
Effects of Thiamethoxam-Dressed Oilseed Rape Seeds and Nosema ceranae on Colonies of Apis mellifera iberiensis, L. under Field Conditions of Central Spain. Is Hormesis Playing a Role?

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Soil Characterization and Climatological Data
2.3. Sampling Schedule
2.4. Palynological and Melissopalynological Assessment
2.5. Chemical Analysis
2.6. Identification of Pathogens
2.6.1. Detection of Varroa Mites
2.6.2. Pathogen Screening
2.7. Statistical Analysis
3. Results
3.1. Soil Characterization and Climatic Conditions
3.2. Palynological and Melissopalynological Assessments
3.3. Chemical Analysis
3.4. Field Monitoring of the Honey Bee Colonies
3.5. Identification of Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brodschneider, R.; Gray, A.; Adjlane, N.; Ballis, A.; Brusbardis, V.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Dahle, B.; de Graaf, D.C.; et al. Multi-country loss rates of honey bee colonies during winter 2016/2017 from the COLOSS survey. J. Apic. Res. 2018, 57, 452–457. [Google Scholar] [CrossRef] [Green Version]
- Brodschneider, R.; Gray, A.; van der Zee, R.; Adjlane, N.; Brusbardis, V.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Crailsheim, K.; Dahle, B.; et al. Preliminary analysis of loss rates of honey bee colonies during winter 2015/16 from the COLOSS survey. J. Apic. Res. 2016, 55, 375–378. [Google Scholar] [CrossRef] [Green Version]
- Gray, A.; Adjlane, N.; Arab, A.; Ballis, A.; Brusbardis, V.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Cornelissen, B.; da Costa, C.A.; et al. Honey bee colony winter loss rates for 35 countries participating in the COLOSS survey for winter 2018–2019, and the effects of a new queen on the risk of colony winter loss. J. Apic. Res. 2020, 59, 744–751. [Google Scholar] [CrossRef]
- Gray, A.; Brodschneider, R.; Adjlane, N.; Ballis, A.; Brusbardis, V.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Cornelissen, B.; Amaro da Costa, C.; et al. Loss rates of honey bee colonies during winter 2017/18 in 36 countries participating in the COLOSS survey, including effects of forage sources. J. Apic. Res. 2019, 58, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.V.; Steinhauer, N.; Rennich, K.; Wilson, M.E.; Tarpy, D.R.; Caron, D.M.; Rose, R.; Delaplane, K.S.; Baylis, K.; Lengerich, E.J.; et al. A national survey of managed honey bee 2013–2014 annual colony losses in the USA. Apidologie 2015, 46, 292–305. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.; Carreck, N.L. Honey bee colony losses. J. Apic. Res. 2010, 49, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Van der Zee, R.; Gray, A.; Pisa, L.; de Rijk, T. An Observational Study of Honey Bee Colony Winter Losses and Their Association with Varroa destructor, Neonicotinoids and Other Risk Factors. PLoS ONE 2015, 10, e0131611. [Google Scholar] [CrossRef] [Green Version]
- Van der Zee, R.; Pisa, L.; Andonov, S.; Brodschneider, R.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Crailsheim, K.; Dahle, B.; Gajda, A.; et al. Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008–2009 and 2009–2010. J. Apic. Res. 2012, 51, 100–114. [Google Scholar] [CrossRef]
- VanEngelsdorp, D.; Caron, D.; Hayes, J.; Underwood, R.; Henson, M.; Rennich, K.; Spleen, A.; Andree, M.; Snyder, R.; Lee, K.; et al. A national survey of managed honey bee 2010–11 winter colony losses in the USA: Results from the Bee Informed Partnership. J. Apic. Res. 2012, 51, 115–124. [Google Scholar] [CrossRef] [Green Version]
- VanEngelsdorp, D.; Hayes, J., Jr.; Underwood, R.M.; Pettis, J. A Survey of Honey Bee Colony Losses in the U.S., Fall 2007 to Spring 2008. PLoS ONE 2009, 3, e4071. [Google Scholar] [CrossRef]
- VanEngelsdorp, D.; Hayes, J.; Underwood, R.M.; Pettis, J.S. A survey of honey bee colony losses in the United States, Fall 2008 to Spring 2009. J. Apic. Res. 2010, 49, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Carreck, N.L. A beekeeper’s perspective on the neonicotinoid ban. Pest Manag. Sci. 2017, 73, 1295–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, A.; Eickermann, M.; Kraus, F.; Georges, C.; Hoffmann, L.; Beyer, M. A survey on some factors potentially affecting losses of managed honey bee colonies in Luxembourg over the winters 2010/2011 and 2011/2012. J. Apic. Res. 2014, 53, 43–56. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors Associated with Honey Bee Colony Losses: A Mini-Review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Johnson, R.M. Honey Bee Toxicology. Annu. Rev. Entomol. 2015, 60, 415–434. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Switanek, M.; Crailsheim, K.; Truhetz, H.; Brodschneider, R. Modelling seasonal effects of temperature and precipitation on honey bee winter mortality in a temperate climate. Sci. Total Environ. 2017, 579, 1581–1587. [Google Scholar] [CrossRef]
- VanEngelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef]
- Neov, B.; Georgieva, A.; Shumkova, R.; Radoslavov, G.; Hristov, P. Biotic and Abiotic Factors Associated with Colonies Mortalities of Managed Honey Bee (Apis mellifera). Diversity 2019, 11, 237. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, I.a.C.; John, E. Nicotinoid Insecticides and the Nicotinic Acetylcholine Receptor; Springer: Tokyo, Japan, 1999. [Google Scholar]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest Manag. Sci. 2008, 64, 1099–1105. [Google Scholar] [CrossRef]
- Jeschke, P. Status and outlook for acaricide and insecticide discovery. Pest Manag. Sci. 2021, 77, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R. Neonicotinoids—From zero to hero in insecticide chemistry. Pest Manag. Sci. 2008, 64, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R.; Beck, M.E. Nicotinic Acetylcholine Receptor Agonists: A Milestone for Modern Crop Protection. Angew. Chem. Int. Ed. 2013, 52, 9464–9485. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M.; Sattelle, D.B. Neonicotinoids: Insecticides acting on insect nicotinic acetylcholine receptors. Trends Pharmacol. Sci. 2001, 22, 573–580. [Google Scholar] [CrossRef]
- Matsuda, K.; Kanaoka, S.; Akamatsu, M.; Sattelle, D.B. Diverse Actions and Target-Site Selectivity of Neonicotinoids: Structural Insights. Mol. Pharmacol. 2009, 76, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tomizawa, M.; Casida, J.E. Neonicotinoid Insecticide Toxicology: Mechanisms of Selective Action. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 247–268. [Google Scholar] [CrossRef] [Green Version]
- Tomizawa, M.; Lee, D.L.; Casida, J.E. Neonicotinoid Insecticides: Molecular Features Conferring Selectivity for Insect versus Mammalian Nicotinic Receptors. J. Agric. Food Chem. 2000, 48, 6016–6024. [Google Scholar] [CrossRef]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.D.; et al. Environmental fate and exposure; Neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef]
- Borsuah, J.F.; Messer, T.L.; Snow, D.D.; Comfort, S.D.; Mittelstet, A.R. Literature Review: Global Neonicotinoid Insecticide Occurrence in Aquatic Environments. Water 2020, 12, 3388. [Google Scholar] [CrossRef]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 24, 17285–17325. [Google Scholar] [CrossRef] [PubMed]
- Van der Sluijs, J.P.; Simon-Delso, N.; Goulson, D.; Maxim, L.; Bonmatin, J.-M.; Belzunces, L.P. Neonicotinoids, bee disorders and the sustainability of pollinator services. Curr. Opin. Environ. Sustain. 2013, 5, 293–305. [Google Scholar] [CrossRef]
- Blacquière, T.; Smagghe, G.; van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decourtye, A.; Devillers, J. Ecotoxicity of neonicotinoid insecticides to bees. Adv. Exp. Med. Biol. 2010, 683, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulson, D. Review: An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- Hatjina, F.; Papaefthimiou, C.; Charistos, L.; Dogaroglu, T.; Bouga, M.; Emmanouil, C.; Arnold, G. Sublethal doses of imidacloprid decreased size of hypopharyngeal glands and respiratory rhythm of honeybees in vivo. Apidologie 2013, 44, 467–480. [Google Scholar] [CrossRef] [Green Version]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A Common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef]
- Lundin, O.; Rundlöf, M.; Smith, H.G.; Fries, I.; Bommarco, R. Neonicotinoid Insecticides and Their Impacts on Bees: A Systematic Review of Research Approaches and Identification of Knowledge Gaps. PLoS ONE 2015, 10, e0136928. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.W.; Tautz, J.; Grünewald, B.; Fuchs, S. RFID Tracking of Sublethal Effects of Two Neonicotinoid Insecticides on the Foraging Behavior of Apis mellifera. PLoS ONE 2012, 7, e30023. [Google Scholar] [CrossRef] [Green Version]
- Tackenberg, M.C.; Giannoni-Guzmán, M.A.; Sanchez-Perez, E.; Doll, C.A.; Agosto-Rivera, J.L.; Broadie, K.; Moore, D.; McMahon, D.G. Neonicotinoids disrupt circadian rhythms and sleep in honey bees. Sci. Rep. 2020, 10, 17929. [Google Scholar] [CrossRef] [PubMed]
- Tavares, D.A.; Dussaubat, C.; Kretzschmar, A.; Carvalho, S.M.; Silva-Zacarin, E.C.M.; Malaspina, O.; Bérail, G.; Brunet, J.-L.; Belzunces, L.P. Exposure of larvae to thiamethoxam affects the survival and physiology of the honey bee at post-embryonic stages. Environ. Pollut. 2017, 229, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Tavares, D.A.; Roat, T.C.; Carvalho, S.M.; Silva-Zacarin, E.C.M.; Malaspina, O. In vitro effects of thiamethoxam on larvae of Africanized honey bee Apis mellifera (Hymenoptera: Apidae). Chemosphere 2015, 135, 370–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosi, S.; Nieh, J.C. A common neonicotinoid pesticide, thiamethoxam, alters honey bee activity, motor functions, and movement to light. Sci. Rep. 2017, 7, 15132. [Google Scholar] [CrossRef]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid Pesticide Reduces Bumble Bee Colony Growth and Queen Production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [Green Version]
- Annoscia, D.; Di Prisco, G.; Becchimanzi, A.; Caprio, E.; Frizzera, D.; Linguadoca, A.; Nazzi, F.; Pennacchio, F. Neonicotinoid Clothianidin reduces honey bee immune response and contributes to Varroa mite proliferation. Nat. Commun. 2020, 11, 5887. [Google Scholar] [CrossRef]
- Catae, A.F.; Roat, T.C.; De Oliveira, R.A.; Nocelli, R.C.; Malaspina, O. Cytotoxic effects of thiamethoxam in the midgut and malpighian tubules of Africanized Apis mellifera (Hymenoptera: Apidae). Microsc. Res. Tech. 2014, 77, 274–281. [Google Scholar] [CrossRef]
- Gregorc, A.; Silva-Zacarin, E.C.M.; Carvalho, S.M.; Kramberger, D.; Teixeira, E.W.; Malaspina, O. Effects of Nosema ceranae and thiametoxam in Apis mellifera: A comparative study in Africanized and Carniolan honey bees. Chemosphere 2016, 147, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.A.; Roat, T.C.; Carvalho, S.M.; Malaspina, O. Side-effects of thiamethoxam on the brain andmidgut of the africanized honeybee Apis mellifera (Hymenopptera: Apidae). Environ. Toxicol. 2014, 29, 1122–1133. [Google Scholar] [CrossRef]
- Shi, T.-F.; Wang, Y.-F.; Liu, F.; Qi, L.; Yu, L.-S. Sublethal Effects of the Neonicotinoid Insecticide Thiamethoxam on the Transcriptome of the Honey Bees (Hymenoptera: Apidae). J. Econ. Entomol. 2017, 110, 2283–2289. [Google Scholar] [CrossRef]
- Paris, L.; Peghaire, E.; Moné, A.; Diogon, M.; Debroas, D.; Delbac, F.; El Alaoui, H. Honeybee gut microbiota dysbiosis in pesticide/parasite co-exposures is mainly induced by Nosema ceranae. J. Invertebr. Pathol. 2020, 172, 107348. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; Martín-Hernández, R.; Meana, A. Nosema ceranae in Europe: An emergent type C nosemosis. Apidologie 2010, 41, 375–392. [Google Scholar] [CrossRef] [Green Version]
- Martín-Hernández, R.; Bartolomé, C.; Chejanovsky, N.; Le Conte, Y.; Dalmon, A.; Dussaubat, C.; García-Palencia, P.; Meana, A.; Pinto, M.A.; Soroker, V.; et al. Nosema ceranae in Apis mellifera: A 12 years postdetection perspective. Environ. Microbiol. 2018, 20, 1302–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Palencia, P.; Martín-Hernández, R.; González-Porto, A.-V.; Marin, P.; Meana, A.; Higes, M. Natural infection by Nosema ceranae causes similar lesions as in experimentally infected caged-worker honey bees (Apis mellifera). J. Apic. Res. 2010, 49, 278–283. [Google Scholar] [CrossRef]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef]
- Goblirsch, M. Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie 2018, 49, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Paris, L.; El Alaoui, H.; Delbac, F.; Diogon, M. Effects of the gut parasite Nosema ceranae on honey bee physiology and behavior. Curr. Opin. Insect Sci. 2018, 26, 149–154. [Google Scholar] [CrossRef]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G.; et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef]
- Dussaubat, C.; Maisonnasse, A.; Alaux, C.; Tchamitchan, S.; Brunet, J.-L.; Plettner, E.; Belzunces, L.P.; Le Conte, Y. Nosema spp. Infection Alters Pheromone Production in Honey Bees (Apis mellifera). J. Chem. Ecol. 2010, 36, 522–525. [Google Scholar] [CrossRef]
- Goblirsch, M.; Huang, Z.Y.; Spivak, M. Physiological and Behavioral Changes in Honey Bees (Apis mellifera) Induced by Nosema ceranae Infection. PLoS ONE 2013, 8, e58165. [Google Scholar] [CrossRef]
- Ares, A.M.; Nozal, M.J.; Bernal, J.L.; Martín-Hernández, R.; Higes, M.; Bernal, J. Liquid chromatography coupled to ion trap-tandem mass spectrometry to evaluate juvenile hormone III levels in bee hemolymph from Nosema spp. infected colonies. J. Chromatogr. B 2012, 899, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Vilarem, C.; Piou, V.; Vogelweith, F.; Vétillard, A. Varroa destructor from the Laboratory to the Field: Control, Biocontrol and IPM Perspectives—A Review. Insects 2021, 12, 800. [Google Scholar] [CrossRef] [PubMed]
- Traynor, K.S.; Mondet, F.; de Miranda, J.R.; Techer, M.; Kowallik, V.; Oddie, M.A.Y.; Chantawannakul, P.; McAfee, A. Varroa destructor: A Complex Parasite, Crippling Honey Bees Worldwide. Trends Parasitol. 2020, 36, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, K.P.; Madel, G. The influence of the mite Varroa jacobsoni Oud. on the protein concentration and the haemolymph volume of the brood of worker bees and drones of the honey bee Apis mellifera L. Apidologie 1985, 16, 421–436. [Google Scholar] [CrossRef] [Green Version]
- Sammataro, D.; Gerson, U.; Needham, G. Parasitic Mites of Honey Bees: Life History, Implications, and Impact. Annu. Rev. Entomol. 2000, 45, 519–548. [Google Scholar] [CrossRef]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [Green Version]
- Kralj, J.; Brockmann, A.; Fuchs, S.; Tautz, J. The parasitic mite Varroa destructor affects non-associative learning in honey bee foragers, Apis mellifera L. J. Comp. Physiol. A 2007, 193, 363–370. [Google Scholar] [CrossRef]
- Kralj, J.; Fuchs, S. Parasitic Varroa destructor mites influence flight duration and homing ability of infested Apis mellifera foragers. Apidologie 2006, 37, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Coulon, M.; Dalmon, A.; Di Prisco, G.; Prado, A.; Arban, F.; Dubois, E.; Ribière-Chabert, M.; Alaux, C.; Thiéry, R.; Le Conte, Y. Interactions Between Thiamethoxam and Deformed Wing Virus Can Drastically Impair Flight Behavior of Honey Bees. Front. Microbiol. 2020, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Brunet, J.-L.; Dussaubat, C.; Mondet, F.; Tchamitchan, S.; Cousin, M.; Brillard, J.; Baldy, A.; Belzunces, L.P.; Le Conte, Y. Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera). Environ. Microbiol. 2010, 12, 774–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettis, J.S.; vanEngelsdorp, D.; Johnson, J.; Dively, G. Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema. Naturwissenschaften 2012, 99, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doublet, V.; Labarussias, M.; de Miranda, J.R.; Moritz, R.F.A.; Paxton, R.J. Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 2015, 17, 969–983. [Google Scholar] [CrossRef]
- Vidau, C.; Diogon, M.; Aufauvre, J.; Fontbonne, R.; Viguès, B.; Brunet, J.-L.; Texier, C.; Biron, D.G.; Blot, N.; El Alaoui, H.; et al. Exposure to Sublethal Doses of Fipronil and Thiacloprid Highly Increases Mortality of Honeybees Previously Infected by Nosema ceranae. PLoS ONE 2011, 6, e21550. [Google Scholar] [CrossRef] [Green Version]
- Tesovnik, T.; Zorc, M.; Ristanić, M.; Glavinić, U.; Stevanović, J.; Narat, M.; Stanimirović, Z. Exposure of honey bee larvae to thiamethoxam and its interaction with Nosema ceranae infection in adult honey bees. Environ. Pollut. 2020, 256, 113443. [Google Scholar] [CrossRef]
- Dussaubat, C.; Brunet, J.-L.; Higes, M.; Colbourne, J.K.; Lopez, J.; Choi, J.-H.; Martín-Hernández, R.; Botías, C.; Cousin, M.; McDonnell, C.; et al. Gut pathology and responses to the microsporidium Nosema ceranae in the honey bee Apis mellifera. PLoS ONE 2012, 7, e37017. [Google Scholar] [CrossRef] [Green Version]
- Morfin, N.; Goodwin, P.H.; Guzman-Novoa, E. The combined effects of Varroa destructor parasitism and exposure to neonicotinoids affects honey bee (Apis mellifera L.) Memory and Gene Expression. Biology 2020, 9, 237. [Google Scholar] [CrossRef]
- Odemer, R. Effects of Chronic Pesticide and Pathogen Exposure on Honey Bee (Apis mellifera L.) Health at the Colony Level; Kommunikations, Informations und Medienzentrum der Universität Hohenheim: Stuttgart, Germany, 2018. [Google Scholar]
- Van Dooremalen, C.; Cornelissen, B.; Poleij-Hok-Ahin, C.; Blacquière, T. Single and interactive effects of Varroa destructor, Nosema spp., and imidacloprid on honey bee colonies (Apis mellifera). Ecosphere 2018, 9, e02378. [Google Scholar] [CrossRef] [Green Version]
- European and Mediterranean Plant Protection Organization. PP 1/170 (4): Side-effects on honeybees. EPPO Bull. 2010, 40, 313–319. [Google Scholar] [CrossRef]
- Schulz, D.J.; Huang, Z.-Y.; Robinson, G.E. Effects of colony food shortage on behavioral development in honey bees. Behav. Ecol. Sociobiol. 1998, 42, 295–303. [Google Scholar] [CrossRef]
- Higes, M.; Nozal, M.J.; Alvaro, A.; Barrios, L.; Meana, A.; Martín-Hernández, R.; Bernal, J.L.; Bernal, J. The stability and effectiveness of fumagillin in controlling Nosema ceranae (Microsporidia) infection in honey bees (Apis mellifera) under laboratory and field conditions. Apidologie 2011, 42, 364–377. [Google Scholar] [CrossRef] [Green Version]
- Ministerio de Agricultura, Pesca y Alimentación. Real Decreto 608/2006, de 19 de Mayo, por el que se Establece y Regula un Programa Nacional de Lucha y Control de las Enfermedades de las Abejas de la Miel; Ministerio de Agricultura: Madrid, Spain, 2006. [Google Scholar]
- International Soil Reference and Information Centre. Procedures for Soil Analysis, 6th ed.; van Reeuwijk, L., Ed.; ISRIC: Wageningen, The Netherlands, 2002. [Google Scholar]
- Bouyoucos, G.J. A Recalibration of the Hydrometer Method for Making Mechanical Analysis of Soils1. Agron. J. 1951, 43, 434–438. [Google Scholar] [CrossRef] [Green Version]
- Walter, H.; Lieth, H. Klimadiagramm—Weltatlas; VEB Gustav Fisher: Jena, Germany, 1967. [Google Scholar]
- Rivas-Martinez, S. Worldwide Bioclimatic Classification System, 1996–2021. Available online: http://www.globalbioclimatics.org/default.htm (accessed on 7 July 2021).
- Pérez, J.L.; Higes, M.; Suárez, M.; Llorente, J.; Meana, A. Easy ways to determine honey bee mortality using dead-bee traps. J. Apic. Res. 2001, 40, 25–28. [Google Scholar] [CrossRef]
- Botías, C.; Martín-Hernández, R.; Días, J.; García-Palencia, P.; Matabuena, M.; Juarranz, Á.; Barrios, L.; Meana, A.; Nanetti, A.; Higes, M. The effect of induced queen replacement on Nosema spp. infection in honey bee (Apis mellifera iberiensis) colonies. Environ. Microbiol. 2012, 14, 845–859. [Google Scholar] [CrossRef] [PubMed]
- Bernal, J.; Garrido-Bailón, E.; Del Nozal, M.J.; González-Porto, A.V.; Martín-Hernández, R.; Diego, J.C.; Jiménez, J.J.; Bernal, J.L.; Higes, M. Overview of pesticide residues in stored pollen and their potential effect on bee colony (Apis mellifera) losses in Spain. J. Econ. Entomol. 2010, 103, 1964–1971. [Google Scholar] [CrossRef] [PubMed]
- Cepero, A.; Ravoet, J.; Gómez-Moracho, T.; Bernal, J.L.; Del Nozal, M.J.; Bartolomé, C.; Maside, X.; Meana, A.; González-Porto, A.V.; de Graaf, D.C.; et al. Holistic screening of collapsing honey bee colonies in Spain: A case study. BMC Res. Notes 2014, 7, 649. [Google Scholar] [CrossRef] [Green Version]
- Higes, M.; Martín-Hernández, R.; Garrido-Bailón, E.; González-Porto, A.V.; García-Palencia, P.; Meana, A.; Del Nozal, M.J.; Mayo, R.; Bernal, J.L. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 2009, 1, 110–113. [Google Scholar] [CrossRef]
- Hidalgo, M.I.B.; Pacheco, M.L.; Pacheco, J. Origen floral de las cargas de polen recogidas por Apis mellifera L. en Álora (Málaga, España). Acta Bot. Malacit. 1990, 15, 33–44. [Google Scholar] [CrossRef]
- Louveaux, J.; Maurizio, A.; Vorwohl, G. Methods of Melissopalynology. Bee World 1978, 59, 139–157. [Google Scholar] [CrossRef]
- Erdtman, G. Morphology—Taxonomy—Ecology. An Introduction to the Study of Pollen Grains and Spores. In Handbook of Palynology; Munksgaard: Copenhagen, Denmark, 1969. [Google Scholar]
- Ohe, W.V.D.; Oddo, L.P.; Piana, M.L.; Morlot, M.; Martin, P. Harmonized methods of melissopalynology. Apidologie 2004, 35, S18–S25. [Google Scholar]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; John Wiley & Sons: Chichester, UK, 1989. [Google Scholar]
- Valdés, B.; Fernández, I.D.M.J. Atlas Polínico de Andalucía Occidental; Instituto de Desarrollo Regional de la Universidad de Sevilla y Excma, Diputación de Cádiz: Sevilla, Spain, 1987. [Google Scholar]
- Valverde, S.; Ares, A.M.; Bernal, J.L.; Nozal, M.J.; Bernal, J. Effect of the Storage Conditions (Light and Temperature) on the Detection of Thiamethoxam and Clothianidin Content in Rapeseeds by LC-DAD. Food Anal. Methods 2018, 11, 161–169. [Google Scholar] [CrossRef]
- Valverde, S.; Bernal, J.L.; Martín, M.T.; Nozal, M.J.; Bernal, J. Fast determination of neonicotinoid insecticides in bee pollen using QuEChERS and ultra-high performance liquid chromatography coupled to quadrupole time-of-flight mass spectrometry. Electrophoresis 2016, 37, 2470–2477. [Google Scholar] [CrossRef] [PubMed]
- Valverde, S.; Ibáñez, M.; Bernal, J.L.; Nozal, M.J.; Hernández, F.; Bernal, J. Development and validation of ultra high performance-liquid chromatography–tandem mass spectrometry based methods for the determination of neonicotinoid insecticides in honey. Food Chem. 2018, 266, 215–222. [Google Scholar] [CrossRef]
- Jara, L.; Ruiz, C.; Martín-Hernández, R.; Muñoz, I.; Higes, M.; Serrano, J.; De la Rúa, P. The Effect of Migratory Beekeeping on the Infestation Rate of Parasites in Honey Bee (Apis mellifera) Colonies and on Their Genetic Variability. Microorganisms 2021, 9, 22. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Botías, C.; Bailón, E.G.; Martínez-Salvador, A.; Prieto, L.; Meana, A.; Higes, M. Microsporidia infecting Apis mellifera: Coexistence or competition. Is Nosema ceranae replacing Nosema apis? Environ. Microbiol. 2012, 14, 2127–2138. [Google Scholar] [CrossRef]
- Chantawannakul, P.; Ward, L.; Boonham, N.; Brown, M. A scientific note on the detection of honeybee viruses using real-time PCR (TaqMan) in Varroa mites collected from a Thai honeybee (Apis mellifera) apiary. J. Invertebr. Pathol. 2006, 91, 69–73. [Google Scholar] [CrossRef]
- Francis, R.; Kryger, P. Single Assay Detection of Acute Bee Paralysis Virus, Kashmir Bee Virus and Israeli Acute Paralysis Virus. J. Apic. Sci. 2012, 56, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Medrzycki, P.; Giffard, H.; Aupinel, P.; Belzunces, L.P.; Chauzat, M.-P.; Claßen, C.; Colin, M.E.; Dupont, T.; Girolami, V.; Johnson, R.; et al. Standard methods for toxicology research in Apis mellifera. J. Apic. Res. 2013, 52, 1–60. [Google Scholar] [CrossRef] [Green Version]
- Pirk, C.W.W.; de Miranda, J.R.; Kramer, M.; Murray, T.E.; Nazzi, F.; Shutler, D.; van der Steen, J.J.M.; van Dooremalen, C. Statistical guidelines for Apis mellifera research. J. Apic. Res. 2013, 52, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Divine, G.W.; Norton, H.J.; Barón, A.E.; Juarez-Colunga, E. The Wilcoxon–Mann–Whitney Procedure Fails as a Test of Medians. Am. Stat. 2018, 72, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Fritz, C.O.; Morris, P.E.; Richler, J.J. Effect size estimates: Current use, calculations, and interpretation. J. Exp. Psychol. Gen. 2012, 141, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaiontz, C. Real Statistics Using Excel. Available online: www.real-statistics.com (accessed on 15 March 2021).
- Lenhard, W.; Lenhard, A. Calculation of Effect Sizes. Available online: https://www.psychometrica.de/effect_size.html (accessed on 6 June 2021).
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. The Use of Ranks to Avoid the Assumption of Normality Implicit in the Analysis of Variance. J. Am. Stat. Assoc. 1937, 32, 675–701. [Google Scholar] [CrossRef]
- Instituto Geológico y Minero de España. IGME_Geode_50 (MapServer): Continuous Digital Geological Map of Spain, Scale 1:50.000; Instituto Geológico y Minero de España: Madrid, Spain, 2014. [Google Scholar]
- World Reference Base, I.W.G. World Reference Base for Soil Resources 2014, Update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2015; Volume 106. [Google Scholar]
- Rivas-Martinez, S. Memoria del Mapa de Series de Vegetación de España; Serie Técnica; Ministerio de Agricultura, Pesca y Alimentación; I.C.O.N.A: Madrid, Spain, 1987. [Google Scholar]
- Nedić, N.; Mačukanović-Jocić, M.; Rančić, D.; Rørslett, B.; Šoštarić, I.; Stevanović, Z.D.; Mladenović, M. Melliferous potential of Brassica napus L. subsp. napus (Cruciferae). Arthropod-Plant Interact. 2013, 7, 323–333. [Google Scholar] [CrossRef]
- Talavera, S.; Gibbs, P.E.; Herrera, J. Reproductive biology of Cistus ladanifer (Cistaceae). Plant Syst. Evol. 1993, 186, 123–134. [Google Scholar] [CrossRef]
- Ortiz, P.L. The Cistaceae as food resources for honey bees in SW Spain. J. Apic. Res. 1994, 33, 136–144. [Google Scholar] [CrossRef]
- Atsalakis, E.; Chinou, I.; Makropoulou, M.; Karabournioti, S.; Graikou, K. Evaluation of Phenolic Compounds in Cistus creticus Bee Pollen from Greece. Antioxidant and Antimicrobial Properties. Nat. Prod. Commun. 2017, 12, 1934578X1701201141. [Google Scholar] [CrossRef] [Green Version]
- Papaefthimiou, D.; Papanikolaou, A.; Falara, V.; Givanoudi, S.; Kostas, S.; Kanellis, A.K. Genus Cistus: A model for exploring labdane-type diterpenes’ biosynthesis and a natural source of high value products with biological, aromatic, and pharmacological properties. Front. Chem. 2014, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Sponsler, D.B.; Johnson, R.M. Mechanistic modeling of pesticide exposure: The missing keystone of honey bee toxicology. Environ. Toxicol. Chem. 2017, 36, 871–881. [Google Scholar] [CrossRef]
- Huang, Z.-Y.; Robinson, G.E. Regulation of honey bee division of labor by colony age demography. Behav. Ecol. Sociobiol. 1996, 39, 147–158. [Google Scholar] [CrossRef]
- Schott, M.; Sandmann, M.; Cresswell, J.E.; Becher, M.A.; Eichner, G.; Brandt, D.T.; Halitschke, R.; Krueger, S.; Morlock, G.; Düring, R.-A.; et al. Honeybee colonies compensate for pesticide-induced effects on royal jelly composition and brood survival with increased brood production. Sci. Rep. 2021, 11, 62. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. The Hormetic Dose-Response Model Is More Common than the Threshold Model in Toxicology. Toxicol. Sci. 2003, 71, 246–250. [Google Scholar] [CrossRef]
- Luckey, T.D. Insecticide Hormoligosis. J. Econ. Entomol. 1968, 61, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Southam, C.M.; Ehrlich, J. Effects of Extract of western red-cedar heartwood on certain wood-decaying fungi in culture. Phytopathology 1943, 33, 517–524. [Google Scholar]
- Kendig, E.L.; Le, H.H.; Belcher, S.M. Defining hormesis: Evaluation of a complex concentration response phenomenon. Int. J. Toxicol. 2010, 29, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E. Pesticide-mediated homeostatic modulation in arthropods. Pestic. Biochem. Physiol. 2006, 85, 21–27. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Cutler, G.C. Insecticide-induced hormesis and arthropod pest management. Pest Manag. Sci. 2014, 70, 690–697. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J.; Blain, R. The occurrence of hormetic dose responses in the toxicological literature, the hormesis database: An overview. Toxicol. Appl. Pharm. 2005, 202, 289–301. [Google Scholar] [CrossRef]
- Costantini, D.; Metcalfe, N.B.; Monaghan, P. Ecological processes in a hormetic framework. Ecol. Lett. 2010, 13, 1435–1447. [Google Scholar] [CrossRef]
- Berry, R.; López-Martínez, G. A dose of experimental hormesis: When mild stress protects and improves animal performance. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 242, 110658. [Google Scholar] [CrossRef] [PubMed]
- Bryden, J.; Gill, R.J.; Mitton, R.A.A.; Raine, N.E.; Jansen, V.A.A. Chronic sublethal stress causes bee colony failure. Ecol. Lett. 2013, 16, 1463–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, C.J.; Søvik, E.; Myerscough, M.R.; Barron, A.B. Rapid behavioral maturation accelerates failure of stressed honey bee colonies. Proc. Natl. Acad. Sci. USA 2015, 112, 3427–3432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higes, M.; Meana, A.; Bartolomé, C.; Botías, C.; Martín-Hernández, R. Nosema ceranae (Microsporidia), a controversial 21st century honey bee pathogen. Environ. Microbiol. Rep. 2013, 5, 17–29. [Google Scholar] [CrossRef]
- Saitanis, C.J.; Agathokleous, E. Stress response and population dynamics: Is Allee effect hormesis? Sci. Total Environ. 2019, 682, 623–628. [Google Scholar] [CrossRef]
- Dennis, B.; Kemp, W.P. How Hives Collapse: Allee Effects, Ecological Resilience, and the Honey Bee. PLoS ONE 2016, 11, e0150055. [Google Scholar] [CrossRef] [Green Version]
- Courchamp, F.; Clutton-Brock, T.; Grenfell, B. Inverse density dependence and the Allee effect. Trends Ecol. Evol. 1999, 14, 405–410. [Google Scholar] [CrossRef]
- Ministerio de Agricultura, Pesca y Alimentación. Informes de Resultados del Programa de Vigilancia Sobre la Pérdida de Colonias de Abejas. Available online: https://www.mapa.gob.es/es/ganaderia/temas/sanidad-animal-higiene-ganadera/sanidad-animal/enfermedades/otras-enfermedades-abejas/otras-enf-abejas.aspx (accessed on 11 November 2021).
- Sadd, B.M.; Schmid-Hempel, P. Perspective: Principles of ecological immunology. Evol. Appl. 2009, 2, 113–121. [Google Scholar] [CrossRef]
- Liu, Z.; Li, S.; Li, H. Interactions between sublethal doses of thiamethoxam and Nosema ceranae in the honey bee, Apis mellifera. J. Apic. Res. 2021, 60, 717–725. [Google Scholar] [CrossRef]
- McKinstry, M.; Chung, C.; Truong, H.; Johnston, B.A.; Snow, J.W. The heat shock response and humoral immune response are mutually antagonistic in honey bees. Sci. Rep. 2017, 7, 8850. [Google Scholar] [CrossRef] [Green Version]
- Straub, L.; Williams, G.R.; Vidondo, B.; Khongphinitbunjong, K.; Retschnig, G.; Schneeberger, A.; Chantawannakul, P.; Dietemann, V.; Neumann, P. Neonicotinoids and ectoparasitic mites synergistically impact honeybees. Sci. Rep. 2019, 9, 8159. [Google Scholar] [CrossRef] [PubMed]
- Locke, B.; Semberg, E.; Forsgren, E.; de Miranda, J.R. Persistence of subclinical deformed wing virus infections in honeybees following Varroa mite removal and a bee population turnover. PLoS ONE 2017, 12, e0180910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möckel, N.; Gisder, S.; Genersch, E. Horizontal transmission of deformed wing virus: Pathological consequences in adult bees (Apis mellifera) depend on the transmission route. J. Gen. Virol. 2011, 92, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Potts, R.; Clarke, R.M.; Oldfield, S.E.; Wood, L.K.; de Ibarra, N.H.; Cresswell, J.E. The effect of dietary neonicotinoid pesticides on non-flight thermogenesis in worker bumble bees (Bombus terrestris). J. Insect Physiol. 2018, 104, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Tosi, S.; Démares, F.J.; Nicolson, S.W.; Medrzycki, P.; Pirk, C.W.W.; Human, H. Effects of a neonicotinoid pesticide on thermoregulation of African honey bees (Apis mellifera scutellata). J. Insect Physiol. 2016, 93, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordier, C.; Dechatre, H.; Suchail, S.; Peruzzi, M.; Soubeyrand, S.; Pioz, M.; Pélissier, M.; Crauser, D.; Conte, Y.L.; Alaux, C. Colony adaptive response to simulated heat waves and consequences at the individual level in honeybees (Apis mellifera). Sci. Rep. 2017, 7, 3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmon, A.; Peruzzi, M.; Le Conte, Y.; Alaux, C.; Pioz, M. Temperature-driven changes in viral loads in the honey bee Apis mellifera. J. Invertebr. Pathol. 2019, 160, 87–94. [Google Scholar] [CrossRef]
- McMenamin, A.J.; Daughenbaugh, K.F.; Flenniken, M.L. The Heat Shock Response in the Western Honey Bee (Apis mellifera) is Antiviral. Viruses 2020, 12, 245. [Google Scholar] [CrossRef] [Green Version]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef] [Green Version]
- Cutler, G.C.; Rix, R.R. Can poisons stimulate bees? Appreciating the potential of hormesis in bee–pesticide research. Pest Manag. Sci. 2015, 71, 1368–1370. [Google Scholar] [CrossRef]
- EFSA Scientific Committee. Statistical Significance and Biological Relevance. EFSA J. 2011, 9, 2372. [Google Scholar] [CrossRef]
- Authority, E.F.S.; Ippolito, A.; Focks, A.; Rundlöf, M.; Arce, A.; Marchesi, M.; Neri, F.M.; Rortais, A.; Szentes, C.; Auteri, D. Analysis of background variability of honey bee colony size. EFSA Supporting Publ. 2021, 18, 6518E. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D.; Sultan, M.; McFarlane, A.D.; Brewer, L. A large-scale field study examining effects of exposure to clothianidin seed-treated canola on honey bee colony health, development, and overwintering success. PeerJ 2014, 2, e652. [Google Scholar] [CrossRef] [PubMed]
- Flores, J.M.; Gámiz, V.; Gil-Lebrero, S.; Rodríguez, I.; Navas, F.J.; García-Valcárcel, A.I.; Cutillas, V.; Fernández-Alba, A.R.; Hernando, M.D. A three-year large scale study on the risk of honey bee colony exposure to blooming sunflowers grown from seeds treated with thiamethoxam and clothianidin neonicotinoids. Chemosphere 2021, 262, 127735. [Google Scholar] [CrossRef] [PubMed]
- Hernando, M.D.; Gámiz, V.; Gil-Lebrero, S.; Rodríguez, I.; García-Valcárcel, A.I.; Cutillas, V.; Fernández-Alba, A.R.; Flores, J.M. Viability of honeybee colonies exposed to sunflowers grown from seeds treated with the neonicotinoids thiamethoxam and clothianidin. Chemosphere 2018, 202, 609–617. [Google Scholar] [CrossRef]
- Rolke, D.; Fuchs, S.; Grünewald, B.; Gao, Z.; Blenau, W. Large-scale monitoring of effects of clothianidin-dressed oilseed rape seeds on pollinating insects in Northern Germany: Effects on honey bees (Apis mellifera). Ecotoxicology 2016, 25, 1648–1665. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef] [Green Version]
- Osterman, J.; Wintermantel, D.; Locke, B.; Jonsson, O.; Semberg, E.; Onorati, P.; Forsgren, E.; Rosenkranz, P.; Rahbek-Pedersen, T.; Bommarco, R.; et al. Clothianidin seed-treatment has no detectable negative impact on honeybee colonies and their pathogens. Nat. Commun. 2019, 10, 692. [Google Scholar] [CrossRef] [Green Version]

 and honey
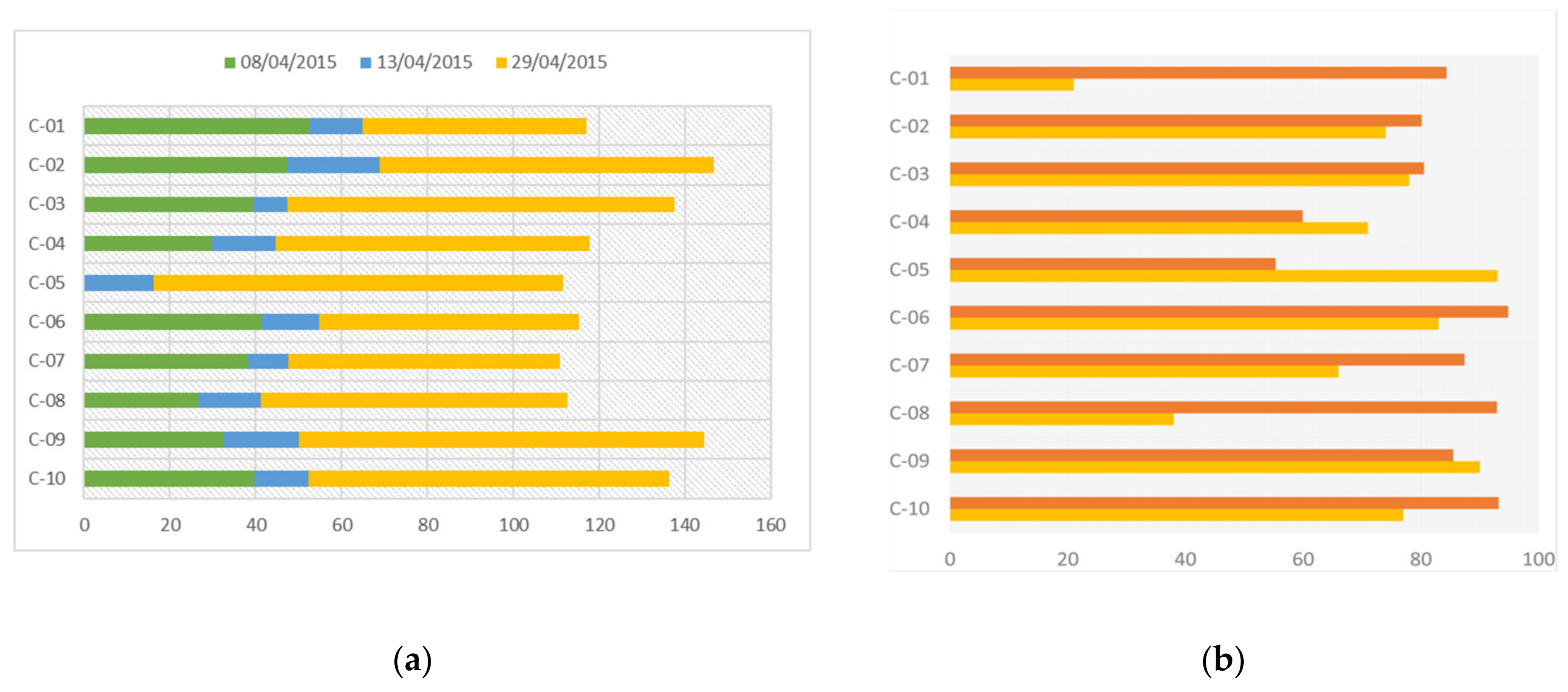
and honey  : C-01 to C-05 = Control hives; C-06 to C-10 = hives exposed to TMX.
and honey : C-01 to C-05 = Control hives; C-06 to C-10 = hives exposed to TMX.
: C-01 to C-05 = Control hives; C-06 to C-10 = hives exposed to TMX.
and honey : C-01 to C-05 = Control hives; C-06 to C-10 = hives exposed to TMX.

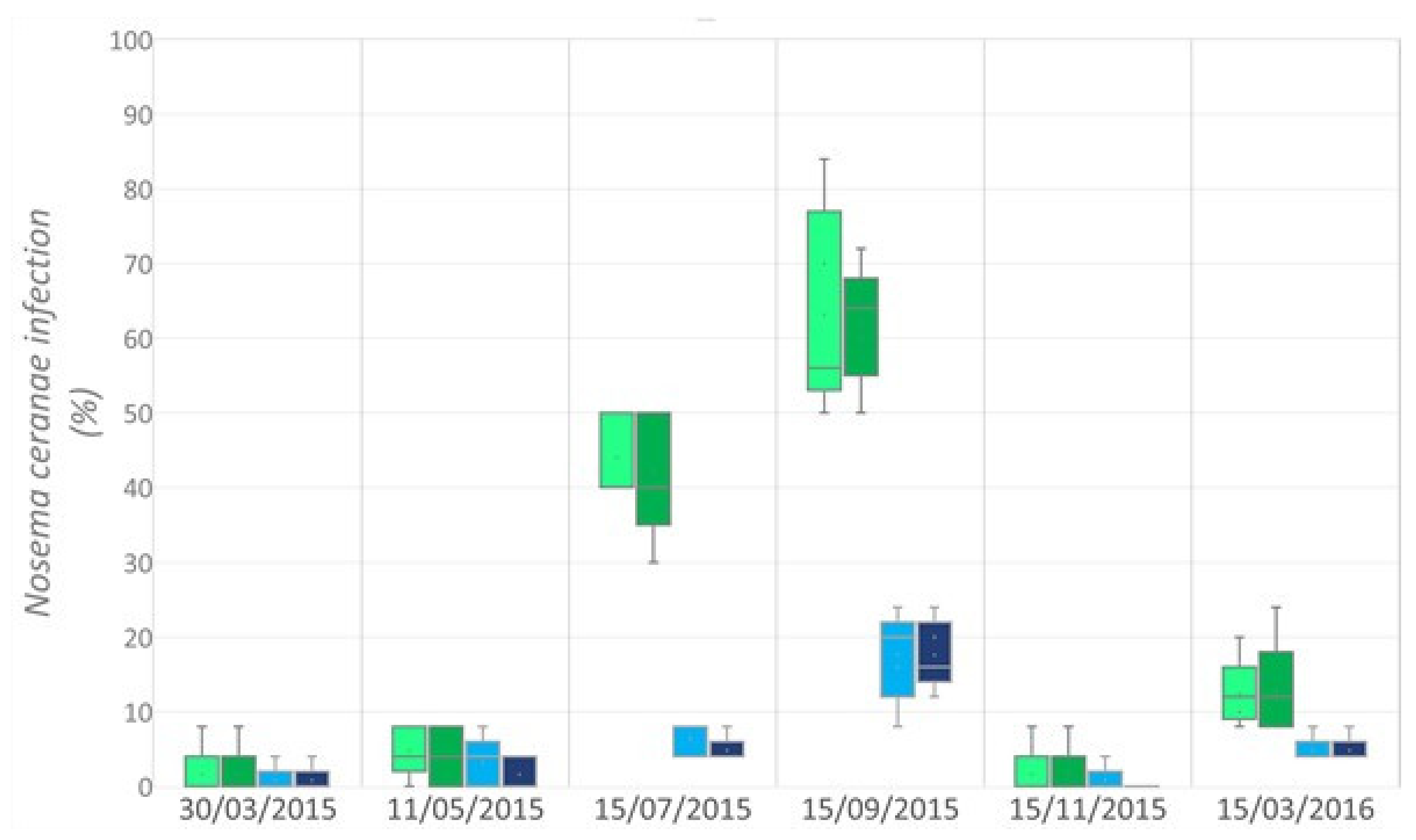
 control;
control;  TMX) and forager honey bees (FB;
TMX) and forager honey bees (FB;  control;
control;  TMX).
control; TMX) and forager honey bees (FB; control; TMX).
TMX).
control; TMX) and forager honey bees (FB; control; TMX).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Date | Forager Death | Colony Strength | Pathogens | Bee Pollen | Beebread | Honey |
|---|---|---|---|---|---|---|
| 30 March 2015 | X | X | X | |||
| 8 April 2015 | X | X | ||||
| 10 April 2015 | X | |||||
| 13 April 2015 | X | X | X | |||
| 15 April 2015 | X | |||||
| 17 April 2015 | X | |||||
| 20 April 2015 | X | |||||
| 22 April 2015 | X | |||||
| 24 April 2015 | X | |||||
| 27 April 2015 | X | |||||
| 29 April 2015 | X | X | ||||
| 4 May 2015 | X | |||||
| 6 May 2015 | X | |||||
| 8 May 2015 | X | |||||
| 11 May 2015 | X | X | X | X | X | |
| 15 July 2015 | X | X | ||||
| 15 September 2015 | X | X | ||||
| 17 September 2015 | ||||||
| 15 November 2015 | X | X | ||||
| 15 March 2016 | X | X |
| Variable | Descriptive Statistics | Control (n = 5) | TMX (n = 5) |
|---|---|---|---|
| Accumulated deaths | Min | 80 | 92 |
| Max | 192 | 394 | |
| Median | 129 | 214 | |
| Mean | 127.6 | 214.4 | |
| Standard deviation (SD) | 44.106 | 116.468 | |
| Variation coefficient (%) | 34.565 | 54.323 | |
| Mean 95% confidence interval | [72.836; 182.364] | [69.785; 359.015] | |
| Power (1-β) | 0.379 | ||
| Cohen’s d | 0.863 | ||
| Number of combs with worker bees | Min | 11 | 11 |
| Max | 14 | 16 | |
| Median | 12 | 16 | |
| Mean | 12.2 | 15 | |
| Standard deviation (SD) | 1.095 | 2.236 | |
| Mean 95% confidence interval | [10.84; 13.56] | [12.22; 17.77] | |
| Variation coefficient (%) | 8.979 | 14.907 | |
| Power (1-β) | 0.541 | ||
| Cohen’s d | 1.245 | ||
| Number of combs with brood | Min | 2 | 9 |
| Max | 13 | 13 | |
| Median | 10.5 | 11.0 | |
| Mean | 9.0 | 11.0 | |
| Standard deviation (SD) | 4.966 | 1.826 | |
| Variation coefficient (%) | 55.184 | 16.598 | |
| Mean 95% confidence interval | [1.097; 16.903] | [8.095; 13.905] | |
| Power (1-β) | 0.103 | ||
| Cohen’s d | 0.310 | ||
| Honey production (kg) | Min | 10.24 | 13.02 |
| Max | 45.22 | 45.58 | |
| Median | 16.204 | 32.034 | |
| Mean | 24.755 | 31.977 | |
| Standard deviation (SD) | 15.075 | 12.21 | |
| Variation coefficient (%) | 60.894 | 38.184 | |
| Mean 95% confidence interval | [6.038; 43.473] | [16.816; 47.137] | |
| Power (1-β) | 0.165 | ||
| Cohen’s d | 0.475 | ||
| Hive | 30 March 2015 | 11 May 2015 | 15 July 2015 | 15 September 2015 | 17 November 2015 | 15 March 2016 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IB | FB | IB | FB | IB | FB | IB | FB | IB | FB | IB | FB | |
| C 01 | BQCV | - | BQCV | BQCV | BQCV | BQCV | - | - | DWV | DWV | DWV | DWV |
| C 02 | BQCV | - | BQCV | - | BQCV | BQCV | BQCV | - | BQCV DWV | DWV | - | - |
| C 03 | BQCV | BQCV-DWV | BQCV | BQCV | BQCV | BQCV | - | BQCV | BQCV-DWV | DWV | - | - |
| C 04 | BQCV | BQCV-DWV | BQCV | DWV | BQCV | BQCV | BQCV | BQCV | BQCV-DWV | DWV | BQCV | - |
| C 05 | DWV | BQCV | BQCV-DWV | - | DWV | BQCV | BQCV | - | BQCV-DWV | DWV | BQCV | - |
| C 06 | BQCV | BQCV | BQCV | - | BQCV | BQCV | - | - | BQCV-DWV | DWV | BQCV | - |
| C 07 | BQCV | - | BQCV | - | BQCV | BQCV | - | BQCV | BQCV-DWV | DWV | BQCV | - |
| C 08 | BQCV | - | BQCV | BQCV | BQCV | BQCV | - | - | BQCV-DWV | DWV | - | DWV |
| C 09 | - | BQCV | - | - | BQCV | BQCV | BQCV | - | BQCV-DWV | DWV | - | - |
| C 10 | - | BQCV-DWV | - | BQCV | BQCV | BQCV | BQCV | - | BQCV-DWV | DWV | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso-Prados, E.; González-Porto, A.V.; García-Villarubia, C.; López-Pérez, J.A.; Valverde, S.; Bernal, J.; Martín-Hernández, R.; Higes, M. Effects of Thiamethoxam-Dressed Oilseed Rape Seeds and Nosema ceranae on Colonies of Apis mellifera iberiensis, L. under Field Conditions of Central Spain. Is Hormesis Playing a Role? Insects 2022, 13, 371. https://doi.org/10.3390/insects13040371
Alonso-Prados E, González-Porto AV, García-Villarubia C, López-Pérez JA, Valverde S, Bernal J, Martín-Hernández R, Higes M. Effects of Thiamethoxam-Dressed Oilseed Rape Seeds and Nosema ceranae on Colonies of Apis mellifera iberiensis, L. under Field Conditions of Central Spain. Is Hormesis Playing a Role? Insects. 2022; 13(4):371. https://doi.org/10.3390/insects13040371
Chicago/Turabian StyleAlonso-Prados, Elena, Amelia Virginia González-Porto, Carlos García-Villarubia, José Antonio López-Pérez, Silvia Valverde, José Bernal, Raquel Martín-Hernández, and Mariano Higes. 2022. "Effects of Thiamethoxam-Dressed Oilseed Rape Seeds and Nosema ceranae on Colonies of Apis mellifera iberiensis, L. under Field Conditions of Central Spain. Is Hormesis Playing a Role?" Insects 13, no. 4: 371. https://doi.org/10.3390/insects13040371
APA StyleAlonso-Prados, E., González-Porto, A. V., García-Villarubia, C., López-Pérez, J. A., Valverde, S., Bernal, J., Martín-Hernández, R., & Higes, M. (2022). Effects of Thiamethoxam-Dressed Oilseed Rape Seeds and Nosema ceranae on Colonies of Apis mellifera iberiensis, L. under Field Conditions of Central Spain. Is Hormesis Playing a Role? Insects, 13(4), 371. https://doi.org/10.3390/insects13040371