
The Use of Tuning Forks for Studying Behavioural Responses in Orb Web Spiders



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Study Species
2.3. Data Collection
2.4. Data Analysis
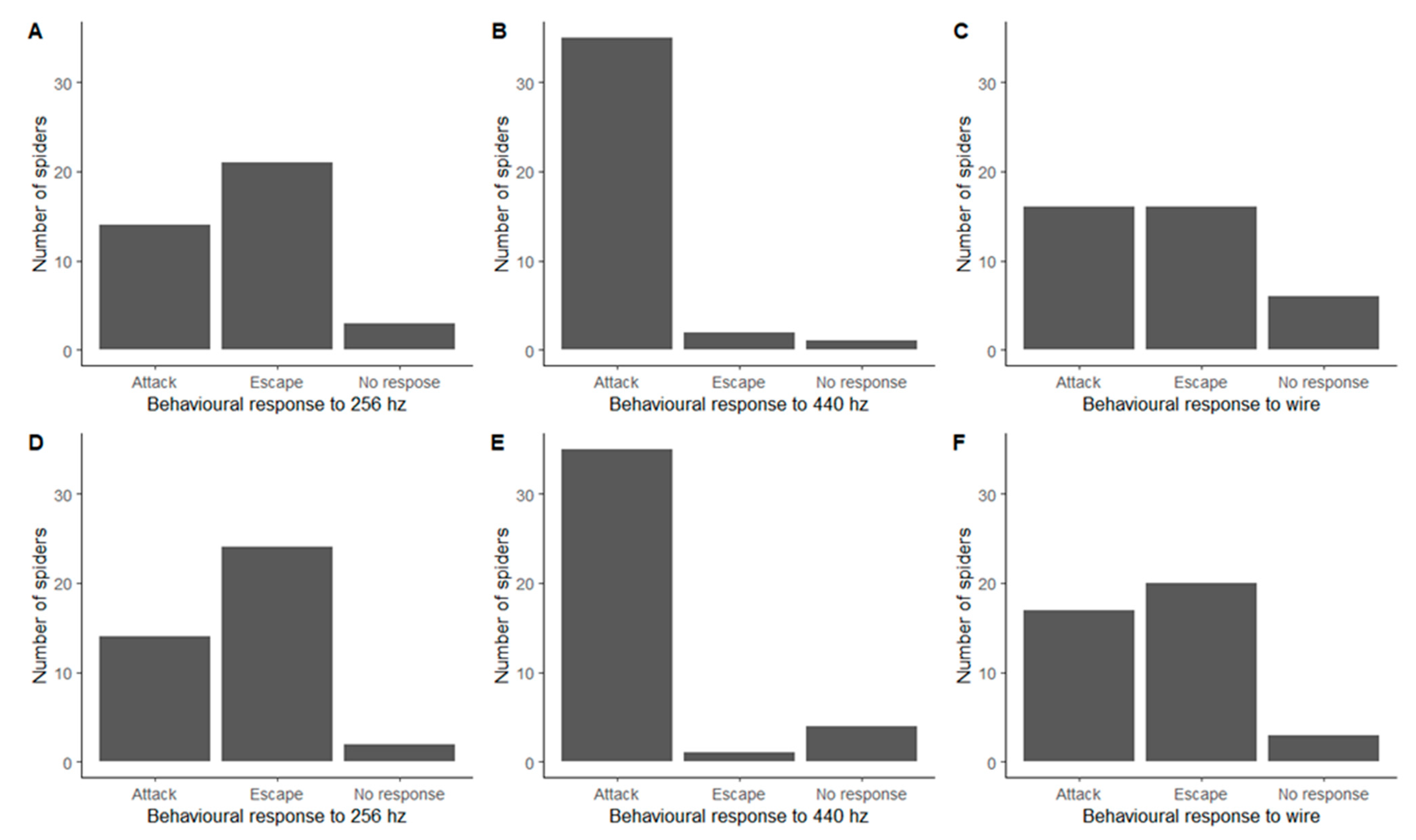
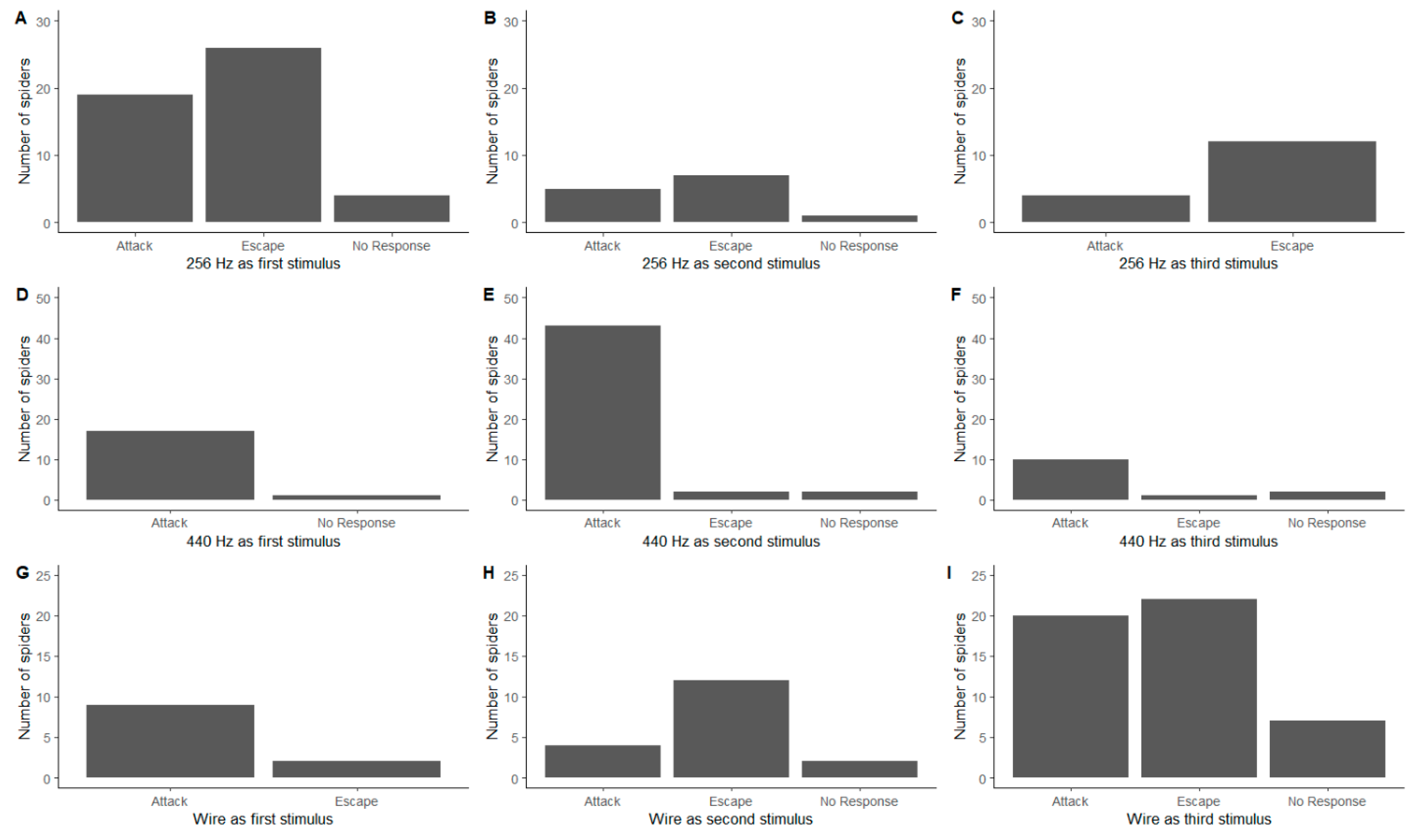
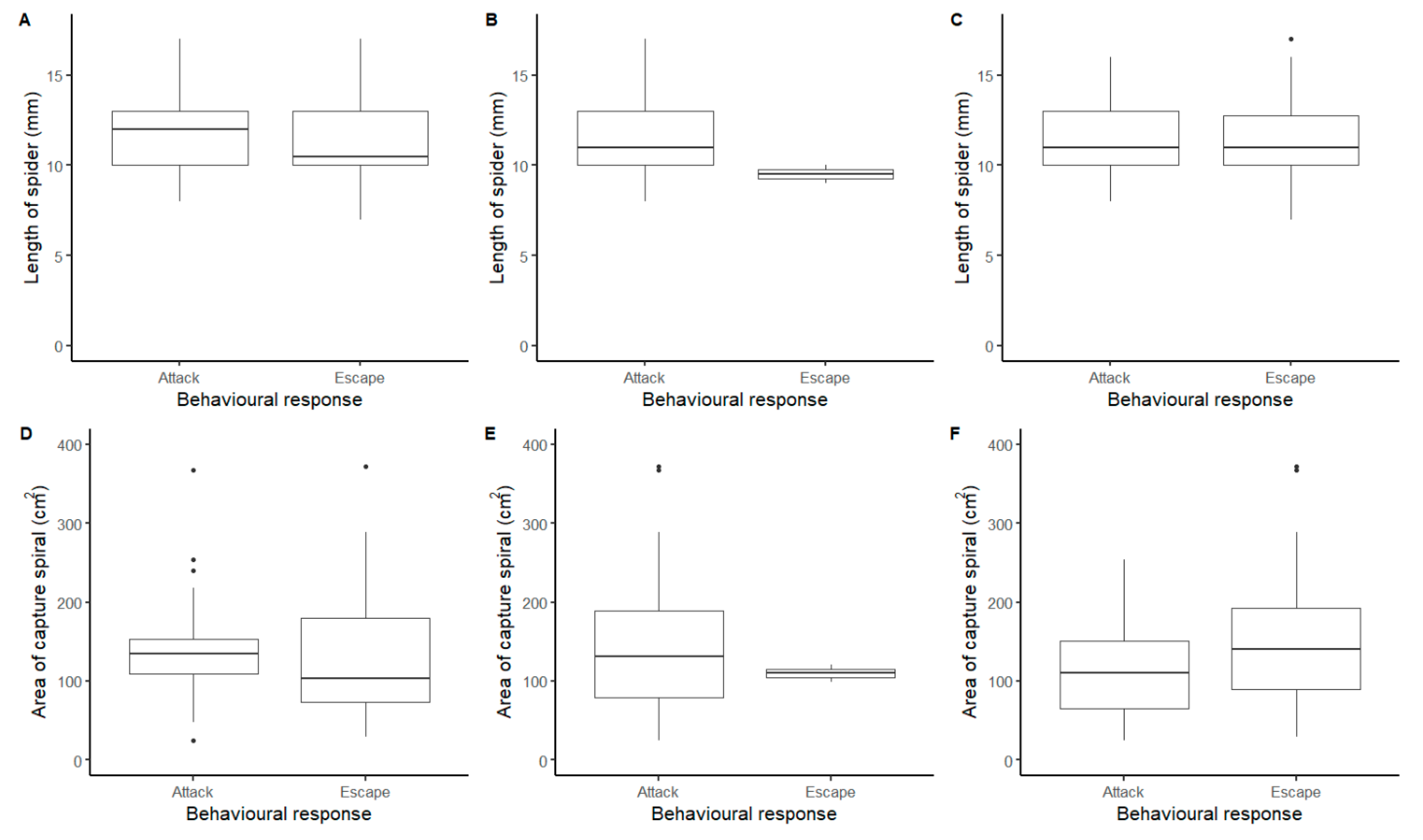
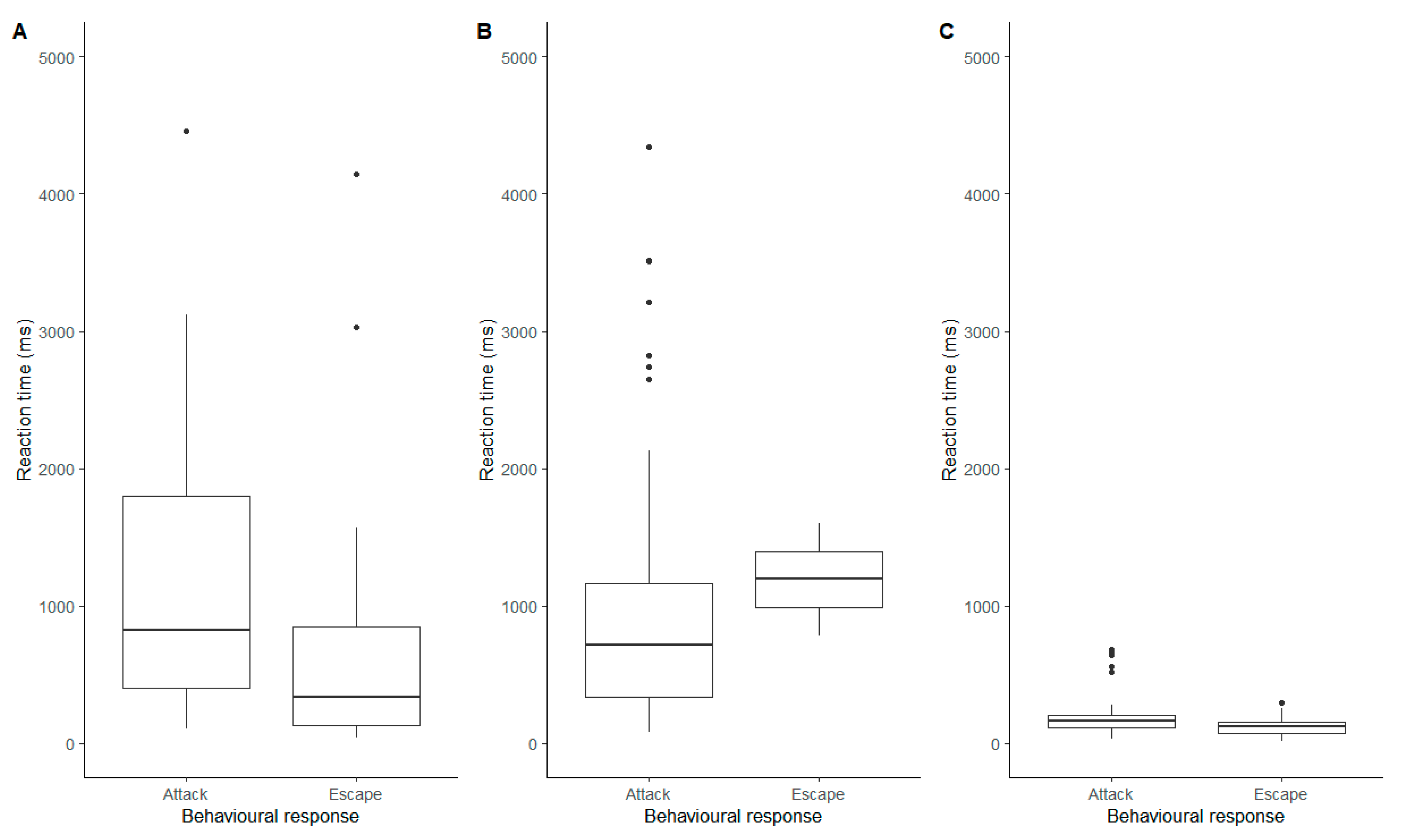
3. Results
3.1. Behavioural Responses
3.2. Order of Stimuli
3.3. Effect of Size and Web Area
3.4. Reaction Times
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herberstein, M.E.; Wignall, A.E.; Hebets, E.A.; Schneider, J.M. Dangerous mating systems: Signal complexity, signal content and neural capacity in spiders. Neurosci. Biobehav. Rev. 2014, 46, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Japyassu, H.F.; Laland, K.N. Extended spider cognition. Anim. Cogn. 2017, 20, 375–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarger, J.L.; Cherry, B.R.; van der Vaart, A. Uncovering the structure-function relationship in spider silk. Nat. Rev. Mater. 2018, 3, 18008. [Google Scholar] [CrossRef]
- Blamires, S.J. Plasticity in extended phenotypes: Orb web architectural responses to variations in prey parameters. J. Exp. Biol. 2010, 213, 3207–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punzo, F. Spiders: Biology, Ecology, Natural History and Behaviour; Brill: Leiden, The Netherlands, 2007. [Google Scholar]
- Blackledge, T.A.; Kunter, M.; Agnarsson, I. The form and function of spider orb webs: Evolution from silk to ecosystems. Adv. Insect Physiol. 2011, 41, 175–262. [Google Scholar] [CrossRef]
- Zschokke, S.; Herberstein, M.E. Laboratory methods for maintaining and studying web-building spiders. J. Arachnol. 2005, 33, 205–213. [Google Scholar] [CrossRef]
- Hesselberg, T. Exploration behaviour and behavioural flexibility in orb-web spiders: A review. Curr. Zool. 2015, 61, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Scharf, I.; Lubin, Y.; Ovadia, O. Foraging decisions and behavioural flexibility in trap-building predators: A review. Biol. Rev. 2011, 86, 626–639. [Google Scholar] [CrossRef]
- Cloudsley-Thompson, J.L. A review of anti-predator devices of spiders. Bull. Br. Arachnol. Soc. 1995, 10, 81–96. [Google Scholar]
- Tolbert, W.W. Predator avoidance behaviours and web defensive structures in the orb weavers Argiope aurantia and Argiope trifasciata (Araneae, Araneidae). Psyche 1975, 82, 29–52. [Google Scholar] [CrossRef] [Green Version]
- Uetz, G.W. Foraging strategies of spiders. Trends Ecol. Evol. 1992, 7, 155–159. [Google Scholar] [CrossRef]
- Turner, J.; Vollrath, F.; Hesselberg, T. Wind speed affects prey-catching behaviour in an orb web spider. Naturwissenschaften 2011, 98, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S.; Hénaut, Y.; Benjamin, S.P.; García-Ballinas, J.A. Prey-capture strategies in sympatric web-building spiders. Can. J. Zool. 2006, 84, 964–973. [Google Scholar] [CrossRef]
- Mulder, T.; Mortimer, B.; Vollrath, F. Functional flexibility in a spider’s orb web. J. Exp. Biol. 2020, 223, jeb234070. [Google Scholar] [CrossRef]
- Tew, E.R.; Adamson, A.; Hesselberg, T. The web repair behaviour of an orb spider. Anim. Behav. 2015, 103, 137–146. [Google Scholar] [CrossRef]
- Watts, J.C.; Herrig, A.; Allen, W.D.; Jones, T.C. Diel patterns of foraging aggression and antipredator behaviour in the trashline orb-weaving spider, Cyclosa turbinata. Anim. Behav. 2014, 94, 79–86. [Google Scholar] [CrossRef]
- Blamires, S.J.; Lee, Y.-H.; Chang, C.-M.; Lin, I.-T.; Chen, J.-A.; Lin, T.-Y.; Tso, I.-M. Multiple structures interactively in-fluence prey capture efficiency in spider orb webs. Anim. Behav. 2010, 80, 947–953. [Google Scholar] [CrossRef]
- Tseng, H.-J.; Cheng, R.-C.; Wu, S.-H.; Blamires, S.J.; Tso, I.-M. Trap barricading and decorating by a well-armored sit-and-wait predator: Extra protection or prey attraction? Behav. Ecol. Sociobiol. 2011, 65, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Corey, T.B.; Hebets, E.A. Testing the hypothesized antipredator defence function of stridulation in the spiny orb-weaving spider, Micrathena gracilis. Anim. Behav. 2020, 169, 103–117. [Google Scholar] [CrossRef]
- Foelix, R. Biology of Spiders; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Mortimer, B.; Soler, A.; Wilkins, L.; Vollrath, F. Decoding the locational information in the orb web vibrations of Araneus diadematus and Zygiella x-notata. J. R. Soc. Interface 2019, 16, 20190201. [Google Scholar] [CrossRef] [Green Version]
- Landolfa, M.A.; Barth, F.G. Vibrations in the orb web of the spider Nephila calvipes: Cues for discrimination and orientation. J. Comp. Physiol. A 1996, 179, 493–508. [Google Scholar] [CrossRef]
- Boys, C.V. The Influence of a tuning fork on the garden spider. Nature 1880, 23, 149–150. [Google Scholar] [CrossRef]
- Barrows, W.B. The reactions of an orb-weaving spider, Eperia sclopetaria Clerck, to rhythmic vibrations of its web. Biol. Bull. 1915, 29, 316–332. [Google Scholar] [CrossRef] [Green Version]
- Ganske, A.-S.; Uhl, G. The sensory equipment of a spider–A morphological survey of different types of sensillum in both sexes of Argiope bruennichi (Araneae, Araneidae). Arthropod Struct. Dev. 2018, 47, 144–161. [Google Scholar] [CrossRef]
- Bays, S.M. A study of the training possibilities of Araneus diadematus Cl. Experientia 1962, 18, 423–424. [Google Scholar] [CrossRef]
- Justice, M.J.; Justice, T.C.; Vesci, R.L. Web orientation, stabilimentum structure and predatory behavior of Argiope florida Chamberlain & Ivie 1944 (Araneae, Araneidae, Argiopinae). J. Arachnol. 2005, 33, 82–92. [Google Scholar]
- Nakata, K.; Mori, Y. Cost of complex behaviours and its implications in antipredator defence in orb-web spiders. Anim. Behav. 2016, 120, 115–121. [Google Scholar] [CrossRef]
- British Spider Recording Scheme. Summary for Metellina segmentata sens. str. (Araneae). 2021. Available online: http://srs.britishspiders.org.uk/portal.php/p/Summary/s/Metellina+segmentata+sens.+str (accessed on 21 November 2021).
- Tew, N.; Hesselberg, T. Web asymmetry in the tetragnathid orb spider Metellina mengei (Blackwell, 1869) is determined by web inclination and web size. J. Arachnol. 2018, 46, 370–372. [Google Scholar] [CrossRef]
- de Nadai, B.L.; Maletzke, A.G.; Corbi, J.J.; Batista, G.E.A.P.A.; Reiskind, M.H. The impact of body size of Aedes [Stegomyia] aegypti wingbeat frequency: Implications for mosquito identification. Med. Vet. Entomol. 2021, 35, 617–624. [Google Scholar] [CrossRef]
- Kawakita, S.; Ichikawa, K. Automated classification of bees and hornet using acoustic analysis of their flight sounds. Apidologie 2019, 50, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Herberstein, M.E.; Tso, I.M. Evaluation of formulae to estimate the capture area of orb webs (Araneoidea, Araneae). J. Arachnol. 2000, 28, 180–184. [Google Scholar] [CrossRef]
- Mehta, C.R.; Patel, N.R. Algorithm 643: FEXACT, a FORTRAN subroutine for Fisher’s exact test on unordered r x c contingency tables. ACM Trans. Math. Softw. 1986, 12, 154–161. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 3 March 2022).
- Sewlal, J.N.; Dempewolf, L. Defence mechanisms of the orb-weaving spider Azilia vachoni (Araneae:Tetragnathidae). Living World J. Trinidad Tobago Field Nat. Club 2005, 2005, 50–52. [Google Scholar]
- Willey, M.B.; Johnson, M.A.; Adler, P.H. Predatory behaviour of the basilica spider Mecynogea lemniscata (Araneae, Araneidae). Psyche 1992, 99, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Lubin, Y.D. The predatory behaviour of Crytophora (Araneae: Araneidae). J. Arachnol. 1980, 8, 159–185. [Google Scholar]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioural syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Kralj-Fišer, S.; Schneider, J.M. Individual behavioural consistency and plasticity in an urban spider. Anim. Behav. 2012, 84, 197–204. [Google Scholar] [CrossRef]
- Nakata, K. Spiders use airborne cues to respond to flying insect predators by building orb-web with fewer silk thread and larger silk decorations. Ethology 2008, 114, 686–692. [Google Scholar] [CrossRef]
- Barth, F.G.; Höller, A. Dynamics of arthropod filiform hairs. V. The response of spider trichobothria to natural stimuli. Philos. Trans. Biol. Sci. 1999, 354, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Savory, T. Arachnida, 2nd ed.; Academic Press: London, UK, 1977. [Google Scholar]
- Hesselberg, T.; Simonsen, D.; Juan, C. Do cave orb spiders show unique behavioural adaptations to subterranean life? A review of the evidence. Behaviour 2019, 156, 969–996. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Lunghi, E.; Bilandžija, H.; Cardoso, P.; Grimm, V.; Schmidt, S.I.; Hesselberg, T.; Martínez, A. Collecting eco-evolutionary data in the data: Impediments to subterranean research and how to overcome them. Ecol. Evol. 2021, 11, 5911–5926. [Google Scholar] [CrossRef] [PubMed]
- Mammola, S. Finding answers in the dark: Caves as models in ecology fifty years after Poulson and White. Ecography 2018, 42, 1331–1351. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, D.; Hesselberg, T. Unique behavioural modifications in the web structure of the cave orb spider Meta menardi (Araneae, Tetragnathidae). Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Hesselberg, T.; Simonsen, D. A comparison of morphology and web geometry between hypogean and epigean species of Metellina orb spiders (family Tetragnathidae). Subterr. Biol. 2019, 32, 1–13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioural Response | Stimulus | No. of Responses | |
| Attack | |||
| Turning towards the tuning fork | The spider, whilst staying on the central hub, turned to face the direction of the tuning fork, but did not move from the hub. | 256 Hz and 440 Hz | 2 (2020) 6 (2021) |
| Moving towards the tuning fork | The spider moved towards the location of the tuning fork, then stopped on the capture spiral, or returned to the central hub. | 256 Hz and 440 Hz | 23 (2020) 8 (2021) |
| Touching the tuning fork | The spider moved towards the tuning fork, and then touched it, but remained on the web. | 440 Hz | 1 (2020) 15 (2021) |
| Moving towards the tuning fork, then dropping from the web | The spider moved towards the tuning fork, but before touching it/getting close, the spider dropped from the web. | 256 Hz and 440 Hz | 17 (2020) 15 (2021) |
| Touching the tuning fork, then dropping from the web | The spider moved towards the tuning fork, after coming into contact with it, the spider dropped from the web. | 440 Hz | 6 (2020) 2 (2021) |
| Attacking | The spider moved its legs, to either grab or fight the wire, and remained on the web. | Wire | 11 (2020) 12 (2021) |
| Grabbing | The spider held onto the wire, coming off the web. | Wire | 5 (2020) 5 (2021) |
| Escape | |||
| Dropping from the web | The spider dropped off the web; either to the floor, vegetation below, or in the air. | 256 Hz, 440 Hz and wire | 22 (2020) 27 (2021) |
| Moving/running away | The spider ran away, usually in the opposite direction to the tuning fork, moving off the web onto adjacent vegetation | 256 Hz and wire | 8 (2020) 8 (2021) |
| Jumping away | The spider jumped away after being touched by the wire, but remained on the web. | Wire | 9 (2020) 10 (2021) |
| No response | |||
| Flinching | The spider’s body moved slightly. It recoiled, but remained on the web. | Wire | 6 (2020) 2 (2021) |
| No response | The spider did not respond to the stimuli in any way. | 256 Hz, 440 Hz, wire | 4 (2020) 7 (2021) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davies, M.S.; Hesselberg, T. The Use of Tuning Forks for Studying Behavioural Responses in Orb Web Spiders. Insects 2022, 13, 370. https://doi.org/10.3390/insects13040370
Davies MS, Hesselberg T. The Use of Tuning Forks for Studying Behavioural Responses in Orb Web Spiders. Insects. 2022; 13(4):370. https://doi.org/10.3390/insects13040370
Chicago/Turabian StyleDavies, Mollie S., and Thomas Hesselberg. 2022. "The Use of Tuning Forks for Studying Behavioural Responses in Orb Web Spiders" Insects 13, no. 4: 370. https://doi.org/10.3390/insects13040370
APA StyleDavies, M. S., & Hesselberg, T. (2022). The Use of Tuning Forks for Studying Behavioural Responses in Orb Web Spiders. Insects, 13(4), 370. https://doi.org/10.3390/insects13040370