
A New Species in Notobitiella Hsiao in China Confirmed by Morphological and Molecular Data (Hemiptera: Heteroptera: Coreidae)




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
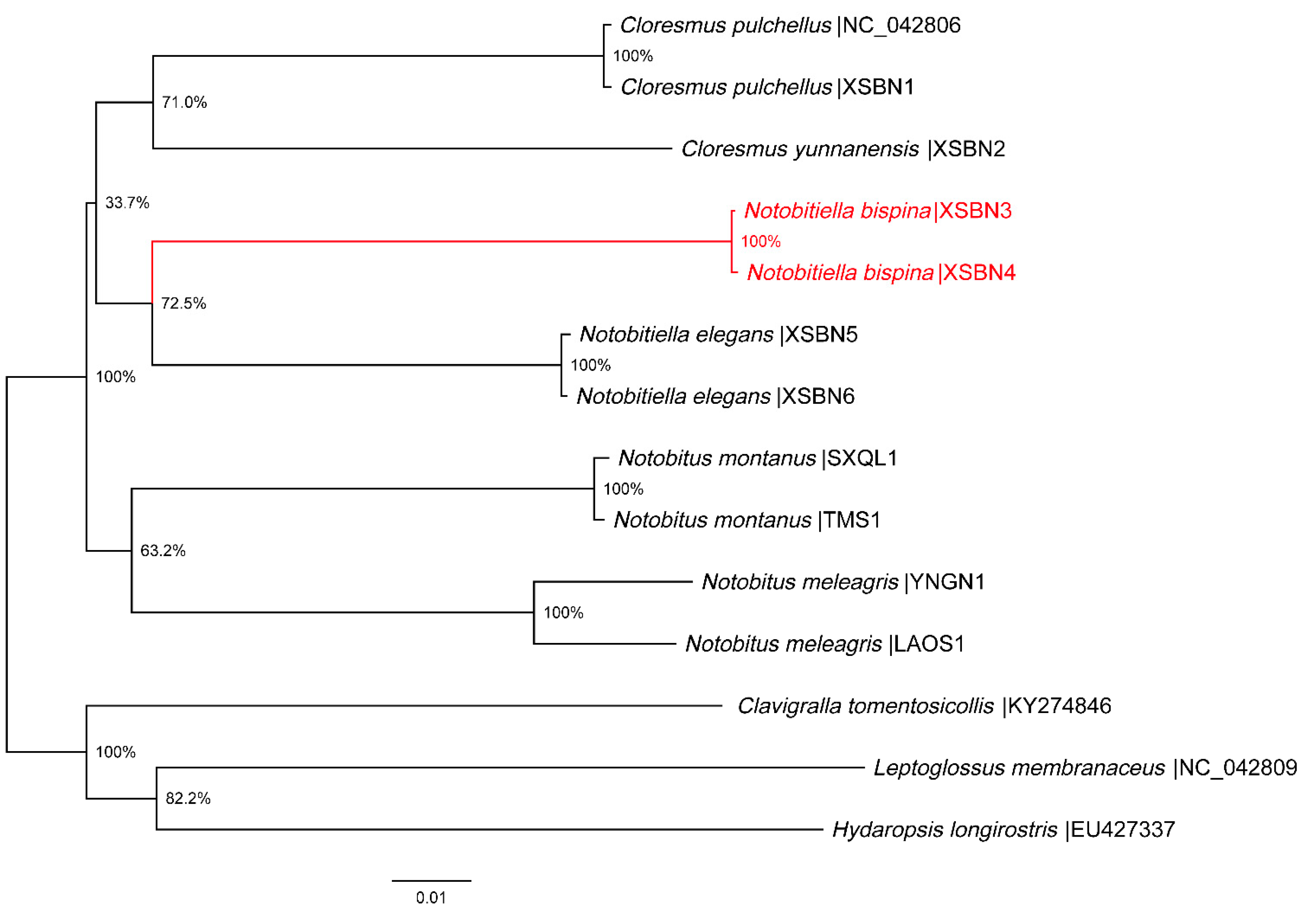
3. Results
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsiao, T.-Y. Result of the zoologico-botanical expedition to southwest China, 1955–1957 (Hemiptera, Coreidae). Acta Entomol. Sin. 1963, 12, 310–344. [Google Scholar]
- Zheng, L.-Y. Heteropteran Insects (Hemiptera) Feeding on Bamboos in China. Ann. Entomol. Soc. Am. 1994, 87, 91–96. [Google Scholar] [CrossRef]
- Hsiao, T.-Y. The Chinese bamboo Coreids of the tribe Cloresmini Stal with the description of a new subspecies (Hemiptera-Heteroptera). Acta Entomol. Sin. 1963, 12, 506–510. [Google Scholar]
- Khalaji-Pirbalouty, V.; Oraie, H.; Santamaria, C.A.; Wägele, J.W. Redescription of Tylos maindroni Giordani Soika, 1954 (Crustacea, Isopoda, Oniscidea) based on SEM and molecular data. ZooKeys 2022, 1087, 123–139. [Google Scholar] [CrossRef]
- Wang, S.; Bu, W. A new species in the genus Malcus Stål (Hemiptera: Heteroptera: Malcidae) from China based on morphological and molecular data. Zootaxa 2020, 4759, 252–257. [Google Scholar] [CrossRef]
- Krčmar, S.; Kučinić, M.; Pezzi, M.; Bruvo Mađarić, B. DNA barcoding of the horsefly fauna (Diptera, Tabanidae) of Croatia with notes on the morphology and taxonomy of selected species from Chrysopsinae and Tabaninae. ZooKeys 2022, 1087, 141–161. [Google Scholar] [CrossRef]
- Park, D.S.; Foottit, R.; Maw, E.; Hebert, P.D. Barcoding bugs: DNA-based identification of the true bugs (Insecta: Hemiptera: Heteroptera). PLoS ONE 2011, 6, e18749. [Google Scholar] [CrossRef]
- Raupach, M.J.; Hendrich, L.; Küchler, S.M.; Deister, F.; Morinière, J.; Gossner, M.M. Building-up of a DNA barcode library for true bugs (insecta: Hemiptera: Heteroptera) of Germany reveals taxonomic uncertainties and surprises. PLoS ONE 2014, 9, e106940. [Google Scholar] [CrossRef]
- Havemann, N.; Gossner, M.M.; Hendrich, L.; Morinière, J.; Niedringhaus, R.; Schäfer, P.; Raupach, M.J. From water striders to water bugs: The molecular diversity of aquatic Heteroptera (Gerromorpha, Nepomorpha) of Germany based on DNA barcodes. PeerJ 2018, 6, e4577. [Google Scholar] [CrossRef] [Green Version]
- Katz, A.D.; Giordano, R.; Soto-Adames, F.N. Operational criteria for cryptic species delimitation when evidence is limited, as exemplified by North American Entomobrya (Collembola: Entomobryidae). Zool. J. Linn. Soc. 2015, 173, 818–840. [Google Scholar] [CrossRef] [Green Version]
- Lefébure, T.; Douady, C.J.; Gouy, M.; Gibert, J. Relationship between morphological taxonomy and molecular divergence within Crustacea: Proposal of a molecular threshold to help species delimitation. Mol. Phylogenetics Evol. 2006, 40, 435–447. [Google Scholar] [CrossRef]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic. Acids. Res. 2019, 47, e63. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic. Acids. Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Song, F.; Zhao, Y.; Wilson, J.J.; Cai, W. Higher-level phylogeny and evolutionary history of Pentatomomorpha (Hemiptera: Heteroptera) inferred from mitochondrial genome sequences. Syst. Entomol. 2019, 44, 810–819. [Google Scholar] [CrossRef]
- Hua, J.; Li, M.; Dong, P.; Cui, Y.; Xie, Q.; Bu, W. Comparative and phylogenomic studies on the mitochondrial genomes of Pentatomomorpha (Insecta: Hemiptera: Heteroptera). BMC Genom. 2008, 9, 610. [Google Scholar] [CrossRef] [Green Version]
- Steele, L.D.; Sun, W.; Valero, M.C.; Ojo, J.A.; Seong, K.M.; Coates, B.S.; Margam, V.M.; Tamò, M.; Pittendrigh, B.R. The mitogenome of the brown pod-sucking bug Clavigralla tomentosicollis Stäl (Hemiptera: Coreidae). Agri. Gene. 2017, 5, 27–36. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Lin, X.L.; Mo, L.; Bu, W.J.; Wang, X.H. The first comprehensive DNA barcode reference library of Chinese Tanytarsus (Diptera: Chironomidae) for environmental DNA metabarcoding. Divers. Distrib. 2021, 27, 1932–1941. [Google Scholar] [CrossRef]
- Zheng, L.-Y.; Zou, H.-G. Records of Heteropterous insects on bamboos from Yunnan. Zool. Res. 1982, 3, 120–127. [Google Scholar]
- Brailovsky, H.; Barrera, E. A revision of the genus Priocnemicoris Costa with the description of eight new species (Hemiptera: Heteroptera: Coreidae: Cloresmini). Proc. Entomol. Soc. Wash. 2007, 109, 547–571. [Google Scholar]
- Brailovsky, H. Wasbauerellus, a new replacement name for Priocnemioides brailovsky, 2006 (Hemiptera: Heteroptera: Coreidae: Cloresmini). Proc. Entomol. Soc. Wash. 2007, 109, 957. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Date | Species Name | Location | Longitude | Latitude | Altitude | Collector |
|---|---|---|---|---|---|---|---|
| XSBN1 | 23 July 2021 | Cloresmus pulchellus | Haiwang road, Menga, Xishuangbanna, Yunnan, China | E100.390 | N22.046 | 1193 m | Juhong Chen |
| XSBN2 | 23 July 2021 | Cloresmus yunnanensis | Menghun, Menghai, Xishuangbanna, Yunnan, China | E100.419 | N21.869 | 931 m | Kun Jinag |
| XSBN3 | 23 July 2021 | Notobitiella bispina | Haiwang road, Menga, Xishuangbanna, Yunnan, China | E100.390 | N22.046 | 1193 m | Juhong Chen |
| XSBN4 | 23 July 2021 | Notobitiella bispina | Haiwang road, Menga, Xishuangbanna, Yunnan, China | E100.390 | N22.046 | 1193 m | Juhong Chen |
| XSBN5 | 22 July 2021 | Notobitiella elegans | Haiwang road, Menga, Xishuangbanna, Yunnan, China | E100.364 | N22.110 | 1071 m | Juhong Chen |
| XSBN6 | 23 July 2021 | Notobitiella elegans | Haiwang road, Menga, Xishuangbanna, Yunnan, China | E100.390 | N22.046 | 1193 m | Juhong Chen |
| YNGN1 | 9 August 2020 | Notobitus meleagris | Babao, Guangnan, Yunnan, China | E105.495 | N23.756 | 1046 m | Xue Dong |
| LAOS1 | 19 August 2019 | Notobitus meleagris | Nadana Village, Salavan, Laos | E106.384 | N15.717 | 189 m | Jiayue Zhou |
| SXQL1 | 25 June 2014 | Notobitus montanus | Zhonghe Village, Zuoshui, Shanxi, China | E109.401 | N33.569 | 850 m | Wenjun Bu |
| TMS1 | 26 July 2015 | Notobitus montanus | Tianmu mountain, Zhejiang, China | E119.455 | N30.329 | 784 m | Wenbo Yi |
| Species | Pairwise Genetic Distances | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cloresmus pulchellus|NC_042806 | |||||||||||||
| Cloresmus pulchellus|XSBN1 | 0.0020 | ||||||||||||
| Cloresmus yunnanensis|XSBN2 | 0.1230 | 0.1230 | |||||||||||
| Notobitiella bispina|XSBN3 | 0.1496 | 0.1496 | 0.1447 | ||||||||||
| Notobitiella bispina|XSBN4 | 0.1513 | 0.1513 | 0.1455 | 0.0013 | |||||||||
| Notobitiella elegans|XSBN5 | 0.1246 | 0.1238 | 0.1354 | 0.1264 | 0.1256 | ||||||||
| Notobitiella elegans|XSBN6 | 0.1238 | 0.1230 | 0.1354 | 0.1264 | 0.1256 | 0.0020 | |||||||
| Notobitus meleagris|YNGN1 | 0.1349 | 0.1358 | 0.1566 | 0.1466 | 0.1475 | 0.1413 | 0.1413 | ||||||
| Notobitus meleagris|LAOS1 | 0.1300 | 0.1308 | 0.1582 | 0.1526 | 0.1517 | 0.1404 | 0.1404 | 0.0379 | |||||
| Notobitus montanus|SXQL1 | 0.1320 | 0.1320 | 0.1403 | 0.1542 | 0.1542 | 0.1227 | 0.1227 | 0.1338 | 0.1240 | ||||
| Notobitus montanus|TMS1 | 0.1320 | 0.1320 | 0.1403 | 0.1517 | 0.1517 | 0.1212 | 0.1212 | 0.1355 | 0.1240 | 0.0033 | |||
| L. membranaceus|NC_042809 | 0.1954 | 0.1954 | 0.1848 | 0.1971 | 0.1980 | 0.1782 | 0.1773 | 0.1937 | 0.1954 | 0.1780 | 0.1771 | ||
| C. tomentosicollis|KY274846 | 0.1635 | 0.1635 | 0.1689 | 0.1886 | 0.1886 | 0.1558 | 0.1550 | 0.1791 | 0.1791 | 0.1679 | 0.1687 | 0.1817 | |
| H. longirostris|EU427337 | 0.1878 | 0.1887 | 0.1853 | 0.1951 | 0.1960 | 0.1767 | 0.1758 | 0.1892 | 0.1805 | 0.1743 | 0.1726 | 0.1730 | 0.1691 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, K.; Chen, J.; Bu, W. A New Species in Notobitiella Hsiao in China Confirmed by Morphological and Molecular Data (Hemiptera: Heteroptera: Coreidae). Insects 2022, 13, 411. https://doi.org/10.3390/insects13050411
Jiang K, Chen J, Bu W. A New Species in Notobitiella Hsiao in China Confirmed by Morphological and Molecular Data (Hemiptera: Heteroptera: Coreidae). Insects. 2022; 13(5):411. https://doi.org/10.3390/insects13050411
Chicago/Turabian StyleJiang, Kun, Juhong Chen, and Wenjun Bu. 2022. "A New Species in Notobitiella Hsiao in China Confirmed by Morphological and Molecular Data (Hemiptera: Heteroptera: Coreidae)" Insects 13, no. 5: 411. https://doi.org/10.3390/insects13050411
APA StyleJiang, K., Chen, J., & Bu, W. (2022). A New Species in Notobitiella Hsiao in China Confirmed by Morphological and Molecular Data (Hemiptera: Heteroptera: Coreidae). Insects, 13(5), 411. https://doi.org/10.3390/insects13050411