
Exploring Cold Hardiness within a Butterfly Clade: Supercooling Ability and Polyol Profiles in European Satyrinae


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
- What are the supercooling abilities and concentrations of putatively cryoprotectant compounds in the studied species, do these measures of cold hardiness correlate with each other, and where do the studied species stand relative to the high-elevation Erebia spp.?
- How do the above characteristics of cold hardiness change from late autumn to winter?
- Which of the sugars and polyols, based on correlations between their concentration and cold hardiness measures, play a cryoprotective role in the studied species?
- Is there a relationship among the Satyrinae cold hardiness characteristics and their vertical distribution (the latter standing for the harshness of conditions experienced by overwintering larvae)?
- Is there a phylogenetic signal of the identity of cryoprotectant compounds used?
2. Materials and Methods
2.1. Study Species
2.2. Captive Rearing
2.3. Supercooling Point and Polyol Profiles
2.4. Statistical Analyses
2.5. Phylogeny of Sugar and Polyol Profiles
2.6. Sugar and Polyol Profiles versus Cold Hardiness
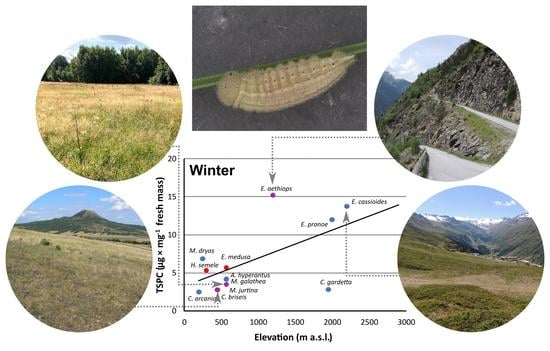
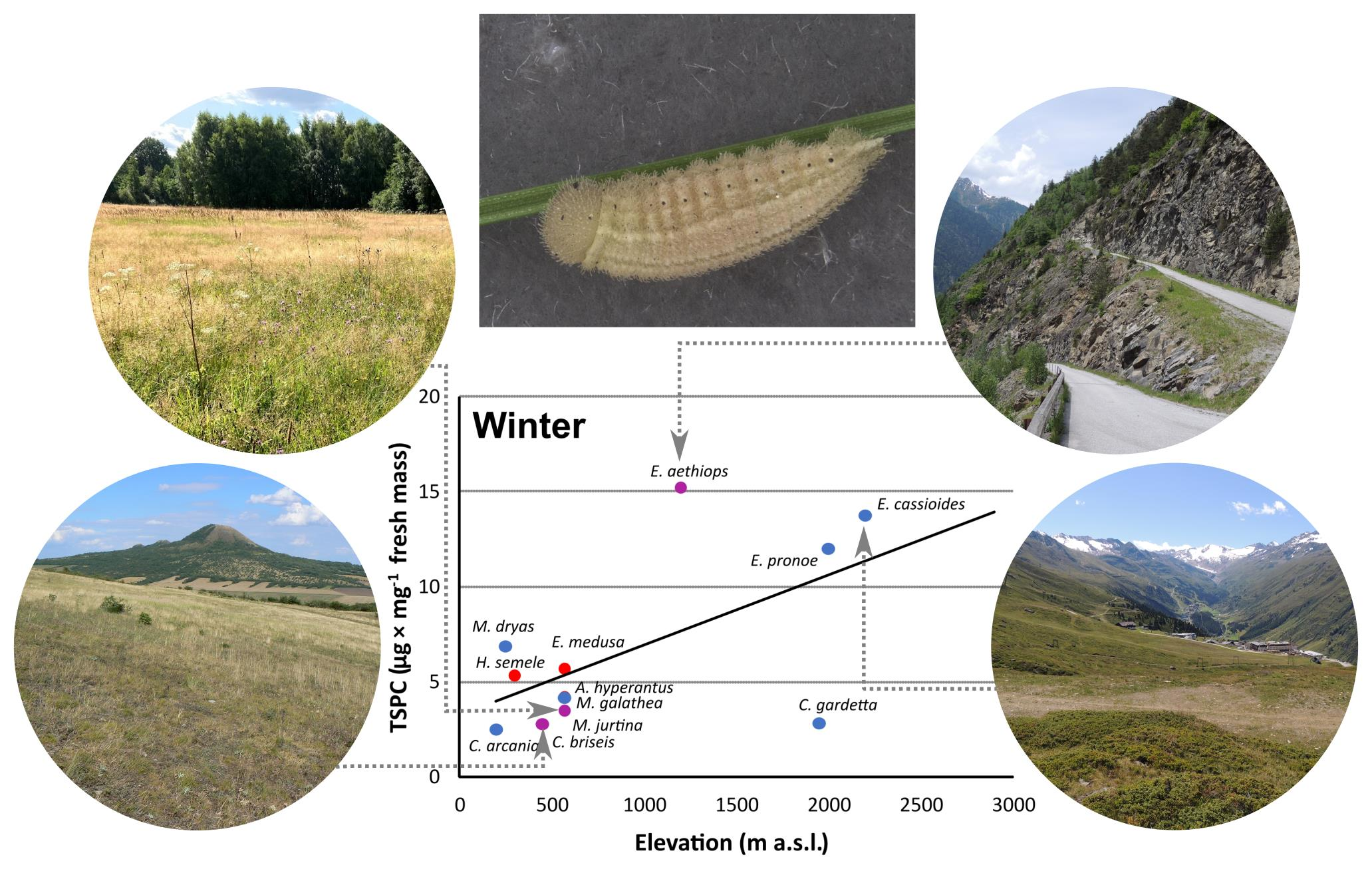
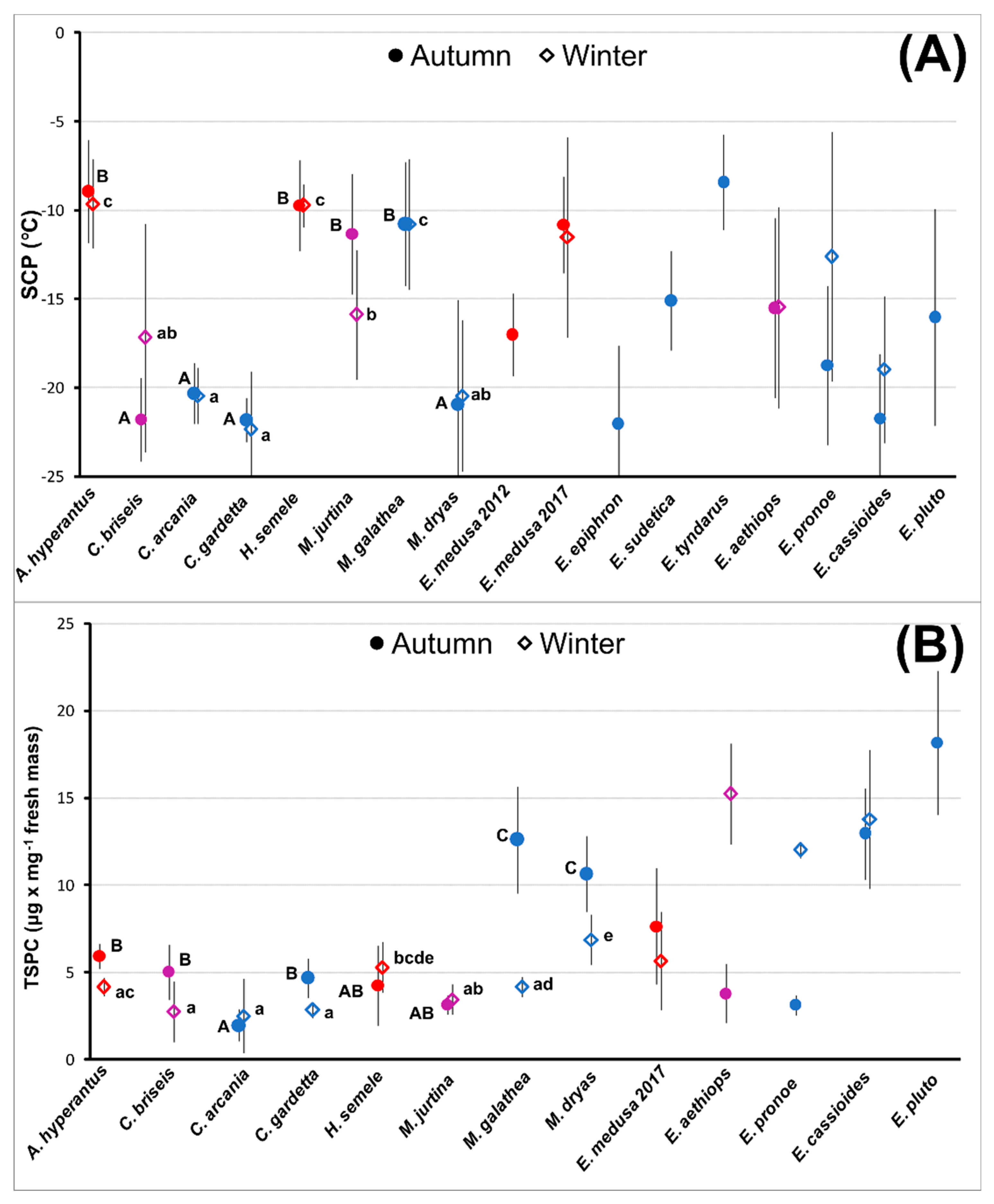
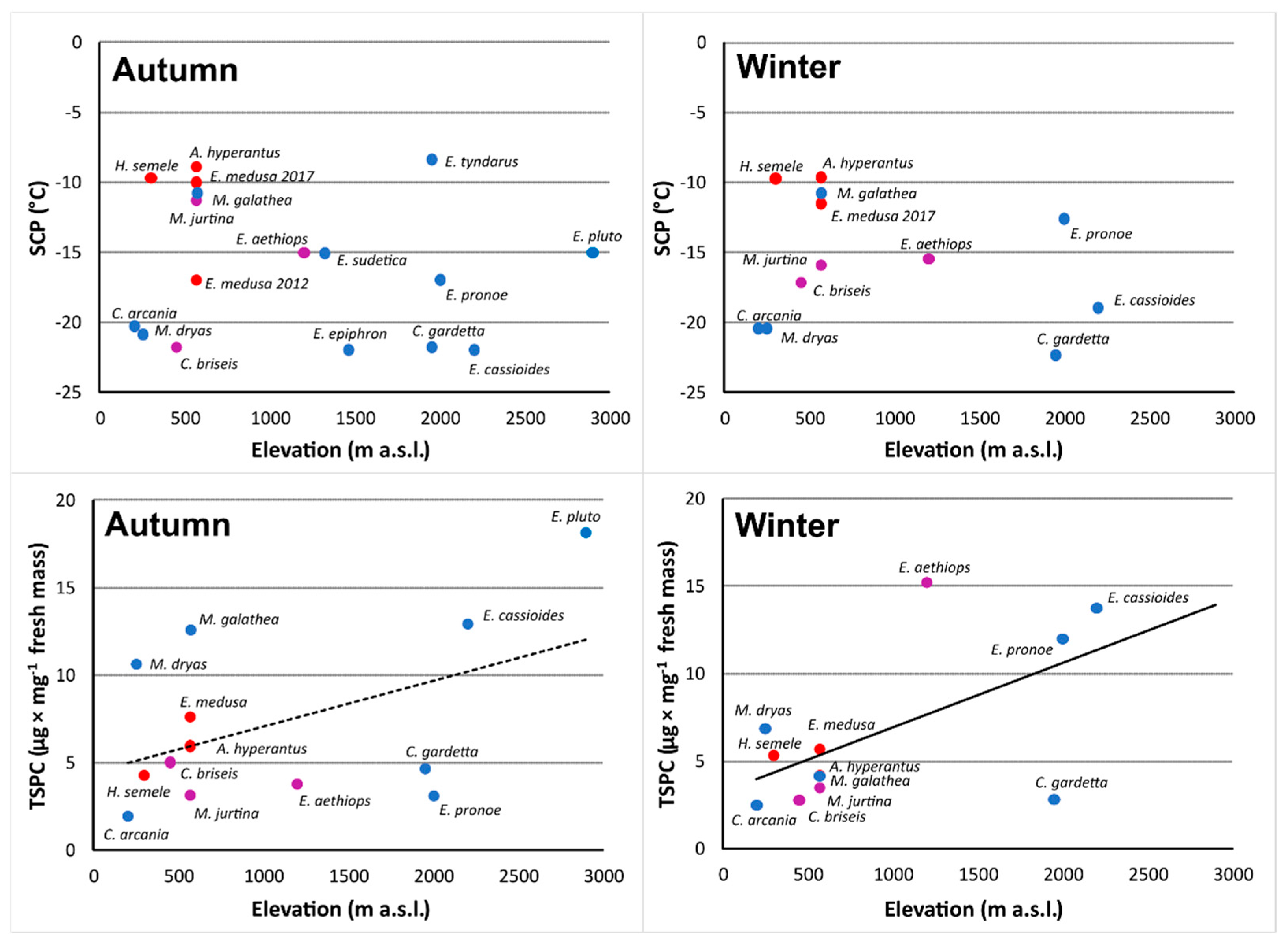
3. Results
3.1. Supercooling Ability and Sugar and Polyol Concentrations
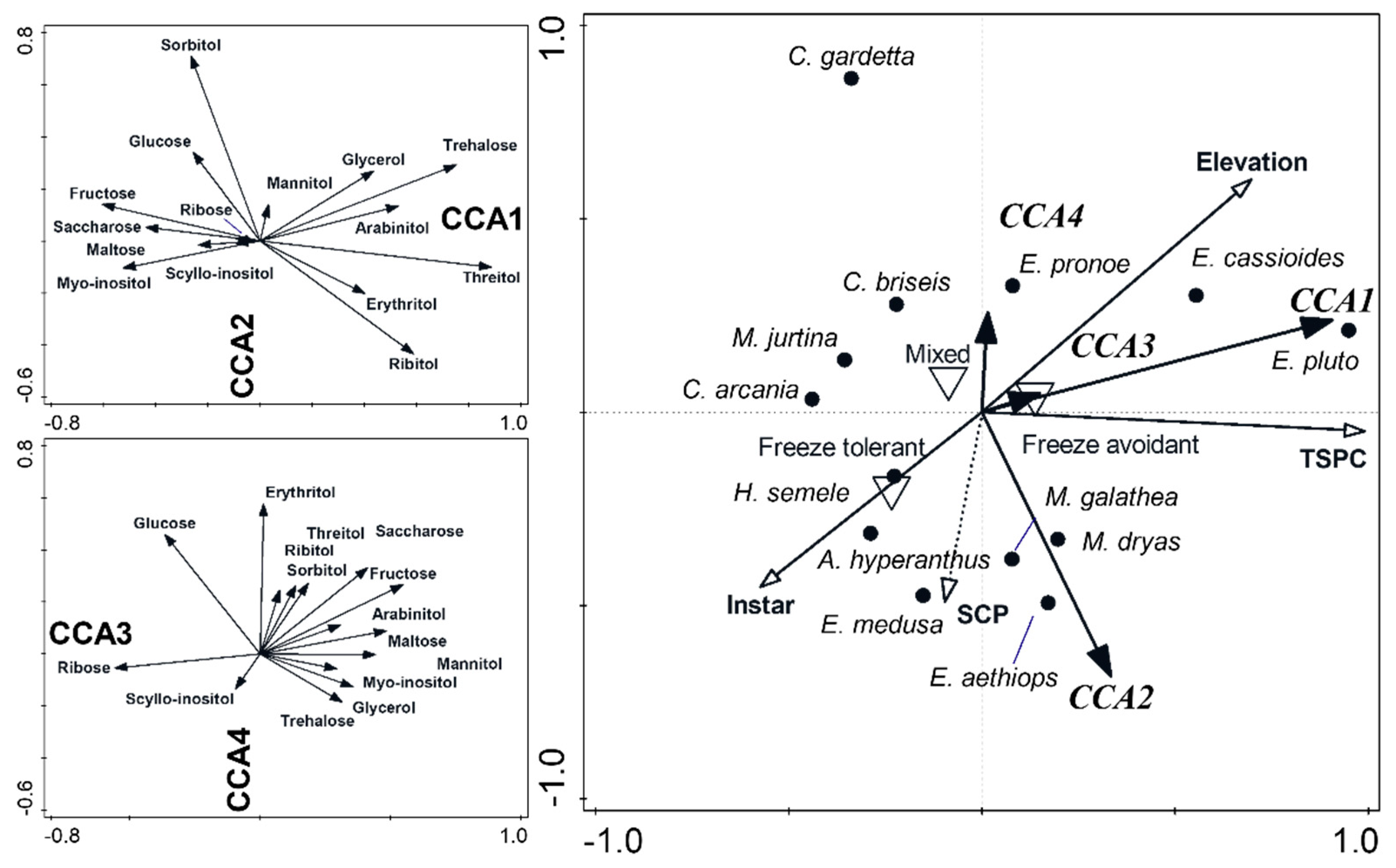
3.2. Sugar and Polyol Profiles
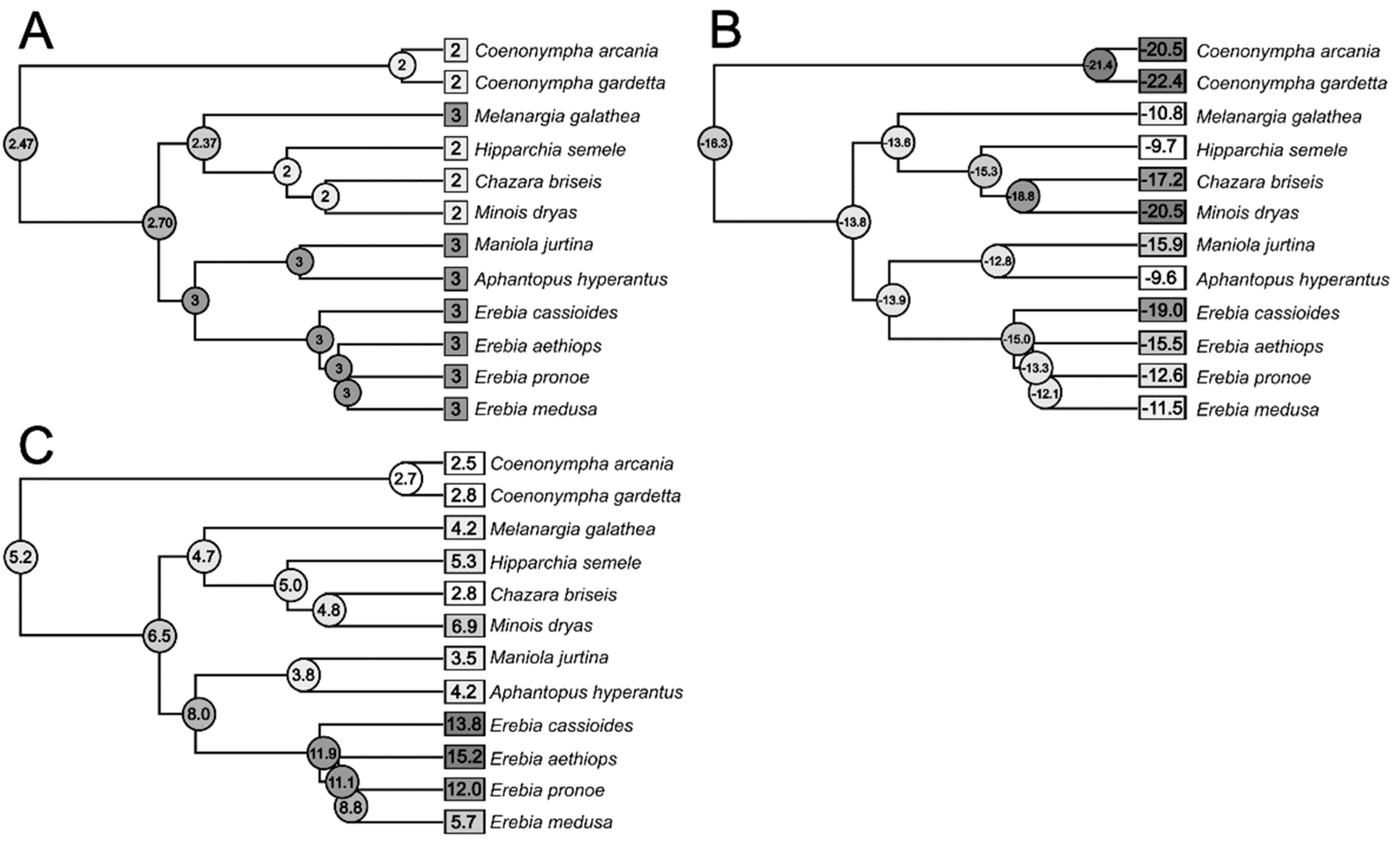
3.3. Phylogenetic Signal and Ancestral State Reconstruction
3.4. Relations of Cold Hardiness Traits to Sugars and Polyol Profiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
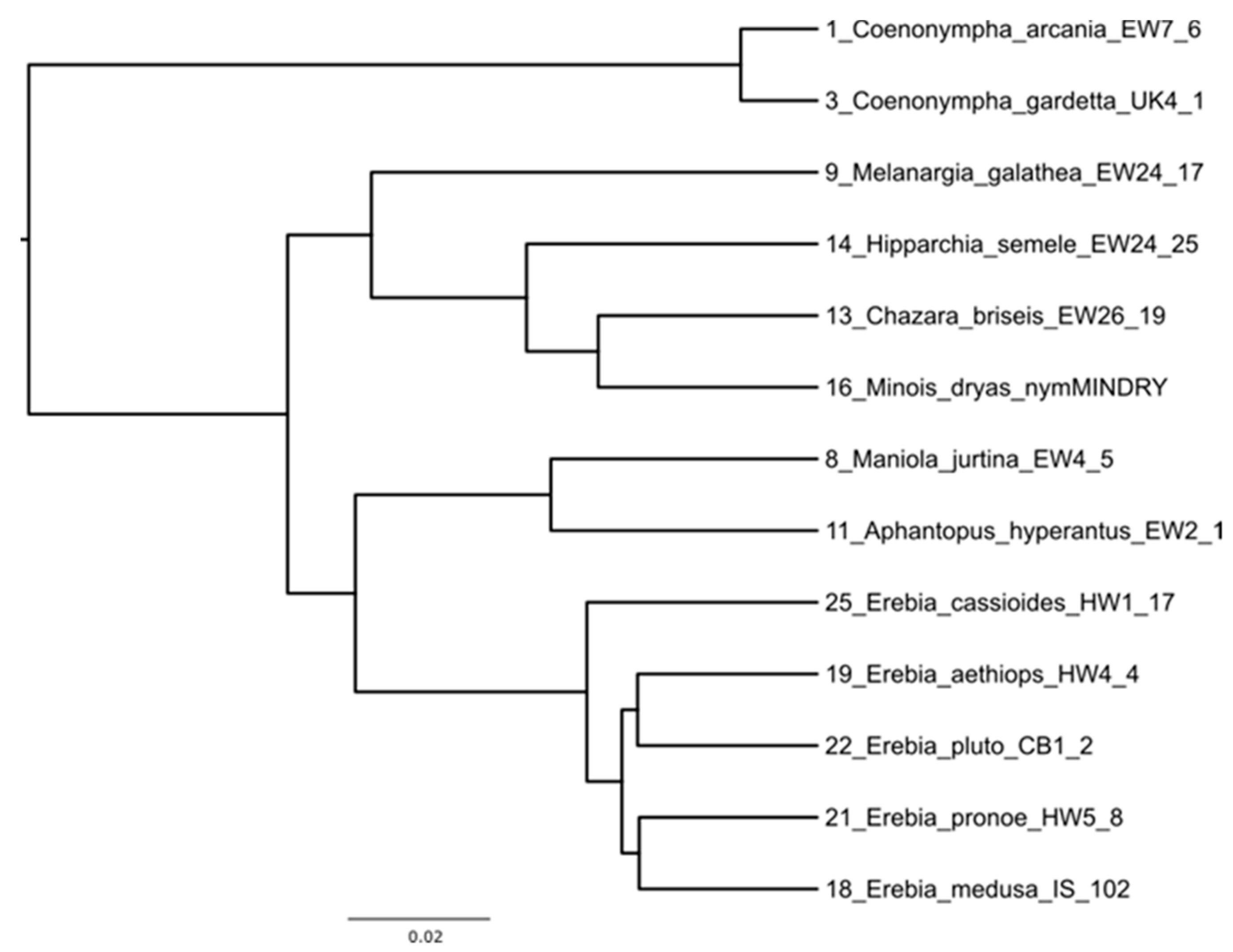
Appendix A. Reconstruction of Phylogenetic Tree Used for Phylogeny of Sugar and Polyol Profiles in Satyrinae Butterflies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Voucher | Species | GenBank Codes | ||||
|---|---|---|---|---|---|---|
| COI | EF-1 Alpha | Wingless | GAPDH | RpS5 | ||
| EW7-6 | Coenonympha arcania | EU920744 | EU920777 | EU920807 | ||
| EW18-25 | Coenonympha arcania | EU920769 | EU920801 | EU920827 | ||
| UK4-1 | Coenonympha gardetta | EU920747 | EU920781 | EU920811 | ||
| EW5-18 | Coenonympha glycerion | EU920749 | EU920783 | EU920812 | GQ892089 | GQ892095 |
| M66 | Maniola jurtina | KP032298 | KP032635 | KP032497 | ||
| M40 | Maniola jurtina | KP032276 | KP032628 | KP032488 | ||
| M69 | Maniola jurtina | KP032300 | KP032636 | KP032499 | ||
| EW4-5 | Maniola jurtina | AY090214 | AY090180 | KR139116 | EU141481 | EU141376 |
| EW24-17 | Melanargia galathea | DQ338843 | DQ338993 | DQ338706 | EU528398 | EU528444 |
| DNAwthmel 190 | Melanargia galathea | GQ200938 | GQ201380 | |||
| EW2-1 | Aphantopus hyperantus | AY090211 | AY090177 | AY090144 | GQ357458 | GQ357588 |
| MM00045 | Aphantopus hyperantus | KT713830, KT713873 | KT713921 | JN204924 | ||
| EW26-19 | Chazara briseis | DQ338866 | DQ339021 | DQ338730 | GQ357502 | GQ357628 |
| EW24-25 | Hipparchia semele | DQ338868 | DQ339023 | DQ338732 | ||
| midry1 | Minois dryas | KM111645 | KJ805881 | |||
| nymMINDRY | Minois dryas | JX185830 | JX185940 | |||
| JM10-8 | Erebia medusa | KR138851 | KR231870 | KR138936 | ||
| IS-102 | Erebia medusa | KR138784 | KR139139 | KR138914 | ||
| HW4-4 | Erebia aethiops | KR138772 | KR139132 | KR139021 | KR138907 | |
| HW2-15 | Erebia aethiops | KR138768 | KR139018 | KR138903 | ||
| HW5-8 | Erebia pronoe | KR138780 | KR139138 | KR139026 | KR138912 | |
| CB1-2 | Erebia pluto | KR138846 | KR139089 | KR138981 | KR138862 | |
| CB1-4 | Erebia pluto | KR138847 | KR139092 | KR138984 | KR138865 | |
| LC340526 | Erebia pluto | LC340526 | ||||
| HW1-17 | Erebia cassioides | KR138758 | KR139120 | KR139010 | KR138892 | |
| HW1-20 | Erebia cassioides | KR138759 | KR139121 | KR139011 | KR138893 | |
| NW147-3 | Charaxes jasius | GQ256824 | GQ256950 | GQ256703 | GQ257152 | |
| NW69-6 | Apatura iris | AY090199 | AY090165 | AY090132 | EU141513 | EU141692 |
| TS12 | Brenthis daphne | DQ922848 | DQ922880 | DQ922816 | KY773429 | KY773533 |
| EW1-1 | Pararge aegeria | DQ176379 | DQ338913 | DQ338620 | EU141476 | EU141372 |
| CP15-04 | Dira clytus | EU528319 | EU528296 | EU528275 | EU528388 | EU528432 |
| CP-AC23-35 | Paralasa jordana | DQ338597 | DQ339027 | EU532176 | EU528455 | |
| CP06-89 | Oressinoma sorata | GQ357209 | GQ357278 | GQ357342 | GQ357440 | GQ357570 |
| NW136-1 | Argyronympha gracilipes | DQ338816 | GQ357263 | DQ338676 | GQ357402 | GQ357532 |
| D30 | Zipaetis saitis | DQ338831 | DQ338981 | DQ338696 | EU528418 | EU528472 |
| NW144-2 | Cassionympha cassius | GQ357213 | GQ357282 | GQ357346 | GQ357450 | GQ357579 |
| JM9-3 | Erebia callias | KR138824 | KR139159 | KR139062 | KR138954 | |
| JM8-15 | Erebia melampus | KR138818 | KR231871 | KR139058 | KR138948 | |
| JL5-12 | Erebia epiphron | KR138798 | KR231861 | KR139042 | KR138927 | |
| HW1-22 | Erebia triaria | KR138760 | KR139122 | KR139012 | KR138894 | |
| IS-65 | Erebia styx | KR138797 | KR139154 | KR139040 | KR138925 | |
| CB11-5 | Erebia lefebvrei | KR138717 | KR139088 | KR138980 | KR138861 | |
| JM10-14 | Erebia neoridas | KR138801 | KR139156 | KR139044 | KR138930 | |
| EW25-31 | Erebia ligea | KR138753 | KR139115 | KR138890 | ||
| JM9-2 | Erebia pandrose | KR138823 | KR231880 | KR138953 | ||
| CB7-1 | Erebia eriphyle | KR138738 | KR139108 | KR139000 | KR138881 | |
| CB3-11 | Erebia gorge | KR138720 | KR139094 | KR138986 | KR138867 | |


References
- Sauer, J.; Domisch, S.; Nowak, C.; Haase, P. Low mountain ranges: Summit traps for montane freshwater species under climate change. Biodivers. Conserv. 2011, 20, 3133–3146. [Google Scholar] [CrossRef]
- McCain, C.M.; Garfinkel, C.F. Climate change and elevational range shifts in insects. Curr. Opin. Insect Sci. 2021, 47, 111–118. [Google Scholar]
- Sistri, G.; Menchetti, M.; Santini, L.; Pasquali, L.; Sapienti, S.; Cini, A.; Platania, L.; Balletto, E.; Barbero, F.; Bonelli, S.; et al. The isolated Erebia pandrose Apennine population is genetically unique and endangered by climate change. Insect Conserv. Divers. 2022, 15, 136–148. [Google Scholar] [CrossRef]
- Matter, S.F.; Doyle, A.; Illerbrun, K.; Wheeler, J.; Roland, J. An assessment of direct and indirect effects of climate change for populations of the Rocky Mountain Apollo butterfly (Parnassius smintheus Doubleday). Insect Sci. 2011, 18, 385–392. [Google Scholar] [CrossRef]
- Abarca, M.; Larsen, E.A.; Ries, L. Heatwaves and Novel Host Consumption Increase Overwinter Mortality of an Imperiled Wetland Butterfly. Front. Ecol. Evol. 2019, 7, 193. [Google Scholar] [CrossRef] [Green Version]
- Roland, J.; Filazzola, A.; Matter, S.F. Spatial variation in early-winter snow cover determines local dynamics in a network of alpine butterfly populations. Ecography 2020, 44, 334–343. [Google Scholar] [CrossRef]
- Konvicka, M.; Kuras, T.; Liparova, J.; Slezak, V.; Horazna, D.; Klecka, J.; Kleckova, I. Low winter precipitation, but not warm autumns and springs, threatens mountain butterflies in middle-high mountains. PeerJ 2021, 9, 12021. [Google Scholar] [CrossRef]
- Klockmann, M.; Fischer, K. Strong reduction in diapause survival under warm and humid overwintering conditions in a temperate-zone butterfly. Popul. Ecol. 2019, 61, 150–159. [Google Scholar] [CrossRef]
- Kuras, T.; Benes, J.; Fric, Z.; Konvicka, M. Dispersal patterns of endemic alpine butterflies with contrasting population structures: Erebia epiphron and E. sudetica. Popul. Ecol. 2003, 45, 115–123. [Google Scholar] [CrossRef]
- Sonderegger, P. Die Erebien der Schweiz (Lepidoptera: Satyrinae, Genus Erebia); Verlag Peter Sonderegger: Biel/Bienne, Switzerland, 2005; 712p. [Google Scholar]
- Schmitt, T.; Habel, J.C.; Rodder, D.; Louy, D. Effects of recent and past climatic shifts on the genetic structure of the high mountain Yellow-spotted ringlet butterfly Erebia manto (Lepidoptera, Satyrinae): A conservation problem. Global Change Biol. 2014, 20, 2045–2061. [Google Scholar] [CrossRef]
- Pena, C.; Witthauer, H.; Kleckova, I.; Fric, Z.; Wahlberg, N. Adaptive radiations in butterflies: Evolutionary history of the genus Erebia (Nymphalidae: Satyrinae). Biol. J. Linn. Soc. 2015, 116, 449–467. [Google Scholar] [CrossRef] [Green Version]
- Ehl, S.; Dalstein, V.; Tull, F.; Gros, P.; Schmitt, T. Specialized or opportunistic-how does the high mountain endemic butterfly Erebia nivalis survive in its extreme habitats? Insect Sci. 2018, 25, 161–171. [Google Scholar] [PubMed]
- Vrba, P.; Konvicka, M.; Nedved, O. Reverse altitudinal cline in cold hardiness among Erebia butterflies. CryoLetters 2012, 33, 251–258. [Google Scholar] [PubMed]
- Vrba, P.; Nedved, O.; Zahradnickova, H.; Konvicka, M. More complex than expected: Cold hardiness and the concentration of cryoprotectants in overwintering larvae of five Erebia butterflies (Lepidoptera: Nymphalidae). Eur. J. Entomol. 2017, 114, 470–480. [Google Scholar] [CrossRef]
- Williams, C.M.; Nicolai, A.; Ferguson, L.V.; Bernards, M.A.; Hellmann, J.J.; Sinclair, B.J. Cold hardiness and deacclimation of overwintering Papilio zelicaon pupae. Comp. Biochem. Physiol. A 2014, 178, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chazot, N.; Condamine, F.L.; Dudas, G.; Pena, C.; Kodandaramaiah, U.; Matos-Maravi, P.; Aduse-Poku, K.; Elias, M.; Warren, A.D.; Lohman, D.J.; et al. Conserved ancestral tropical niche but different continental histories explain the latitudinal diversity gradient in brush-footed butterflies. Nat. Comm. 2021, 12, 5717. [Google Scholar] [CrossRef]
- Kleckova, I.; Konvicka, M.; Klecka, J. Thermoregulation and microhabitat use in mountain butterflies of the genus Erebia: Importance of fine-scale habitat heterogeneity. J. Therm. Biol. 2014, 41, 50–58. [Google Scholar] [CrossRef]
- Zachariassen, K.E. Physiology of cold tolerance in insects. Physiol. Rev. 1985, 65, 799–832. [Google Scholar] [CrossRef]
- Ramlov, H.; Bedford, J.; Leader, J. Freezing tolerance of the New Zealand weta, Hemideina maori Hutton (Orthoptera, Stenopelmatidae). J. Therm. Biol. 1992, 17, 51–54. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Vernon, P.; Klok, C.J.; Chown, S.L. Insects at low temperatures: An ecological perspective. Trends Ecol. Evol. 2003, 18, 257–262. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Alvarado, L.E.C.; Ferguson, L.V. An invitation to measure insect cold tolerance: Methods, approaches, and workflow. J. Therm. Biol. 2015, 53, 180–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedved, O. Modelling the relationship between cold injury and accumulated degree days in terrestrial arthropods. Cryo-Lett. 1998, 19, 267–274. [Google Scholar]
- Vrba, P.; Nedved, O.; Konvicka, M. Contrasting Supercooling Ability in Lowland and Mountain European Colias Butterflies. J. Entomol. Sci. 2014, 49, 63–69. [Google Scholar] [CrossRef]
- Somme, L. Supercooling and winter survival in terrestrial arthropods. Comp. Biochem. Physiol. A 1982, 73, 519–543. [Google Scholar] [CrossRef]
- Ramlov, H. Aspects of natural cold tolerance in ectothermic animals. Hum. Reprod. 2000, 15, 26–46. [Google Scholar] [CrossRef]
- Kostal, V.; Slachta, M.; Simek, P. Cryoprotective role of polyols independent of the increase in supercooling capacity in diapausing adults of Pyrrhocotis apterus (Heteroptera: Insecta). Comp. Biochem. Phys. B 2001, 130, 365–374. [Google Scholar] [CrossRef]
- Kadlec, T.; Vrba, P.; Kepka, P.; Schmitt, T.; Konvicka, M. Tracking the decline of the once-common butterfly: Delayed oviposition, demography and population genetics in the hermit Chazara briseis. Anim. Conserv. 2010, 13, 172–183. [Google Scholar] [CrossRef]
- Suchackova Bartonova, A.; Konvicka, M.; Maresova, J.; Blahova, D.; Cip, D.; Skala, P.; Andres, M.; Hula, V.; Dolek, M.; Geyer, A.; et al. Extremely Endangered Butterflies of Scattered Central European Dry Grasslands Under Current Habitat Alteration. Insect Syst. Divers. 2021, 5, 6. [Google Scholar] [CrossRef]
- Kadlec, T.; Vrba, P. A second generation of Coenonympha arcania (Linnaeus, 1761) in northern Bohemia, Czech Republic (Lepidoptera, Satyrinae). Atalanta 2007, 38, 181–183. [Google Scholar]
- Kodandaramaiah, U.; Wahlberg, N. Phylogeny and biogeography of Coenonympha butterflies (Nymphalidae: Satyrinae)—patterns of colonization in the Holarctic. Syst. Entomol. 2009, 34, 315–323. [Google Scholar] [CrossRef]
- Wiemers, M.; Schweiger, O.; Harpke, A.; Settele, J. Climatic niche characteristics of the butterflies in Europe (CLIMBER)—A new database for measuring the response of butterflies to climatic changes. Zitteliana 2016, 88, 53. [Google Scholar]
- Tropek, R.; Cizek, O.; Kadlec, T.; Klecka, J. Habitat use of Hipparchia semele (Lepidoptera) in its artificial stronghold: Necessity of the resource-based habitat view in restoration of disturbed sites. Pol. J. Ecol. 2017, 65, 385–399. [Google Scholar]
- Kalarus, K.; Nowicki, P. How Do Landscape Structure, Management and Habitat Quality Drive the Colonization of Habitat Patches by the Dryad Butterfly (Lepidoptera: Satyrinae) in Fragmented Grassland? PLoS ONE 2015, 10, e0138557. [Google Scholar] [CrossRef] [PubMed]
- Hanson, A.A.; Venette, R.C. Thermocouple design for measuring temperatures of small insects. CryoLetters 2013, 34, 261–266. [Google Scholar] [PubMed]
- Brunnhofer, V.; Nedved, O.; Hodkova, M. Methodical improvement for measuring of supercooling point in insects. Acta Entomol. Bohemoslov. 1991, 88, 349–350. [Google Scholar]
- Kostal, V.; Zahradnickova, H.; Simek, P.; Zeleny, J. Multiple component system of sugars and polyols in the overwintering spruce bark beetle, Ips typographus. J. Insect Physiol. 2007, 53, 580–586. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. Canoco 5, Windows Release (5.00). 2013. Available online: http://www.canoco5.com/ (accessed on 1 September 2019).
- Blomberg, S.P.; Garland, T.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [CrossRef]
- Revell, L.J. Phytools: Phylogenetic tools for comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating behavior to habitat: Solutions to the fourth-corner problem. Ecology 1997, 78, 547–562. [Google Scholar] [CrossRef]
- Pena, C.; Nylin, S.; Freitas, A.V.L. Higher level phylogeny of Satyrinae butterflies (Lepidoptera: Nymphalidae) based on DNA sequence data. Mol. Phylogenet. Evol. 2006, 40, 29–49. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.R., Jr.; Pan, Q.; Serianni, A.S.; Duman, J.G. Cryoprotectant Biosynthesis and the Selective Accumulation of Threitol in the Freeze-tolerant Alaskan Beetle, Upis ceramboides. J. Biol. Chem. 2009, 284, 16822–16831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Merwe, M.; Chown, S.L.; Smith, V.R. Thermal tolerance limits in six weevil species (Coleoptera, Curculionidae) from sub-Antarctic Marion Island. Polar Biol. 1997, 18, 331–336. [Google Scholar] [CrossRef]
- Vernon, P.; Vannier, G. Freezing susceptibility and freezing tolerance in Palaearctic Cetoniidae (Coleoptera). Can. J. Zool. 2001, 79, 67–74. [Google Scholar] [CrossRef]
- Ditrich, T.; Kostal, V. Comparative analysis of overwintering physiology in nine species of semi-aquatic bugs (Heteroptera: Gerromorpha). Physiol. Entomol. 2011, 36, 261–270. [Google Scholar] [CrossRef]
- Strachan, L.A.; Tarnowski-Garner, H.E.; Marshall, K.E.; Sinclair, B.J. The Evolution of Cold Tolerance in Drosophila Larvae. Physiol. Biochem. Zool. 2011, 84, 43–53. [Google Scholar] [CrossRef]
- Andersen, J.L.; Manenti, T.; Sorensen, J.G.; MacMillan, H.A.; Loeschcke, V.; Overgaard, J. How to assess Drosophila cold tolerance: Chill coma temperature and lower lethal temperature are the best predictors of cold distribution limits. Funct. Ecol. 2015, 29, 55–65. [Google Scholar] [CrossRef]
- Dennis, A.B.; Dunning, L.T.; Sinclair, B.J.; Buckley, T.R. Parallel molecular routes to cold adaptation in eight genera of New Zealand stick insects. Sci. Rep. 2015, 5, 13965. [Google Scholar] [CrossRef] [Green Version]
- Albre, J.; Gers, C.; Legal, L. Molecular phylogeny of the Erebia tyndarus (Lepidoptera, Rhopalocera, Nymphalidae, Satyrinae) species group combining CoxII and ND5 mitochondrial genes: A case study of a recent radiation. Mol. Phylogenet. Evol. 2008, 47, 196–210. [Google Scholar] [CrossRef]
- Pena, C.; Nylin, S.; Wahlberg, N. The radiation of Satyrini butterflies (Nymphalidae: Satyrinae): A challenge for phylogenetic methods. Zool. J. Linn. Soc. 2011, 161, 64–87. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, B.J.; Addo-Bediako, A.; Chown, S.L. Climatic variability and the evolution of insect freeze tolerance. Biol. Rev. 2003, 78, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Zachariassen, K.E. The role of polyols and nucleating agents in cold hardy beetles. J. Comp. Physiol. 1980, 140, 227–234. [Google Scholar] [CrossRef]
- Wallis deVries, M.F.; Baxter, W.; Van Vliet, A.J.H. Beyond climate envelopes effects of weather on regional population trends in butterflies. Oecologia 2011, 167, 559–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, T.H.; Marshall, H.H.; Morecroft, M.D.; Brereton, T.; Prudhomme, C.; Huntingford, C. Interacting effects of climate change and habitat fragmentation on drought-sensitive butterflies. Nat. Clim. Change 2015, 5, 941–945. [Google Scholar] [CrossRef] [Green Version]
- Stuhldreher, G.; Hermann, G.; Fartmann, T. Cold-adapted species in a warming world—An explorative study on the impact of high winter temperatures on a continental butterfly. Entomol. Exp. Appl. 2014, 151, 270–279. [Google Scholar] [CrossRef]
- Slamova, I.; Klecka, J.; Konvicka, M. Diurnal Behavior and Habitat Preferences of Erebia aethiops, an Aberrant Lowland Species of a Mountain Butterfly Clade. J. Insect Behav. 2011, 24, 230–246. [Google Scholar] [CrossRef]
- Kulfan, J.; Strbova, E.; Zach, P. Effect of vegetation and management on occurrence of larvae and adults of generalist Maniola jurtina L. (Lepidoptera) in meadow habitats. Pol. J. Ecol. 2012, 60, 601–609. [Google Scholar]
- Villemey, A.; Peterman, W.E.; Richard, M.; Ouin, A.; van Halder, I.; Stevens, V.M.; Baguette, M.; Roche, P.; Archaux, F. Butterfly dispersal in farmland: A replicated landscape genetics study on the meadow brown butterfly (Maniola jurtina). Landsc. Ecol. 2016, 31, 1629–1641. [Google Scholar] [CrossRef]
- Li, N.G.; Toxopeus, J.; Moos, M.; Sorensen, J.G.; Sinclair, B.J. A comparison of low temperature biology of Pieris rapae from Ontario, Canada, and Yakutia, Far Eastern Russia. Comp. Biochem. Phys. A 2020, 242, 1106492. [Google Scholar] [CrossRef]
- Kock, R.A.; Orynbayev, M.; Robinson, S.; Zuther, S.; Singh, N.J.; Beauvais, W.; Morgan, E.R.; Kerimbayev, A.; Khomenko, S.; Martineau, H.M.; et al. Saigas on the brink: Multidisciplinary analysis of the factors influencing mass mortality events. Sci. Adv. 2018, 4, aao231. [Google Scholar] [CrossRef] [Green Version]
- Kleckova, I.; Klecka, J. Facing the Heat: Thermoregulation and Behaviour of Lowland Species of a Cold-Dwelling Butterfly Genus, Erebia. PLoS ONE 2016, 11, e0150393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullin, A.S.; Bale, J.S.; Fontaine, X.L.R. Physiological aspects of diapause and cold tolerance during overwintering in Pieris brassicae. Physiol. Entomol. 1991, 16, 447–456. [Google Scholar] [CrossRef]
- Lehmann, P.; Pruisscher, P.; Kostal, V.; Moos, M.; Simek, P.; Nylin, S.; Agren, R.; Varemo, L.; Wiklund, C.; Wheat, C.W.; et al. Metabolome dynamics of diapause in the butterfly Pieris napi: Distinguishing maintenance, termination and post-diapause phases. J. Exp. Biol. 2018, 221, jeb169508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullin, A.S.; Bale, J.S. Effects of low temperature on diapausing Aglais urticae and Inachis io (Lepidoptera, Nymphalidae)—overwintering physiology. J. Insect Physiol. 1989, 35, 283–290. [Google Scholar] [CrossRef]
- Vrba, P.; Dolek, M.; Nedved, O.; Zahradnickova, H.; Cerrato, C.; Konvicka, M. Overwintering of the boreal butterfly Colias palaeno in central Europe. CryoLetters 2014, 35, 247–254. [Google Scholar] [PubMed]
- Ahmadi, B.; Moharramipour, S.; Sinclair, B.J. Overwintering biology of the carob moth Apomyelois ceratoniae (Lepidoptera: Pyralidae). Int. J. Pest Manag. 2016, 62, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Han, E.N.; Bauce, E. Non-freeze survival of spruce budworm larvae, Choristoneura fumiferana, at subzero temperatures during diapause. Entomol. Exp. Appl. 1995, 75, 67–74. [Google Scholar] [CrossRef]
- Toxopeus, J.; Kostal, V.; Sinclair, B.J. Evidence for non-colligative function of small cryoprotectants in a freeze-tolerant insect. Proc. R. Soc. B Biol. 2019, 286, 20190050. [Google Scholar] [CrossRef] [Green Version]
- Vrba, P.; Nedved, O.; Zahradnickova, H.; Konvicka, M. Temporal plasticity in cold hardiness and cryoprotectant contents in northern versus temperate Colias butterflies (Lepidoptera: Pieridae). CryoLetters 2017, 38, 330–338. [Google Scholar]
- Li, J.W.; Shi, J.; Xue, Y.; Mao, H.B.; Luo, Y.Q. Major physiological adjustments in freezing-tolerant grey tiger longicorn beetle (Xylotrechus rusticus) during overwintering period. J. Forest. Res. 2014, 25, 653–659. [Google Scholar] [CrossRef]
- Hasanvand, H.; Izadi, H.; Mohammadzadeh, M. Overwintering Physiology and Cold Tolerance of the Sunn Pest, Eurygaster integriceps, an Emphasis on the Role of Cryoprotectants. Front. Physiol. 2020, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, S.; Li, Y.P.; Nakana, K.; Tsumuki, H.; Goto, M. Seasonal changes in glycerol content and cold hardiness in two ecotypes of the rice stem borer, Chilo suppressalis, exposed to the environment in the Shonai district, Japan. J. Insect Physiol. 2007, 53, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Kim, Y.; Park, G.W.; Lee, J.O.; Lee, K.W. Supercooling capacity along with up-regulation of glycerol content in an overwintering butterfly, Parnassius bremeri. J. Asia-Pac. Entomol. 2017, 20, 949–954. [Google Scholar] [CrossRef]
- Mohammadzadeh, M.; Izadi, H. Cold Acclimation of Trogoderma granarium Everts Is Tightly Linked to Regulation of Enzyme Activity, Energy Content, and Ion Concentration. Front. Physiol. 2018, 9, 1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostal, V.; Nedved, O.; Simek, P. Accumulation of high concentrations of myo-inositol in the overwintering ladybird beetle Ceratomegilla undecimnotata. Cryo-Lett. 1996, 17, 267–272. [Google Scholar]
- Yoshida, M.; Abe, J.; Moiyama, M.; Kuwabara, T. Carbohydrate levels among winter wheat cultivars varying in freezing tolerance and snow mold resistance during autumn and winter. Physiol. Plant. 1998, 103, 8–16. [Google Scholar] [CrossRef]
- Kostal, V.; Stetina, T.; Poupardin, R.; Korbelova, J.; Bruce, A.W. Conceptual framework of the ecophysiological phases of insect diapause development justified by transcriptomic profiling. Proc. Nat. Acad. Sci. USA 2017, 114, 8532–8537. [Google Scholar] [CrossRef] [Green Version]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef]
- Tamang, A.M.; Kalra, B.; Parkash, R. Cold and desiccation stress induced changes in the accumulation and utilization of proline and trehalose in seasonal populations of Drosophila immigrans. Comp. Biochem. Physiol. A 2017, 203, 304–313. [Google Scholar] [CrossRef]
- Guo, N.; Puhlev, I.; Brown, D.; Mansbridge, J.; Levine, F. Trehalose expression confers desiccation tolerance on human cells. Nat. Biotechnol. 2000, 18, 168–171. [Google Scholar] [CrossRef]
- Sinclair, B.J. Water relations of the freeze-tolerant New Zealand alpine cockroach Celatoblatta quinquemaculata (Dictyoptera: Blattidae). J. Insect Physiol. 2000, 46, 869–876. [Google Scholar] [CrossRef]
- Clements, R.S. The Polyol Pathway. Drugs 1986, 32, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2016, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]





| Species (Phenomena Studied) | Origin (CZ—Czechia, AT—Austria) | Elevation | n (SCP) * | n (LLt) | n (TSCP) * |
|---|---|---|---|---|---|
| This study (SCP, TSPC) | |||||
| Apanthopus hyperantus | CZ, Český Krumlov, 48°50′ N, 14°19′ E, July 2018 | 570 m | 16/16 | – | 10/10 |
| Chazara briseis | CZ, Raná, 50°24′ N, 13°46′ E, August 2014–18 | 450 m | 16/14 | – | 10/10 |
| Coeneonympha arcania | CZ, Hodonínská Dúbrava, 48°53′ N, 17°6′ E, June 2018 | 200 m | 16/16 | – | 10/10 |
| Coeneonympha gardetta | AT, Heiligenblutt, 47°3′ N, 12°47′ E, August 2018 | 1950 m | 15/10 | – | 10/10 |
| Hipparchia semele | CZ, Prokopské Údolí, 50°2′ N, 14°21′ E, August 2018 | 300 m | 15/10 | – | 10/10 |
| Maniola jurtina | CZ, Český Krumlov, 48°50′ N, 14°19′ E, July 2018 | 570 m | 16/16 | – | 10/10 |
| Melanargia galathea | CZ, Český Krumlov, 48°50′ N, 14°19′ E, July 2018 | 570 m | 16/16 | – | 10/10 |
| Minois dryas | CZ, Lázně Bohdaneč, 50°4′ N, 15°41′ E, August 2018 | 250 m | 15/8 | – | 10/8 |
| Ref. [15] (SCP, TSPC) | |||||
| Erebia medusa | CZ, Český Krumlov, 48°50′ N, 14°19′ E, May 2015 | 570 m | 16/16 | – | 10/10 |
| Erebia aethiops | AT, Tirol, Au, 47°06′ N, 10°57′ E, August 2015 | 1200 m | 16/16 | – | 10/10 |
| Erebia pronoe | AT, Pfafflar, 47°17′ N, 10°39′ E, August 2015 | 1200 m | 16/6 | – | 10/10 |
| Erebia cassioides | AT, Hochgurgl, 46°54′ N, 11°03′ E, August 2015 | 2200 m | 16/12 | – | 10/10 |
| Erebia pluto | AT, Rettenbachgletscher, 46°56′ N, 10°55′ E, August 2015 | 2900 m | 15/– | – | 10/– |
| Ref. [14] (SCP, LLt) | |||||
| Erebia medusa | CZ, Český Krumlov, 48°50′ N, 14°19′ E, May 2010 | 570 m | 16 | 10 | – |
| Erebia sudetica | CZ, Praděd Mt., 50°4′ N, 17°13′ E, August 2010 | 1320 m | 16 | 10 | – |
| Erebia epiphron | CZ, Praděd Mt.,50°4′ N, 17°13′ E, August 2010 | 1460 m | 16 | 10 | – |
| Erebia tyndarus | AT, Sölden, Windachtal, 46°57′ N, 11°3′ E, August 2010 | 1950 m | 16 | 10 | – |
| Eight Non-Erebia Satyrinae | Eig1 | Eig2 | Eig3 | Eig4 | Var. | F, P1st axis | F, Pall axes |
|---|---|---|---|---|---|---|---|
| Species | 0.292 | 0.154 | 0.100 | 0.022 | 58.2% | 62.0 *** | 32.2 *** |
| Treatment | 0.051 | 4.5% | 8.4 ** | ||||
| Species|treatment | 0.293 | 0.153 | 0.100 | 0.022 | 61.8% | 66.6 *** | 36.8 *** |
| Treatment|species | 0.052 | 12.5% | 22.4 *** | ||||
| Added 5 Erebia spp. [15] | |||||||
| Species | 0.326 | 0.124 | 0.065 | 0.047 | 63.1% | 9.5 *** | 33.5 *** |
| Treatment | 0.034 | 3.4% | 8.6 *** | ||||
| Species|treatment | 0.328 | 0.123 | 0.068 | 0.049 | 65.9% | 10.0 *** | 37.7 *** |
| Treatment|species | 0.039 | 10.7% | 27.9 *** |
| Autumn | Winter | |||||||
|---|---|---|---|---|---|---|---|---|
| Trait | K | p | λ | p | K | p | λ | p |
| Glycerol | 0.47 | 0.40 | 0.22 | 0.77 | 1.97 | <0.001 | 1.11 | <0.0001 |
| Fructose | 0.38 | 0.74 | <0.01 | 1.00 | 0.47 | 0.50 | <0.01 | 1.00 |
| Glucose | 0.45 | 0.51 | 0.31 | 0.74 | 0.32 | 0.79 | <0.01 | 1.00 |
| Sucrose | 0.51 | 0.36 | <0.01 | 1.00 | 0.23 | 0.96 | <0.01 | 1.00 |
| Trehalose | 0.36 | 0.77 | <0.01 | 1.00 | 0.42 | 0.61 | <0.01 | 1.00 |
| SCP | 0.69 | 0.11 | <0.01 | 1.00 | 0.94 | 0.03 | 0.79 | 0.15 |
| TSPC | 0.45 | 0.50 | <0.01 | 1.00 | 0.77 | 0.07 | 0.42 | 0.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrba, P.; Sucháčková Bartoňová, A.; Andres, M.; Nedvěd, O.; Šimek, P.; Konvička, M. Exploring Cold Hardiness within a Butterfly Clade: Supercooling Ability and Polyol Profiles in European Satyrinae. Insects 2022, 13, 369. https://doi.org/10.3390/insects13040369
Vrba P, Sucháčková Bartoňová A, Andres M, Nedvěd O, Šimek P, Konvička M. Exploring Cold Hardiness within a Butterfly Clade: Supercooling Ability and Polyol Profiles in European Satyrinae. Insects. 2022; 13(4):369. https://doi.org/10.3390/insects13040369
Chicago/Turabian StyleVrba, Pavel, Alena Sucháčková Bartoňová, Miloš Andres, Oldřich Nedvěd, Petr Šimek, and Martin Konvička. 2022. "Exploring Cold Hardiness within a Butterfly Clade: Supercooling Ability and Polyol Profiles in European Satyrinae" Insects 13, no. 4: 369. https://doi.org/10.3390/insects13040369
APA StyleVrba, P., Sucháčková Bartoňová, A., Andres, M., Nedvěd, O., Šimek, P., & Konvička, M. (2022). Exploring Cold Hardiness within a Butterfly Clade: Supercooling Ability and Polyol Profiles in European Satyrinae. Insects, 13(4), 369. https://doi.org/10.3390/insects13040369