
Somatic Mobilization: High Somatic Insertion Rate of mariner Transposable Element in Drosophila simulans


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
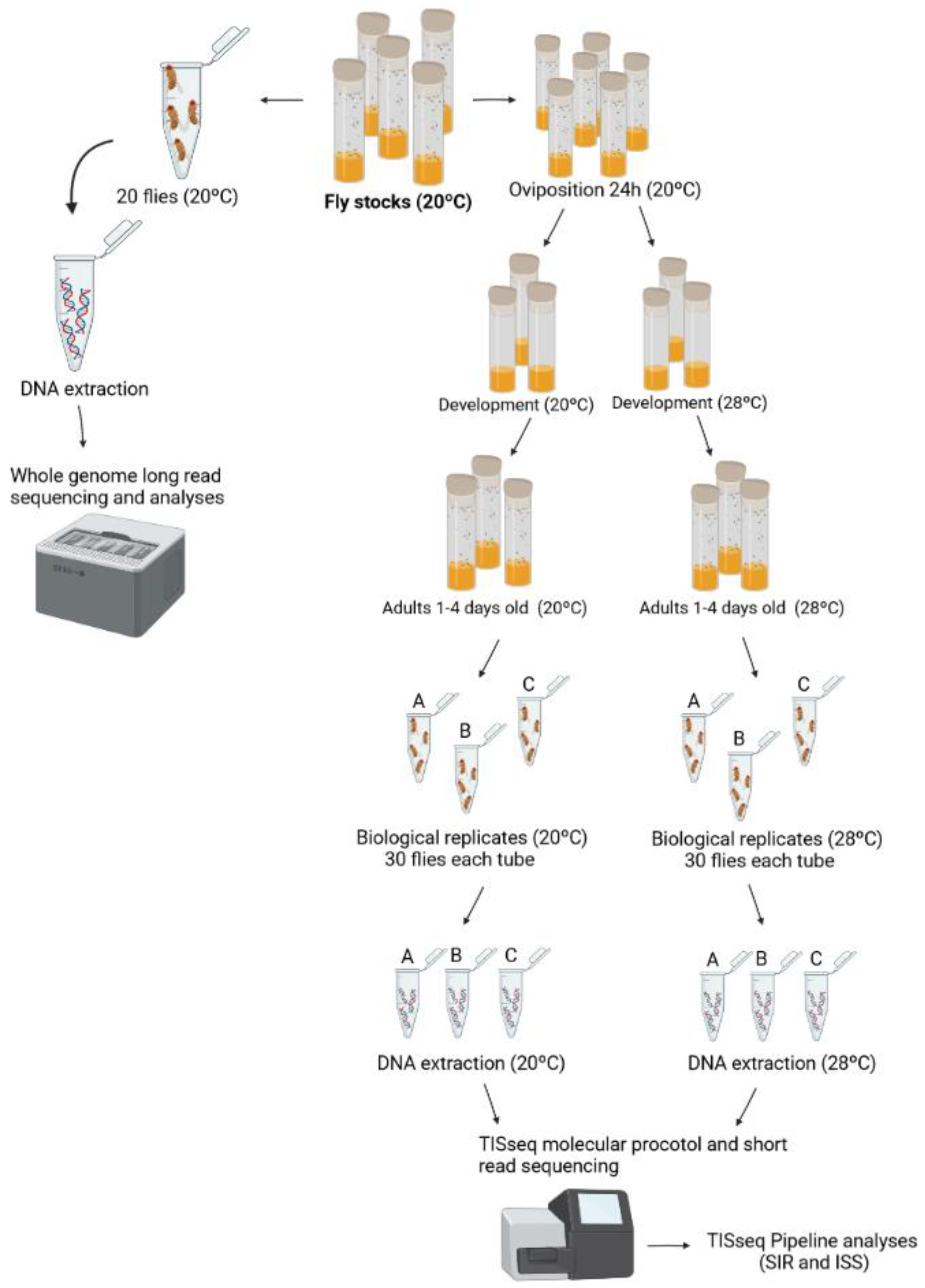
2.1. Drosophila Strain Used
2.2. Biological Treatments and DNA Extraction
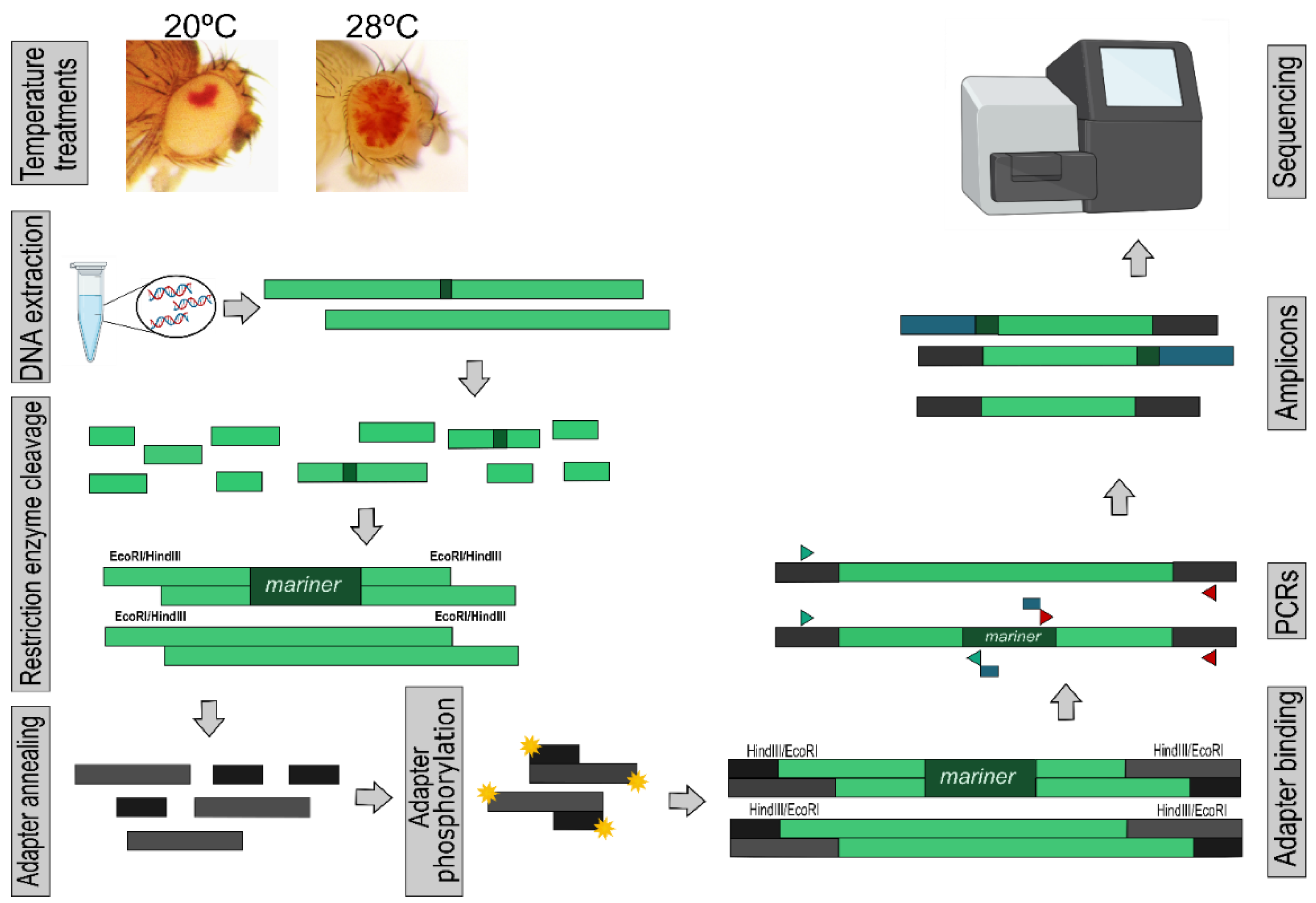
2.3. Transposon Insertion Site Sequencing (TISseq)
2.4. Bioinformatics Analysis
2.5. Somatic Insertion Rate Estimation (SIR)
2.6. Whole-Genome Sequencing and Analyses
3. Results
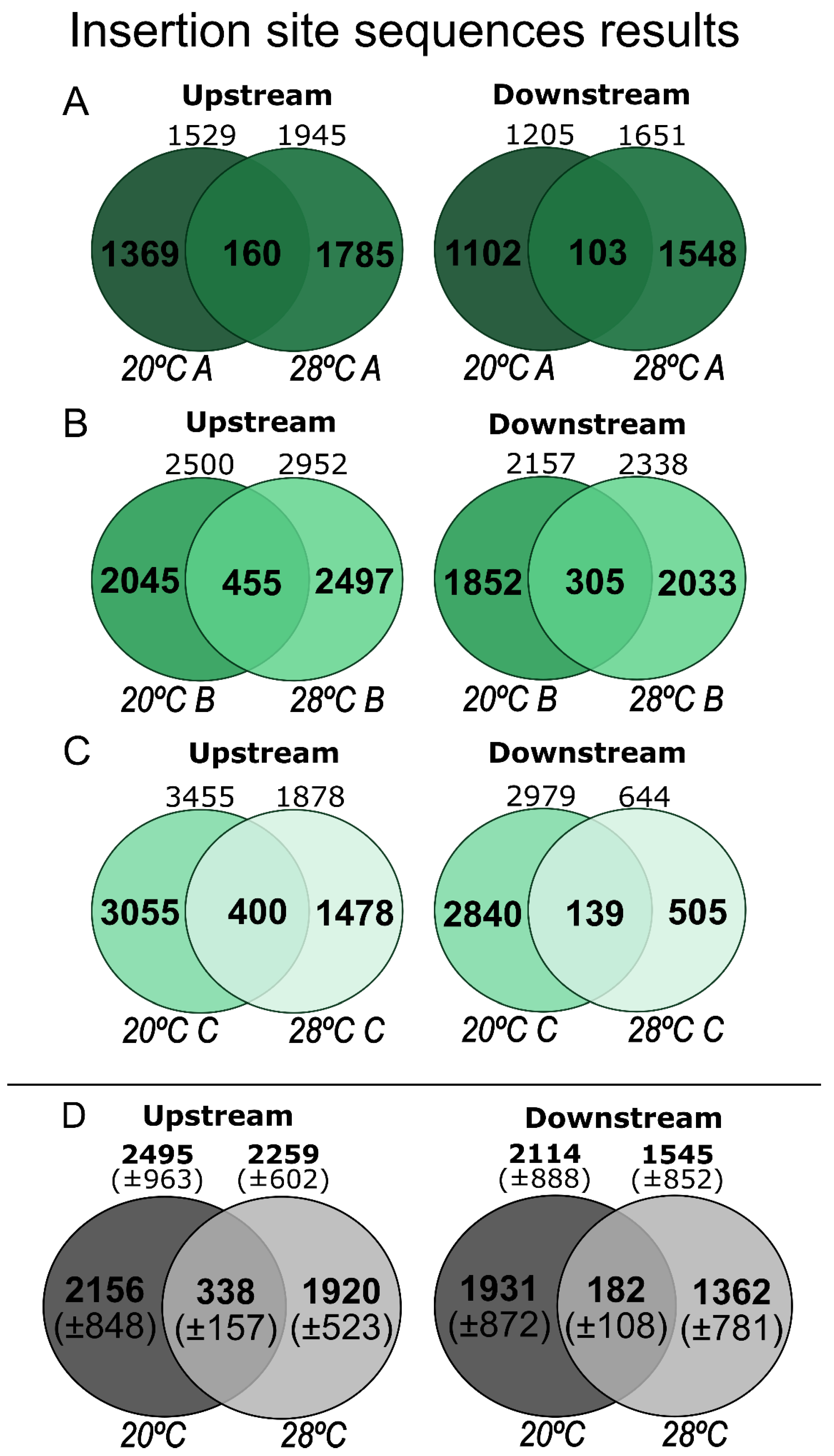
3.1. ISS and SIR Estimation
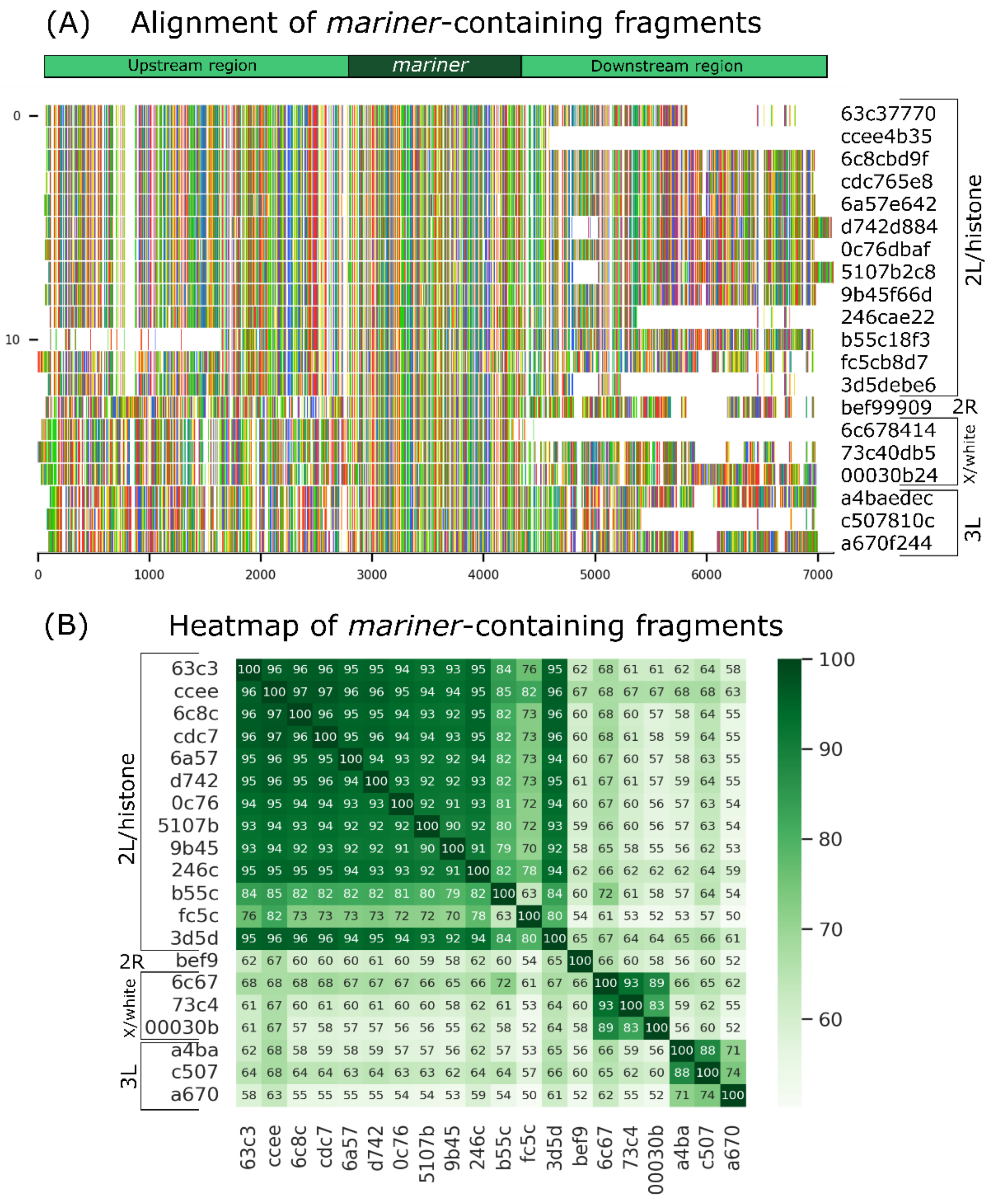
3.2. Analysis of Long-Read Fragments
4. Discussion
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kidwell, M.G.; Lisch, D.R. Perspective: Transposable elements, parasitic DNA, and genome evolution. Evolution 2001, 55, 1–24. [Google Scholar] [CrossRef]
- Diesel, J.F.; Ortiz, M.F.; Marinotti, O.; Vasconcelos, A.T.R.; Elgion, L.S. A re-annotation of the Anopheles darlingi mobilome. Genet. Mol. Biol. 2019, 131, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biémont, C. A brief history of the status of transposable elements: From junk DNA to major players in evolution. Genetics 2010, 186, 1085–1093. [Google Scholar] [CrossRef] [Green Version]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laricchia, K.M.; Zdraljevic, S.; Cook, D.E.; Andersen, E.C. Natural Variation in the Distribution and Abundance of Transposable Elements Across the Caenorhabditis elegans Species. Mol. Biol. Evol. 2017, 34, 2187–2202. [Google Scholar] [CrossRef]
- Nishihara, H. Transposable elements as genetic accelerators of evolution: Contribution to genome size, gene regulatory network rewiring and morphological innovation. Genes Genet. Syst. 2019, 94, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Chénais, B.; Caruso, A.; Hiard, S.; Casse, N. The impact of transposable elements on eukaryotic genomes: From genome size increase to genetic adaptation to stressful environments. Gene 2012, 509, 7–15. [Google Scholar] [CrossRef]
- Wicker, T.; Sabot, F.; Hua-van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef]
- Lapp, H.E.; Hunter, R.G. Early life exposures, neurodevelopmental disorders, and transposable elements. Neurobiol. Stress 2019, 11, 100174. [Google Scholar] [CrossRef]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef]
- Hua-Van, A.; Le Rouzic, A.; Boutin, T.S.; Filée, J.; Capy, P. The struggle for life of the genome’s selfish architects. Biol. Direct 2011, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.G.; Jones, B.C.; Jiang, N.; Chang, C.; Hosier, S.; Wickremesinghe, P.; Garcia, M.; Hartnett, D.A.; Burhenn, L.; Neretti, N.; et al. Chromatin-modifying genetic interventions suppress age-associated transposable element activation and extend life span in Drosophila. Proc. Natl. Acad. Sci. USA 2016, 113, 11277–11282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czech, B.; Hannon, G.J. Small RNA sorting: Matchmaking for argonautes. Nat. Rev. Genet. 2011, 12, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Levin, H.L.; Moran, J.V.; Shriver, E.K. Dynamic interactions between transposable elements and their hosts. Nat. Rev. Genet. 2011, 12, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, P.; Richardson, S.R.; Mager, D.L.; Faulkner, G.J. Transposable elements in the mammalian embryo: Pioneers surviving through stealth and service. Genome Biol. 2016, 17, 100. [Google Scholar] [CrossRef] [Green Version]
- Haig, D. Transposable elements: Self-seekers of the germline, team-players of the soma. BioEssays 2016, 38, 1158–1166. [Google Scholar] [CrossRef]
- Kazazian, H.H. Mobile DNA transposition in somatic cells. BMC Biol. 2011, 9, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Loreto, E.L.S.; Pereira, C.M. Somatizing the transposons action. Mob. Genet. Elem. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Miki, Y.; Nishisho, I.; Horii, A.; Miyoshi, Y.; Utsunomiya, J.; Kinzler, K.W.; Vogelstein, B.; Nakamura, Y. Disruption of the APC Gene by a Retrotransposal Insertion of LI Sequence in a Colon Cancer. Cancer Res. 1992, 52, 643–645. [Google Scholar]
- Rodić, N.; Steranka, J.P.; Makohon-Moore, A.; Moyer, A.; Shen, P.; Sharma, R.; Kohutek, Z.A.; Huang, C.R.; Ahn, D.; Mita, P.; et al. Retrotransposon insertions in the clonal evolution of pancreatic ductal adenocarcinoma. Nat. Med. 2015, 21, 1060–1064. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.S.; Shah, N.M.; Du, A.Y.; Dailey, Z.Z.; Pehrsson, E.C.; Godoy, P.M.; Zhang, D.; Li, D.; Xing, X.; Kim, S.; et al. Transposable elements drive widespread expression of oncogenes in human cancers. Nat. Genet. 2019, 51, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Mérel, V.; Boulesteix, M.; Fablet, M.; Vieira, C. Transposable elements in Drosophila. Mob. DNA 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Siudeja, K.; Beek, M.; Riddiford, N.; Boumard, B.; Wurmser, A.; Stefanutti, M.; Lameiras, S.; Bardin, A.J. Unraveling the features of somatic transposition in the Drosophila intestine. EMBO J. 2021, 40, 93. [Google Scholar] [CrossRef] [PubMed]
- Engels, W.R.; Benz, W.K.; Preston, C.R.; Graham, P.L.; Phillis, R.W.; Robertson, H.M. Somatic effects of P element activity in Drosophila melanogaster: Pupal lethality. Genetics 1987, 117, 745–757. [Google Scholar] [CrossRef]
- Jacobson, J.W.; Medhora, M.M.; Hartl, D.L. Molecular structure of a somatically unstable transposable element in Drosophila. Proc. Natl. Acad. Sci. USA. 1986, 83, 8684–8688. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, D.T.; Sullivan, M.C. Transport defects as the physiological basis for eye color mutants of Drosophila melanogaster. Biochem. Genet. 1975, 13, 603–613. [Google Scholar] [CrossRef]
- Sullivan, D.T.; Bell, L.A.; Paton, D.R.; Sullivan, M.C. Purine transport by Malpighian tubules of pteridine-deficient eye color mutants of Drosophila melanogaster. Biochem. Genet. 1979, 17, 565–573. [Google Scholar] [CrossRef]
- Ewart, G.D.; Howells, A.J. ABC transporters involved in transport of eye pigment precursors in Drosophila melanogaster. Method. Enzymol. 1998, 292, 213–224. [Google Scholar] [CrossRef]
- Haymer, D.S.; Marsh, J.L. Germ line and somatic instability of a white mutation in Drosophila mauritiana due to a transposable genetic element. Dev. Genet. 1985, 6, 281–291. [Google Scholar] [CrossRef]
- Bryan, G.J.; Jacobson, J.W.; Hartl, D.L. Heritable somatic excision of a Drosophila transposon. Science 1987, 235, 1636–1638. [Google Scholar] [CrossRef]
- Giraud, T.; Capy, P. Somatic activity of the mariner transposable element in natural populations of Drosophila simulans. Proc. R. Soc. B Biol. Sci. 1996, 263, 1481–1486. [Google Scholar] [CrossRef]
- Russell, A.L.; Woodruff, R.C. The genetics and evolution of the mariner transposable element in Drosophila simulans: Worldwide distribution and experimental population dynamics. Genetica 1999, 105, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Picot, S.; Wallau, G.L.; Loreto, E.L.S.; Heredia, F.O.; Hua-Van, A.; Capy, P. The mariner transposable element in natural populations of Drosophila simulans. Heredity 2008, 101, 53–59. [Google Scholar] [CrossRef]
- Jardim, S.S.; Schuch, A.P.; Pereira, C.M.; Loreto, E.L.S. Effects of heat and UV radiation on the mobilization of transposon mariner-Mos1. Cell Stress Chaperones 2015, 20, 843–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, C.M.; Stoffel, T.J.R.; Callegari-Jacques, S.M.; Hua-Van, A.; Capy, P.; Loreto, E.L.S. The somatic mobilization of transposable element mariner-Mos1 during the Drosophila lifespan and its biological consequences. Gene 2018, 679, 65–72. [Google Scholar] [CrossRef]
- Bryan, G.; Garza, D.; Hart, D. Insertion and Excision of the Transposable Element. Genetics 1990, 125, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Isolation of high-molecular-weight DNA using organic solvents. Cold Spring Harb. Protoc. 2017, 2017, 356–359. [Google Scholar] [CrossRef] [Green Version]
- Steranka, J.P.; Tang, Z.; Grivainis, M.; Huang, C.R.L.; Payer, L.M.; Rego, F.O.R.; Miller, T.L.A.; Galante, P.A.F.; Ramaswami, S.; Heguy, A.; et al. Transposon insertion profiling by sequencing (TIPseq) for mapping LINE-1 insertions in the human genome. Mob. DNA 2019, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Kircher, M.; Heyn, P.; Kelso, J. Addressing challenges in the production and analysis of illumina sequencing data. BMC Genom. 2011, 12, 382. [Google Scholar] [CrossRef] [Green Version]
- Head, S.R.; Kiyomi Komori, H.; LaMere, S.A.; Whisenant, T.; Van Nieuwerburgh, F.; Salomon, D.R.; Ordoukhanian, P. Library construction for next-generation sequencing: Overviews and challenges. Biotechniques 2014, 56, 61–77. [Google Scholar] [CrossRef] [Green Version]
- Burns, K.H. Transposable elements in cancer. Nat. Rev. Cancer 2017, 17, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Jeong, H.H.; Hsieh, Y.C.; Klein, H.U.; Bennett, D.A.; De Jager, P.L.; Liu, Z.; Shulman, J.M. Tau Activates Transposable Elements in Alzheimer’s Disease. Cell Rep. 2018, 23, 2874–2880. [Google Scholar] [CrossRef] [PubMed]
- Sedivy, J.M.; Kreiling, J.A.; Neretti, N.; Cecco, M.D.; Criscione, S.W.; Hofmann, J.W.; Zhao, X.; Ito, T.; Peterson, A.L. Death by transposition–The enemy within? BioEssays 2013, 35, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturm, Á.; Ivics, Z.; Vellai, T. The mechanism of ageing: Primary role of transposable elements in genome disintegration. Cell. Mol. Life Sci. 2015, 72, 1839–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturm, Á.; Perczel, A.; Ivics, Z.; Vellai, T. The Piwi-piRNA pathway: Road to immortality. Aging Cell 2017, 16, 906–911. [Google Scholar] [CrossRef]
- Li, W.; Prazak, L.; Chatterjee, N.; Grüninger, S.; Krug, L.; Theodorou, D.; Dubnau, J. Activation of transposable elements during aging and neuronal decline in Drosophila. Nat. Neurosci. 2013, 16, 529–531. [Google Scholar] [CrossRef] [Green Version]
- Treiber, C.D.; Waddell, S. Resolving the prevalence of somatic transposition in drosophila. Elife 2017, 6, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Huang, X.; Dou, S.; Tang, X.; Luo, S.; Theurkauf, W.E.; Lu, J.; Weng, Z. A benchmark and an algorithm for detecting germline transposon insertions and measuring de novo transposon insertion frequencies. Nucleic Acids Res. 2021, 49, E44. [Google Scholar] [CrossRef]
- Komkov, A.Y.; Minervina, A.A.; Nugmanov, G.A.; Saliutina, M.V.; Khodosevich, K.V.; Lebedev, Y.B.; Mamedov, I.Z. An advanced enrichment method for rare somatic retroelement insertions sequencing. Mob. DNA 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Daron, J.; Slotkin, R.K. EpiTEome: Simultaneous detection of transposable element insertion sites and their DNA methylation levels. Genome Biol. 2017, 18, 91. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fragments Trimmed (N) | Without Coverage | With Coverage | With + Without Coverage | Insertion Detected per Genome | Insertion Estimated per Genome | |
|---|---|---|---|---|---|---|
| Upstream 20 °C | 3533 ±1431.5 | 2164.6 ±780.7 | 330 ±183.9 | 2494.6 ±963.0 | 83.1 ±32.1 | 279.0 ±107.7 |
| Upstream 28 °C | 3155 ±790.5 | 1904.6 ±558.8 | 353.6 ±167.3 | 2258.3 ±601.6 | 75.3 ±20.0 | 252.6 ±67.3 |
| Downstream 20 °C | 2866.7 ±1186.6 | 1853.7 ±747.5 | 260 ±142.7 | 2113.6 ±887.8 | 70.4 ±29.6 | 236.4 ±99.3 |
| Downstream 28 °C | 1967.7 ±1043.9 | 1397 ±793.2 | 147.3 ±70.9 | 1544.3 ±852.0 | 51.5 ±28.4 | 172.7 ±95.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cancian, M.; da Fontoura Gomes, T.M.F.; Loreto, E.L.S. Somatic Mobilization: High Somatic Insertion Rate of mariner Transposable Element in Drosophila simulans. Insects 2022, 13, 454. https://doi.org/10.3390/insects13050454
Cancian M, da Fontoura Gomes TMF, Loreto ELS. Somatic Mobilization: High Somatic Insertion Rate of mariner Transposable Element in Drosophila simulans. Insects. 2022; 13(5):454. https://doi.org/10.3390/insects13050454
Chicago/Turabian StyleCancian, Mariana, Tiago Minuzzi Freire da Fontoura Gomes, and Elgion Lucio Silva Loreto. 2022. "Somatic Mobilization: High Somatic Insertion Rate of mariner Transposable Element in Drosophila simulans" Insects 13, no. 5: 454. https://doi.org/10.3390/insects13050454
APA StyleCancian, M., da Fontoura Gomes, T. M. F., & Loreto, E. L. S. (2022). Somatic Mobilization: High Somatic Insertion Rate of mariner Transposable Element in Drosophila simulans. Insects, 13(5), 454. https://doi.org/10.3390/insects13050454