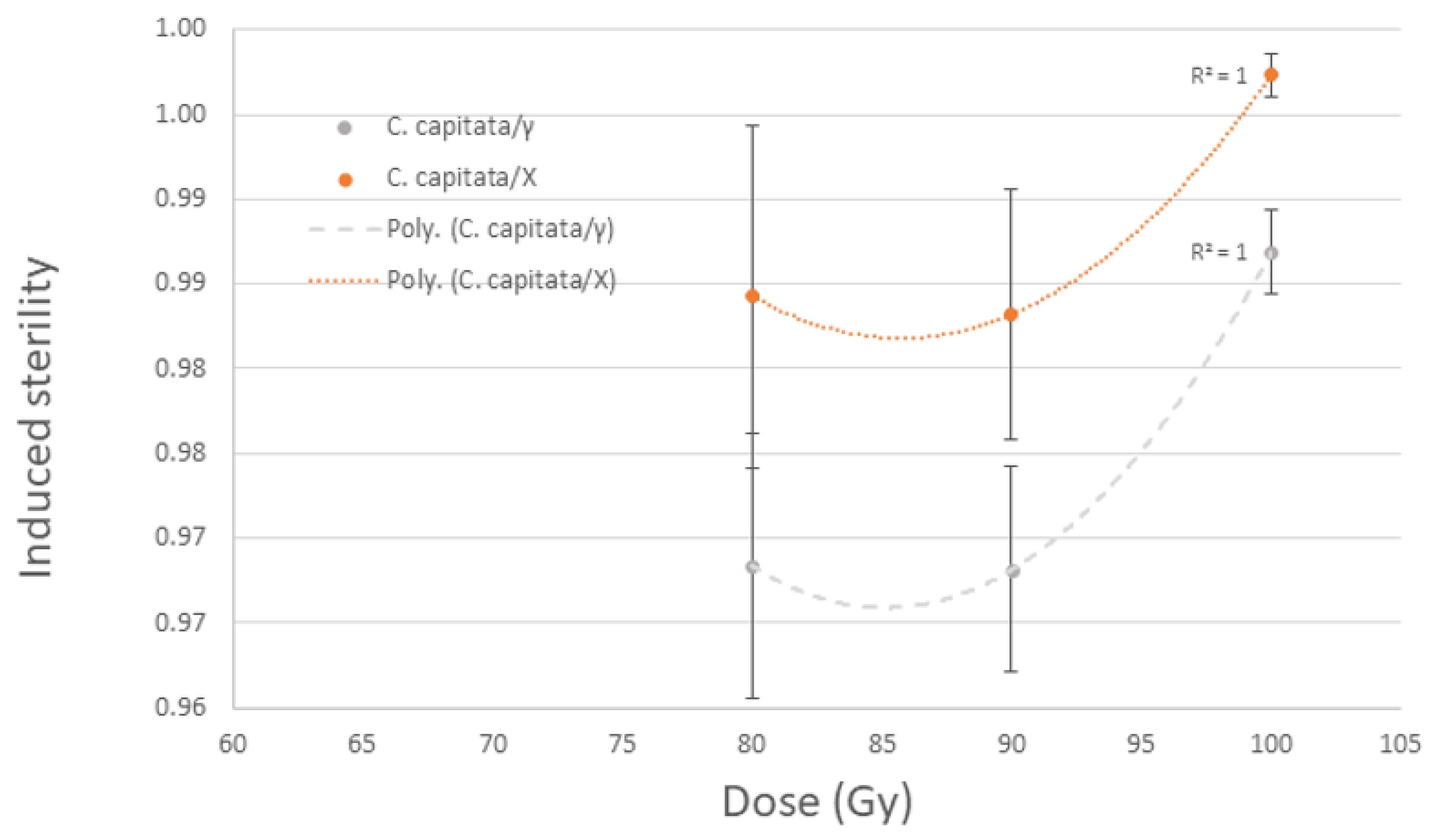
3.2.1. Ceratitis capitata and Anastrepha ludens Pupae
3.2.2. Glossina palpalis gambiensis Pupae
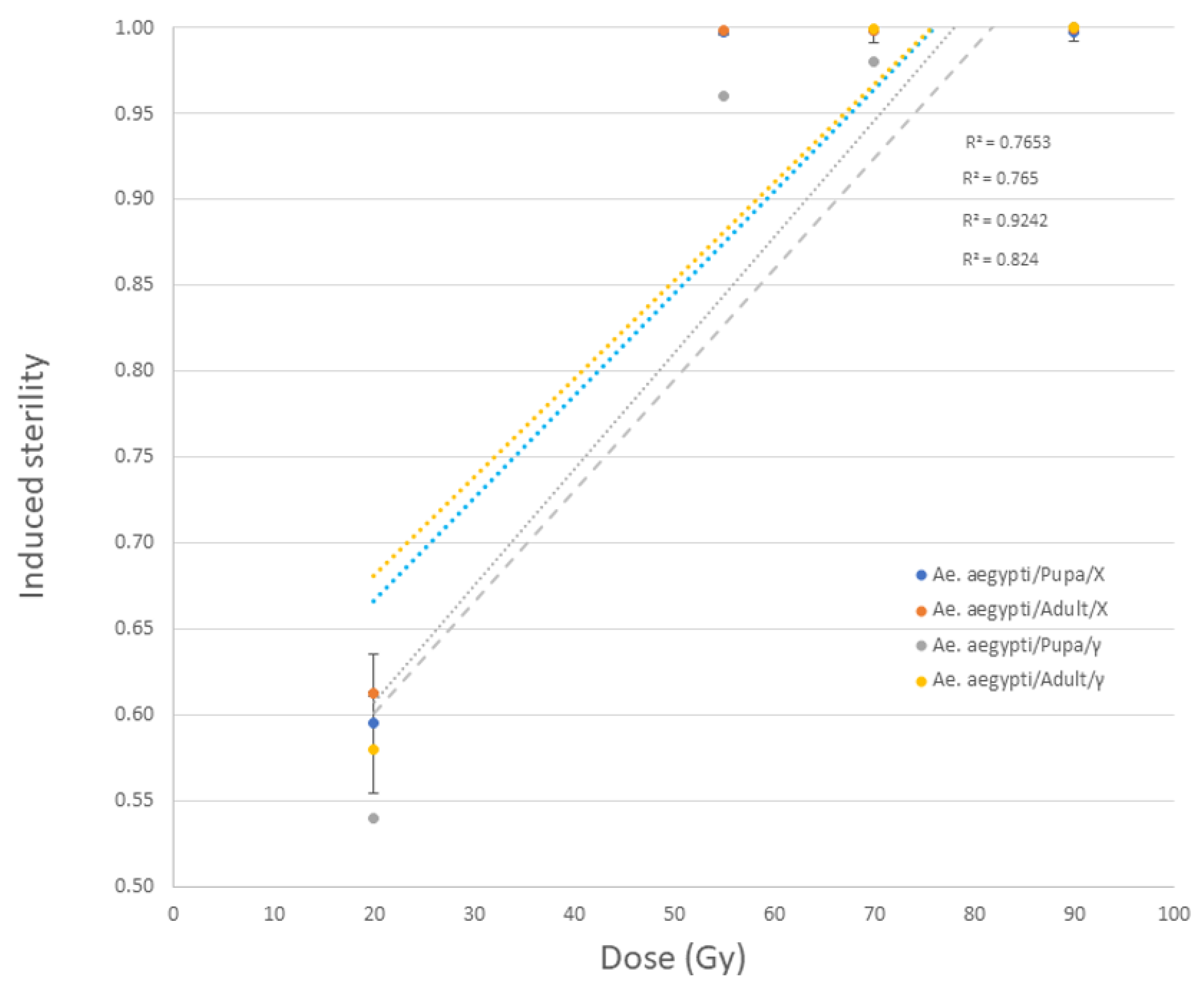
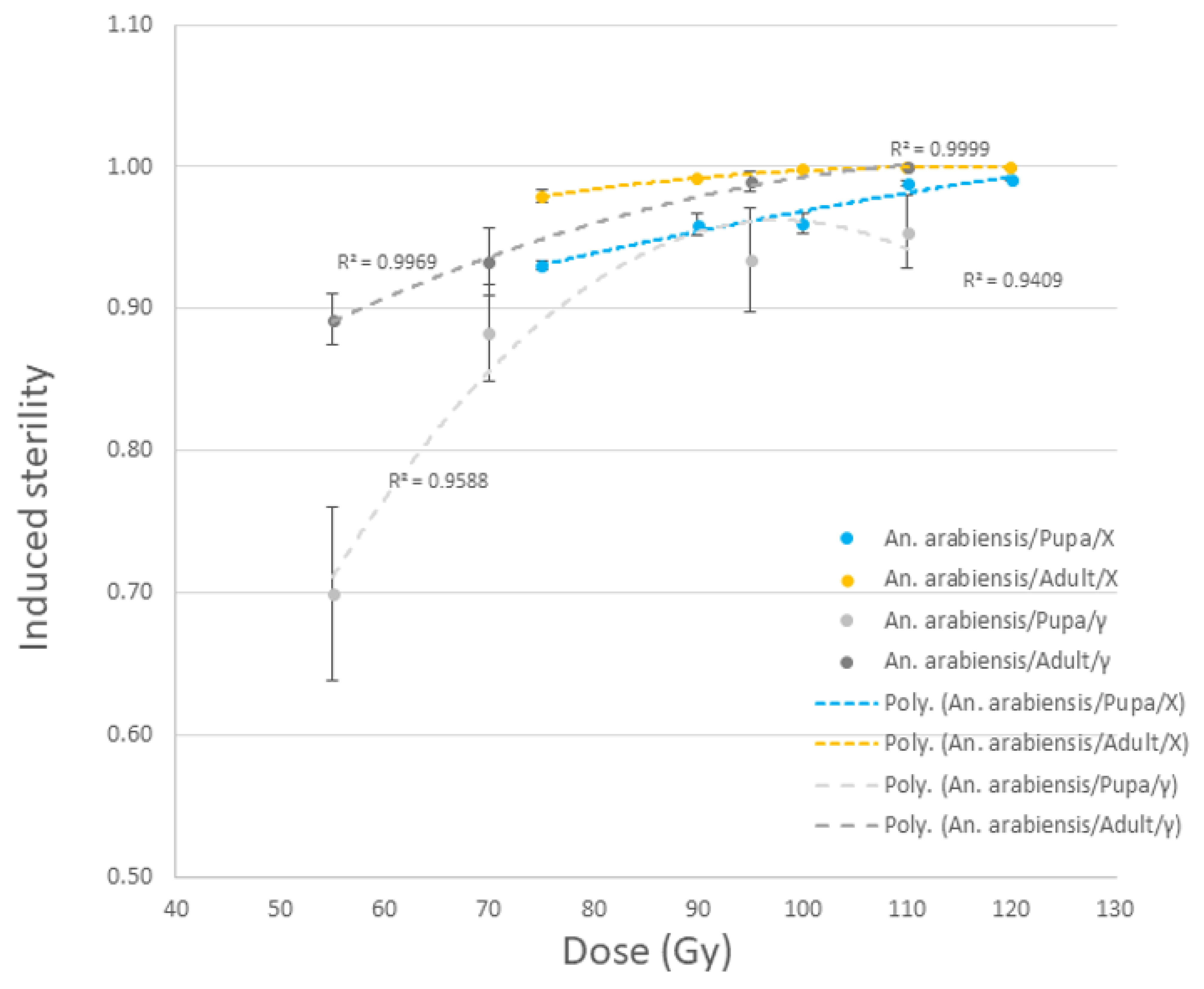
3.2.3. Aedes aegypti and Anopheles arabiensis Pupae and Adults
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. (Eds.) Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Klassen, W.; Vreysen, M.J.B. Area-Wide Integrated Pest Management and the Sterile Insect Technique. In The Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 75–112. [Google Scholar]
- Klassen, W.; Curtis, C.F.; Hendrichs, J. History of the Sterile Insect Technique. In The Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 1–44. [Google Scholar]
- Vreysen, M.J.; Abd-Alla, A.M.; Bourtzis, K.; Bouyer, J.; Caceres, C.; de Beer, C.; Oliveira Carvalho, D.; Maiga, H.; Mamai, W.; Nikolouli, K. The Insect Pest Control Laboratory of the Joint FAO/IAEA Programme: Ten Years (2010–2020) of Research and Development, Achievements and Challenges in Support of the Sterile Insect Technique. Insects 2021, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Bakri, A.; Mehta, K.; Lance, D. Sterilizing Insects with Ionizing Radiation. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J.P., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 355–398. [Google Scholar]
- Gómez-Simuta, Y.; Parker, A.; Cáceres, C.; Vreysen, M.J.B.; Yamada, H. Characterization and Dose-Mapping of an X-ray Blood Irradiator to Assess Application Potential for the Sterile Insect Technique (SIT). Appl. Radiat. Isot. 2021, 176, 109859. [Google Scholar] [CrossRef] [PubMed]
- Bushland, R.C.; Hopkins, D.E. Experiments with Screw-Worm Flies Sterilized by X-rays. J.Econ.Entomol. 1951, 44, 725–731. [Google Scholar] [CrossRef]
- Dey, S.K.; Manna, G.K. Differential Stage Sensitivity to X-rays in a Bug Physopelta schlanbuschi. Natl. Acad. Sci. Lett. 1983, 6, 101–103. [Google Scholar]
- Mastrangelo, T.; Parker, A.G.; Jessup, A.; Pereira, R.; Orozco-Dávila, D.; Islam, A.; Dammalage, T.; Walder JM, M. A new generation of X ray irradiators for insect sterilization. J. Econ. Entomol. 2010, 103, 85–94. [Google Scholar] [CrossRef]
- Mehta, K.; Parker, A. Characterization and Dosimetry of a Practical X-ray Alternative to Self-Shielded Gamma Irradiators. Radiat. Phys. Chem. 2011, 80, 107–113. [Google Scholar] [CrossRef]
- Ndo, C.; Yamada, H.; Damiens, D.D.; N’do, S.; Seballos, G.; Gilles, J.R.L. X-ray Sterilization of the An. Arabiensis Genetic Sexing Strain ‘ANO IPCL1’ at Pupal and Adult Stages. Acta Trop. 2014, 131, 124–128. [Google Scholar] [CrossRef]
- Yamada, H.; Parker, A.G.; Oliva, C.F.; Balestrino, F.; Gilles, J.R.L. X-ray-Induced Sterility in Aedes Albopictus (Diptera: Culicidae) and Male Longevity Following Irradiation. J. Med. Entomol. 2014, 51, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Light, D.M.; Ovchinnikova, I.; Jackson, E.S.; Haff, R.P. Effects of x-ray irradiation on male navel orangeworm moths (Lepidoptera: Pyralidae) on mating, fecundity, fertility, and inherited sterility. J. Econ. Entomol. 2015, 108, 2200–2212. [Google Scholar] [CrossRef] [Green Version]
- Mehta, K. Technical Specification for an X-ray System for the Irradiation of Insects for the Sterile Insect Technique and other related technologies; IAEA: Vienna, Austria, 2017. [Google Scholar]
- IAEA Dosimetry for SIT: Standard Operating Procedures for Gafchromic™ Film Dosimetry System for Low Energy X Radiation v1.0. 2022. Available online: https://www.iaea.org/resources/manual/dosimetry-for-sit-standard-operating-procedures-for-gafchromictm-film-dosimetry-system-for-low-energy-x-radiation-v10 (accessed on 10 November 2022).
- Franz, G. Genetic Sexing Strains in Mediterranean Fruit Fly, an Example for Other Species Amenable to Large-Scale Rearing for the Sterile Insect Technique. In The Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Zepeda-Cisneros, C.S.; Hernández, J.S.M.; Garcáa-Martánez, V.; Ibañez-Palacios, J.; Zacharopoulou, A.; Franz, G. Development, Genetic and Cytogenetic Analyses of Genetic Sexing Strains of the Mexican Fruit Fly, Anastrepha Ludens Loew (Diptera: Tephritidae). BMC Genet. 2014, 15, S1. [Google Scholar] [CrossRef] [Green Version]
- Meza-Hernández, J.S.; Dáaz-Fleischer, F. Comparison of Sexual Compatibility between Laboratory and Wild Mexican Fruit Flies under Laboratory and Field Conditions. J. Econ. Entomol. 2006, 99, 1979–1986. [Google Scholar] [CrossRef]
- Kyritsis, G.A.; Augustinos, A.A.; Caceres, C.; Bourtzis, K. Medfly Gut Microbiota and Enhancement of the Sterile Insect Technique: Similarities and Differences of Klebsiella Oxytoca and Enterobacter Sp. AA26 Probiotics during the Larval and Adult Stages of the VIENNA 8D53+ Genetic Sexing Strain. Front. Microbiol. 2017, 8, 2064. [Google Scholar] [CrossRef]
- Schwarz, A.J.; Zambada, A.; Orozco, D.H.S.; Zavala, J.L.; Calkins, C.O. Mass Production of the Mediterranean Fruit Fly at Metapa, Mexico. Fla. Entomol. 1985, 68, 467–477. [Google Scholar] [CrossRef]
- FAO/IAEA/USDA. Manual for Product Quality Control and Shipping Procedures for Sterile Mass-Reared Tephritid Fruit Flies; IAEA: Vienna, Austria, 2003; Volume 5. [Google Scholar]
- Mutika, G.N.; Kabore, I.; Seck, M.T.; Sall, B.; Bouyer, J.; Parker, A.G.; Vreysen, M.J.B. Mating Performance of Glossina Palpalis Gambiensis Strains from Burkina Faso, Mali, and Senegal. Entomol. Exp. Et Appl. 2013, 146, 177–185. [Google Scholar] [CrossRef]
- Feldmann, U. Rearing Tsetse Flies for Use in Sterile Insect Technique Vector Control Programmes. In Proceedings of the International Symposium on Mangement of Insect pests: Nuclear Related Molecular and Genetic Techniques, Vienna, Austria, 19–23 October 1992; IAEA/FAO, Ed.; IAEA: Vienna, Austria, 1992; pp. 579–601. [Google Scholar]
- FAO/IAEA Standard Operating Procedures for Mass-Rearing Tsetse Flies, Draft Version. 2006. Available online: https://www.iaea.org/sites/default/files/21/06/nafa-ipc-manual-tsetse_rearing_sop_web.pdf (accessed on 7 February 2022).
- FAO/IAEA. Guidelines for Routine Colony Maintenance of Aedes Mosquito Species—Version 1.0. 2017, p. 18. Available online: https://www.iaea.org/resources/manual/guidelines-for-routine-colony-maintenance-of-aedes-mosquito-species-version-10 (accessed on 2 September 2022).
- FAO/IAEA. Guidelines for Standardised Mass Rearing of Anopheles Mosquitoes—Version 1.0. 2017, p. 44. Available online: https://www.iaea.org/resources/manual/guidelines-for-standardised-mass-rearing-of-anopheles-mosquitoes-version-10 (accessed on 2 September 2022).
- Focks, D.A. An Improved Separator for the Developmental Stages, Sexes, and Species of Mosquitoes (Diptera: Culicidae). J. Med. Entomol. 1980, 17, 567–568. [Google Scholar] [CrossRef]
- FAO/IAEA. Guidelines for Irradiation of Mosquito Pupae in Sterile Insect Technique Programmes-version 1.0. 2020. Available online: https://www.iaea.org/sites/default/files/2020-guidelines-for-irradiation.pdf (accessed on 14 January 2023).
- Taze, Y.; Cuisance, D.; Politzar, H.; Clair, M.; Sellin, E. Essais de Determination de La Dose Optimale d’irradiation Des Males de Glossina Palpalis Gambiensis (Vanderplank, 1949) En Vue de La Lutte Biologique Par Lachers de Males Steriles Dans La Region de Bobo Dioulasso (Haute Volta). Rev. Elev. Med. Vet. Pays Trop. 1977, 30, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Ilboudo, K.; Camara, K.; Salou, E.W.; Gimonneau, G. Quality Control and Mating Performance of Irradiated Glossina Palpalis Gambiensis Males. Insects 2022, 13, 476. [Google Scholar] [CrossRef]
- De Beer, C.J.; Moyaba, P.; Boikanyo, S.N.B.; Majatladi, D.; Yamada, H.; Venter, G.J.; Vreysen, M.J.B. Evaluation of Radiation Sensitivity and Mating Performance of Glossina Brevipalpis Males. PLoS Negl. Trop. Dis. 2017, 11, e0005473. [Google Scholar] [CrossRef] [Green Version]
- Van der Vloedt, A.M.V.; Taher, M. Effect of Gamma Radiation on the Tsetse Fly Glossina Palpalis Palpalis (Rob.-Desv.) (Diptera:Glossinidae) with Observations on the Reproductive Biology. Int. J. Appl. Radiat. Isot. 1978, 29, 713–716. [Google Scholar] [CrossRef]
- Helinski, M.E.H.; Parker, A.G.; Knols, B.G. Radiation-Induced Sterility for Pupal and Adult Stages of the Malaria Mosquito Anopheles arabiensis. Malar. J. 2006, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Maiga, H.; Bimbile-Somda, N.S.; Carvalho, D.O.; Mamai, W.; Kraupa, C.; Parker, A.G.; Abrahim, A.; Weltin, G.; Wallner, T.; et al. The Role of Oxygen Depletion and Subsequent Radioprotective Effects during Irradiation of Mosquito Pupae in Water. Parasites Vectors 2020, 13, 198. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Maiga, H.; Kraupa, C.; Mamai, W.; Bimbilé Somda, N.S.; Abrahim, A.; Wallner, T.; Bouyer, J. Effects of Chilling and Anoxia on the Irradiation Dose-Response in Adult Aedes Mosquitoes. Front. Bioeng. Biotechnol. 2022, 10, 856780. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Maiga, H.; Juarez, J.; De Oliveira Carvalho, D.; Mamai, W.; Ali, A.; Bimbile-Somda, N.S.; Parker, A.G.; Zhang, D.; Bouyer, J. Identification of Critical Factors That Significantly Affect the Dose-Response in Mosquitoes Irradiated as Pupae. Parasites Vectors 2019, 12, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, V.; Shetty, N.J.; Ananthanarayana, S.R.; Jha, S.K.; Chaubey, R.C. Evaluation of Gamma Radiation-Induced DNA Damage in Aedes aegypti Using the Comet Assay. Toxicol. Ind. Health 2017, 33, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Pagabeleguem, S.; Seck, M.T.; Sall, B.; Vreysen, M.J.B.; Gimonneau, G.; Fall, A.G.; Bassene, M.; Sidibé, I.; Rayaissé, J.B.; Belem, A.M.G.; et al. Long Distance Transport of Irradiated Male Glossina Palpalis Gambiensis Pupae and Its Impact on Sterile Male Yield. Parasites Vectors 2015, 8, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, U.; Luger, D.; Barnor, H.; Dengwat, L.; Ajagbonna, B.; Vreysen, M.J.B.; Van der Vloedt, A.M.V. Tsetse Fly Mass Rearing: Colony Management, Deployment of Sterile Flies, Related Research and Development. In Tsetse Control, Diagnosis and Chemotherapy Using Nuclear Techniques, Proceedings of the Seminar, Jointly Organized by the IAEA and Fao, Muguga, Kenya, 11–15 February 1991; IAEA-TECDOC-634; IAEA: Vienna, Austria, 1992; pp. 167–180. [Google Scholar]
- Seck, M.T.; Pagabeleguem, S.; Bassene, M.D.; Fall, A.G.; Diouf, T.A.R.; Sall, B.; Vreysen, M.J.B.; Rayaissé, J.B.; Takac, P.; Sidibé, I.; et al. Quality of Sterile Male Tsetse after Long Distance Transport as Chilled, Irradiated Pupae. PLoS Negl. Trop. Dis. 2015, 9, e0004229. [Google Scholar] [CrossRef] [Green Version]
- Vreysen, M.J.B.; Saleh, K.M.; Ali, M.Y.; Abdulla, A.M.; Zhu, Z.-R.; Juma, K.G.; Dyck, V.A.; Msangi, A.R.; Mkonyi, P.A.; Feldmann, H.U. Glossina Austeni (Diptera: Glossinidae) Eradicated on the Island of Unguja, Zanzibar, Using the Sterile Insect Technique. J. Econ. Entomol. 2000, 93, 123–135. [Google Scholar] [CrossRef]
- Culbert, N.J.; Lees, R.S.; Vreysen, M.J.; Darby, A.C.; Gilles, J.R. Optimised Conditions for Handling and Transport of Male Anopheles arabiensis: Effects of Low Temperature, Compaction, and Ventilation on Male Quality. Entomol. Exp. Appl. 2017, 164, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Culbert, N.J.; Gilles, J.R.L. Investigating the Impact of Chilling Temperature on Male Aedes aegypti and Aedes albopictus Survival. PLoS ONE 2019, 14, e0221822. [Google Scholar] [CrossRef]
- IAEA Denial of Shipment. NS Update 2007, 5, 1–2. Available online: https://www-pub.iaea.org/MTCD/Publications/PDF/Newsletters/NSU-05.pdf (accessed on 15 December 2022).
- IAEA Denial of Shipment of Radioactive Material. NS Update 2010, 14, 2–4. Available online: https://www-pub.iaea.org/MTCD/Publications/PDF/Newsletters/NSU-14.pdf (accessed on 3 January 2023).
- Gray, P. Denial of Shipment of Radioactive Material. Packag. Transp. Storage Secur. Radioact. Mater. 2011, 22, 72–77. [Google Scholar] [CrossRef]
- De Wright, T.; Gray, P.; Sobriera, A.; Xavier, C.; Schwela, U. Delay and Denial of Shipment. In Proceedings of the International Conference on the Safe and Secure Transport of Radioactive Material: The Next Fifty Years, Vienna, Austria, 17–21 October 2011; IAEA-TECDOC-CD-1792. IAEA: Vienna, Austria, 2016; p. 7. [Google Scholar]
- Reiche, I. Technical Meeting (Virtual): Denial of Shipment—Issues and Solutions, Chairman’s Report of Federal Office for the Safety of Nuclear Waste Management (BASE), 23–26 March 2021. Available online: https://na.eventscloud.com/file_uploads/cf7798600527b2268b3c17ecd27a28ab_TecnicalMeetingEVT1907111DenialofShipment-Chairmansreport.pdf (accessed on 3 January 2023).





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamada, H.; Kaboré, B.A.; Bimbilé Somda, N.S.; Ntoyi, N.L.; de Beer, C.J.; Bouyer, J.; Caceres, C.; Mach, R.L.; Gómez-Simuta, Y. Suitability of Raycell MK2 Blood X-ray Irradiator for the Use in the Sterile Insect Technique: Dose Response in Fruit Flies, Tsetse Flies and Mosquitoes. Insects 2023, 14, 92. https://doi.org/10.3390/insects14010092
Yamada H, Kaboré BA, Bimbilé Somda NS, Ntoyi NL, de Beer CJ, Bouyer J, Caceres C, Mach RL, Gómez-Simuta Y. Suitability of Raycell MK2 Blood X-ray Irradiator for the Use in the Sterile Insect Technique: Dose Response in Fruit Flies, Tsetse Flies and Mosquitoes. Insects. 2023; 14(1):92. https://doi.org/10.3390/insects14010092
Chicago/Turabian StyleYamada, Hanano, Bénéwendé Aristide Kaboré, Nanwintoum Séverin Bimbilé Somda, Nonhlanhla L. Ntoyi, Chantel Janet de Beer, Jérémy Bouyer, Carlos Caceres, Robert L. Mach, and Yeudiel Gómez-Simuta. 2023. "Suitability of Raycell MK2 Blood X-ray Irradiator for the Use in the Sterile Insect Technique: Dose Response in Fruit Flies, Tsetse Flies and Mosquitoes" Insects 14, no. 1: 92. https://doi.org/10.3390/insects14010092
APA StyleYamada, H., Kaboré, B. A., Bimbilé Somda, N. S., Ntoyi, N. L., de Beer, C. J., Bouyer, J., Caceres, C., Mach, R. L., & Gómez-Simuta, Y. (2023). Suitability of Raycell MK2 Blood X-ray Irradiator for the Use in the Sterile Insect Technique: Dose Response in Fruit Flies, Tsetse Flies and Mosquitoes. Insects, 14(1), 92. https://doi.org/10.3390/insects14010092