1. Introduction
The Tenebrionoidea constitute one of the largest and most complex superfamilies of beetles [
1,
2]. A molecular study on the superfamily suggested that it is monophyletic and that four clades have been suggested within the group; among these clades, ripiphorid-mordellid-meloid were considered the most basal in the superfamily [
3]. Likewise, Bocak et al. [
4], also based on molecular data, considered these three families to be closely related, however in their study the clade formed by them occupies the most derived position in relation to the other Tenebrionoidea. On the other hand, studies by Zhang et al. [
5] and McKenna et al. [
6], based on extensive gene sampling, maintained Ripiphoridae and Mordellidae as a sister group and in a more basal position of the tenebrionoid tree, while Meloidae appears in a higher position in this tree.
The structure of tenebrionoid sperm is known from Baccetti et al. [
7], Dias et al. [
8,
9], Dallai [
10], and Folly et al. [
11]. These works have well established that within this group of beetles, the sperm are characterized by a short acrosome, a cylindrical nucleus, and a flagellum with a 9 + 9 + 2 axoneme flanked by two long mitochondrial derivatives and two cylindrical or elliptical accessory bodies [
9]. This model, however, is variable in the different families, mainly due to the shape of accessory bodies. In most beetles, testicular sperm bundles, formed at the end of spermatogenesis by cell divisions, contain up to 256 (2
8) cells. In Tenebrionoidea, this number usually rises to 512 (2
9), but there are species where this number is only 64 (2
6), and there are also species where bundles contain 1024 (2
10) spermatozoa.
Different morphological cellular mechanisms along the spermatogenic process in insects have shown a source of variability in sperm arrangements within cyst cells [
12,
13]. According to Dias et al. [
8], one characteristic shared by members of several families of Tenebrionoidea is that the spermatozoa do not maintain a single orientation within the cyst, as it usually occurs in insects. Nevertheless, during spermiogenesis, their nuclei migrate towards two opposite poles of the cyst, forming two sets of sperm with antiparallel orientation. Also, a unique spermatogenesis mechanism was described for the Hemiptera
Planococcus citri (Pseudococcidae) [
14] and
Kerria chinensis (Kerriidae) [
12]. In this, the result is two sperm bundles separated at the end of spermiogenesis from a process of inverted meiosis, i.e., by a mechanism different from those tenebrionoids described so far. Studying the sperm ultrastructure of some members of three families considered basal [
9], it was concluded that Ripiphoridae and Mordellidae have a close phylogenetic relationship. In contrast, Meloidae would be better placed in a more advanced position in the superfamily, as was also suggested, based on molecular data, by Zhang et al. [
5] and McKenna et al. [
6]. The present work aims to improve our knowledge of tenebrionid sperm structure, extending the study to other group families. In particular, we have examined two new members of Mordellidae, a member of Oedemeridae and one of Tenebrionidae. The results obtained confirm our previous conclusions [
9] and give details on a peculiar process, the sperm looping [
13], occurring in the testicular cysts in Mordellidae, allowing the sperm to compact in testes of reduced size.
2. Materials and Methods
The following species were studied in the present work:
Mordellidae:
Mordellistena (Mordellistena) brevicauda (Boheman, 1849). Campiglia d’Orcia, Siena, Italy. 12 ex.
Hoshihananomia sp. (ex Machairophora sensu Franciscolo, 1943). Viçosa, MG, Brazil. 4 ex.
Oedemeridae:
Oedemera nobilis (Scopoli, 1763). Gallina, Siena, Italy. numerous ex.
Tenebrionidae:
Accanthopus velikensis (Piller & Mitterpacher, 1783). Monte Amiata, Siena, Italy. 2 ex.
2.1. Light and Epifluorescence Microscopic Preparations
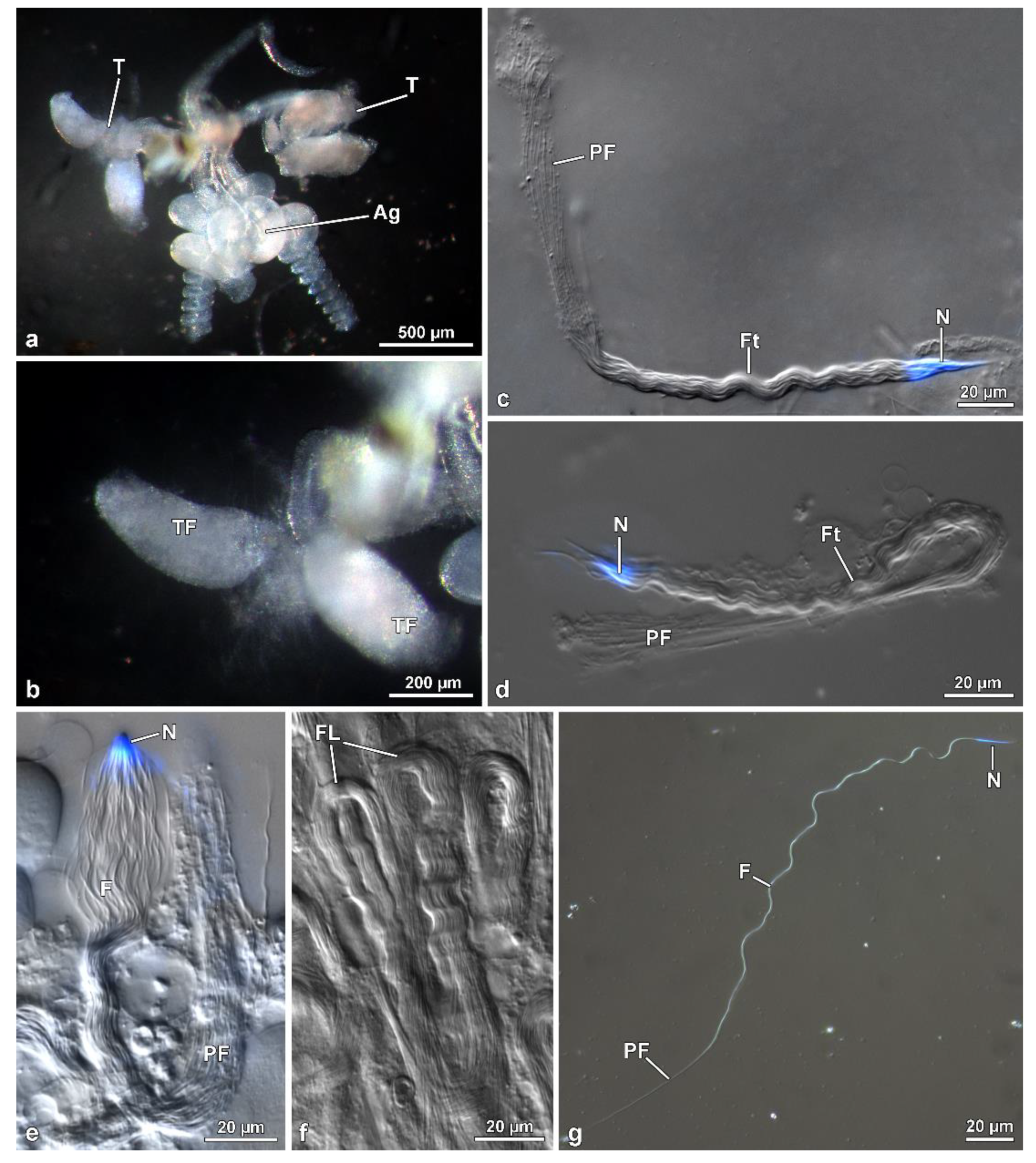
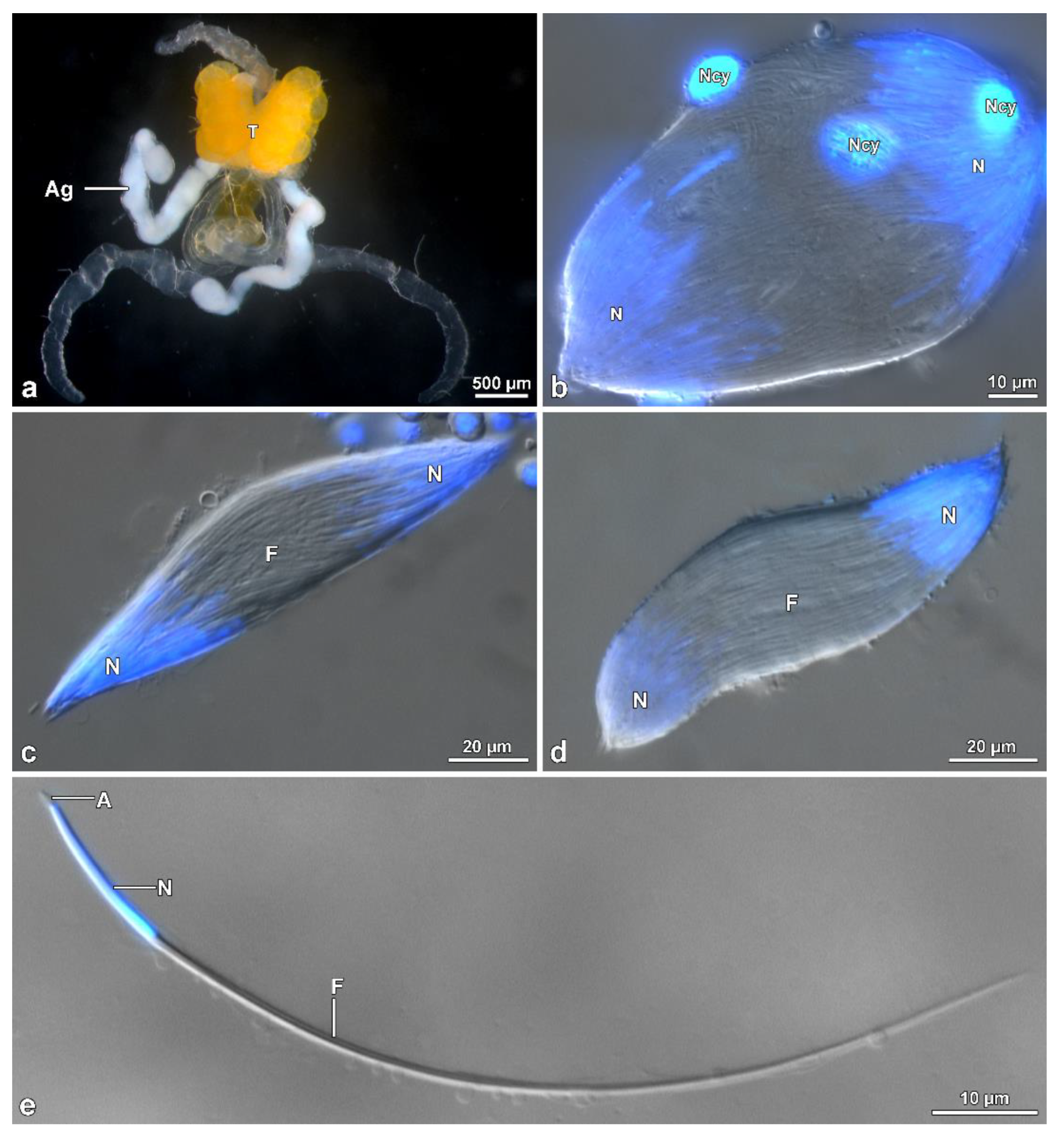
Males of M. brevicauda, O. nobilis and A. velikensis were anesthetized with ether and dissected in 0.1 M phosphate buffer pH 7.2 with 3% of sucrose (PB) to remove the genital system. A drop of sperm, removed from the deferent duct and seminal vesicles, was spread over histological slides and photographed with a Leica DMRB light phase-contrast microscope. The length of spermatozoa was measured using image-J software. For the visualization of sperm nuclei, cells were spread on a histological slide, a drop of 1 ug/mL of the DNA specific dye Hoechst in 0.1 M PB was added, and the sample was finally covered with a glass coverslip. Fluorescence observations of the labelled samples were carried out with a Leica DMRB light microscope equipped with a UV light source, fluorescein, and UV filters and Zeiss AxioCam digital camera with dedicated imaging software.
To observe the entire cysts in Hoshihananomia sp., testes were dissected in PB and transferred to a 2% acetic-orcein solution. After 20 min, the follicles were placed on histological slides with a drop of acetic-orcein solution, dissociated using needles, and covered with coverslips. For testicular histology, testes were fixed in 2.5% glutaraldehyde solution in 0.1 M phosphate buffer, postfixed in 1% osmium tetroxide, dehydrated in alcohol solutions, and embedded in Historesin®. Semithin sections (0.5 µm thick) were stained with Giemsa for 15 min. To measure sperm length, the cells from the vas deferens were spread on histological slides and stained with Giemsa for 15 min. For nuclear size observations and measurements, some samples were stained for 20 min with 0.2 mg/mL DAPI, washed in distilled water, and mounted with 50% sucrose in PB.
2.2. Scanning Electron Microscopy (SEM)
Mature spermatozoa taken from the seminal vesicles and deferent ducts of Mordellistena brevicauda were spread onto coverslips previously treated with poly-l-lysine. The coverslips were placed in 2.5% glutaraldehyde in PB for 30 min at 4 °C and then rinsed several times in PB. Specimens on glass coverslips were dehydrated in a graded series of ethanol and then processed by critical drying method in a Balzer’s CDP 030. The coverslips were sputtered with about 20 nm gold in a Balzer’s MED 010 sputtering device and finally observed in a SEM Phillips XL20 operating at 10 kV electron accelerating voltage.
2.3. Transmission Electron Microscopy (TEM)
Adult males were dissected in PB to isolate the testes and deferent ducts. The material was fixed in 2.5% glutaraldehyde in PB overnight. After careful rinsing, the material was post-fixed in 1% osmium tetroxide for 2 h. After rinsing, the material was dehydrated with ethanol series (50% to 100%), then transferred to propylene oxide, and finally embedded in a mixture of Epon-Araldite resins. Some material was also treated with tannic acid, omitting osmium fixation. Ultrathin sections were obtained with a Reichert Ultracut ultramicrotome, routinely stained with uranyl acetate and lead citrate, and observed with a TEM Philips CM10 operating at 80 kV electron accelerating voltage.
Adult males of the species Hoshihananomia sp. were dissected, and the removed testes were processed following the conventional Transmission Electron Microscopy protocol. Ultrathin sections (~60 nm thick) were obtained with an ultramicrotome (Leica UC6). Then they were contrasted with solutions of 3% uranyl acetate and 0.2% lead citrate and after examined in a Transmission Electron Microscope (Tecnai G2-12—SpiritBiotwin FEI) operating at 120 kV at the Microscopy Center of the Federal University of Minas Gerais (CM-UFMG), Belo Horizonte, Minas Gerais, Brazil.
4. Discussion
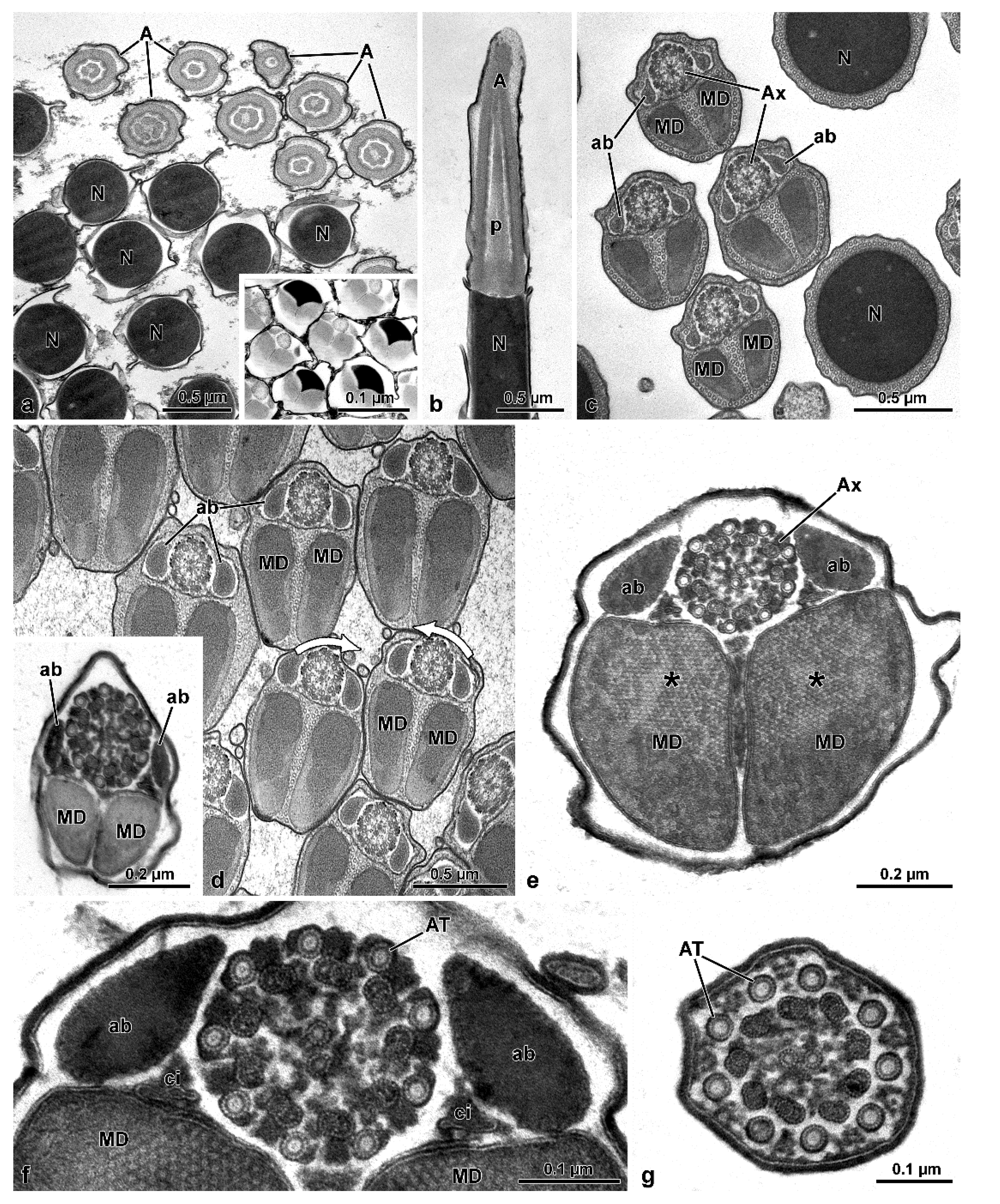
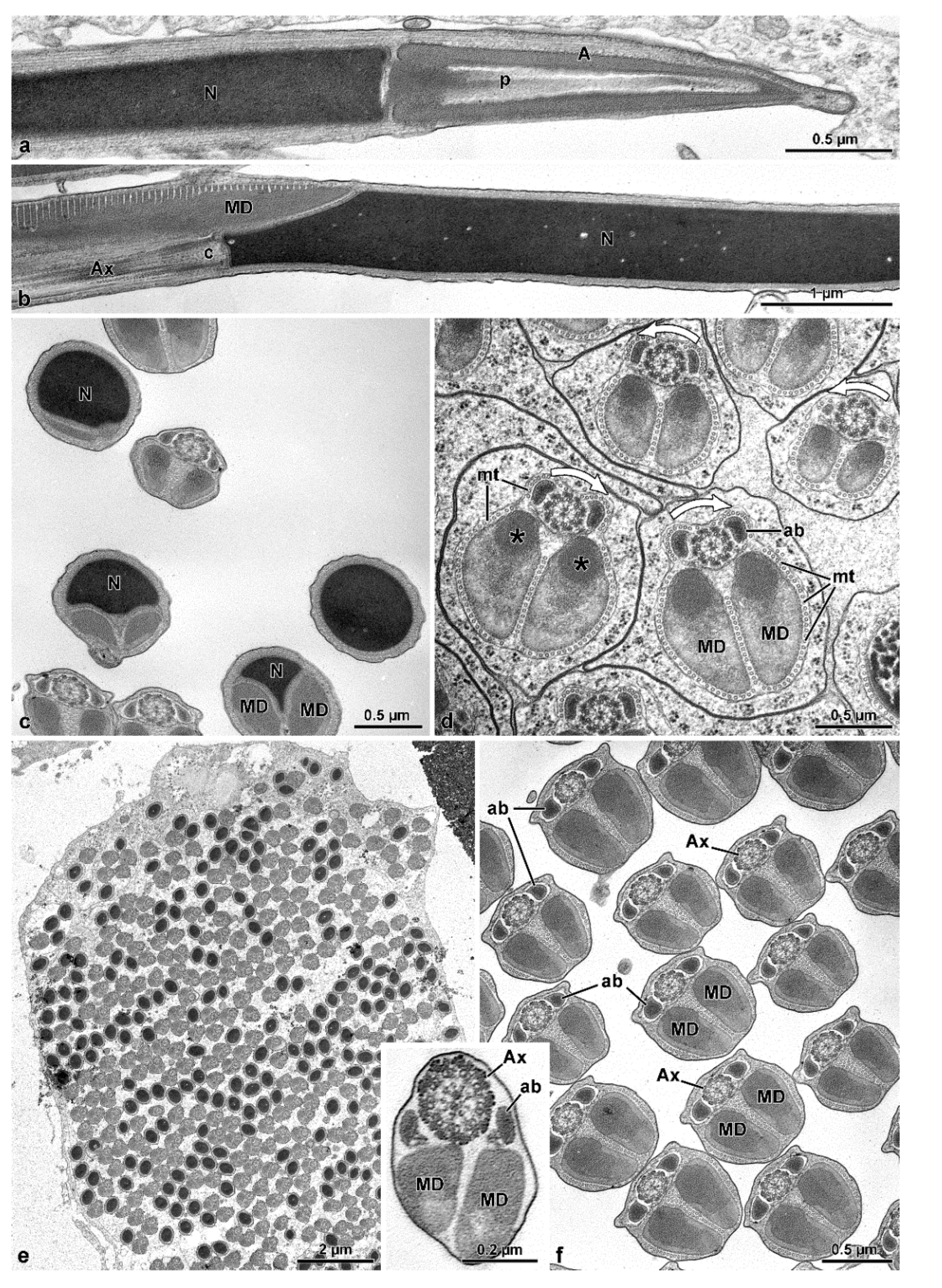
The sperm ultrastructure of the four species studied here not only confirms our previously published data (see [
9]) but, in addition, provides new useful data for a better understanding of the phylogenetic relationships between the various families of Tenebrionoidea. In Oedemeridae, the new family examined here, spermatozoa show the same organization seen in the other advanced members of the superfamily [
7,
8,
9,
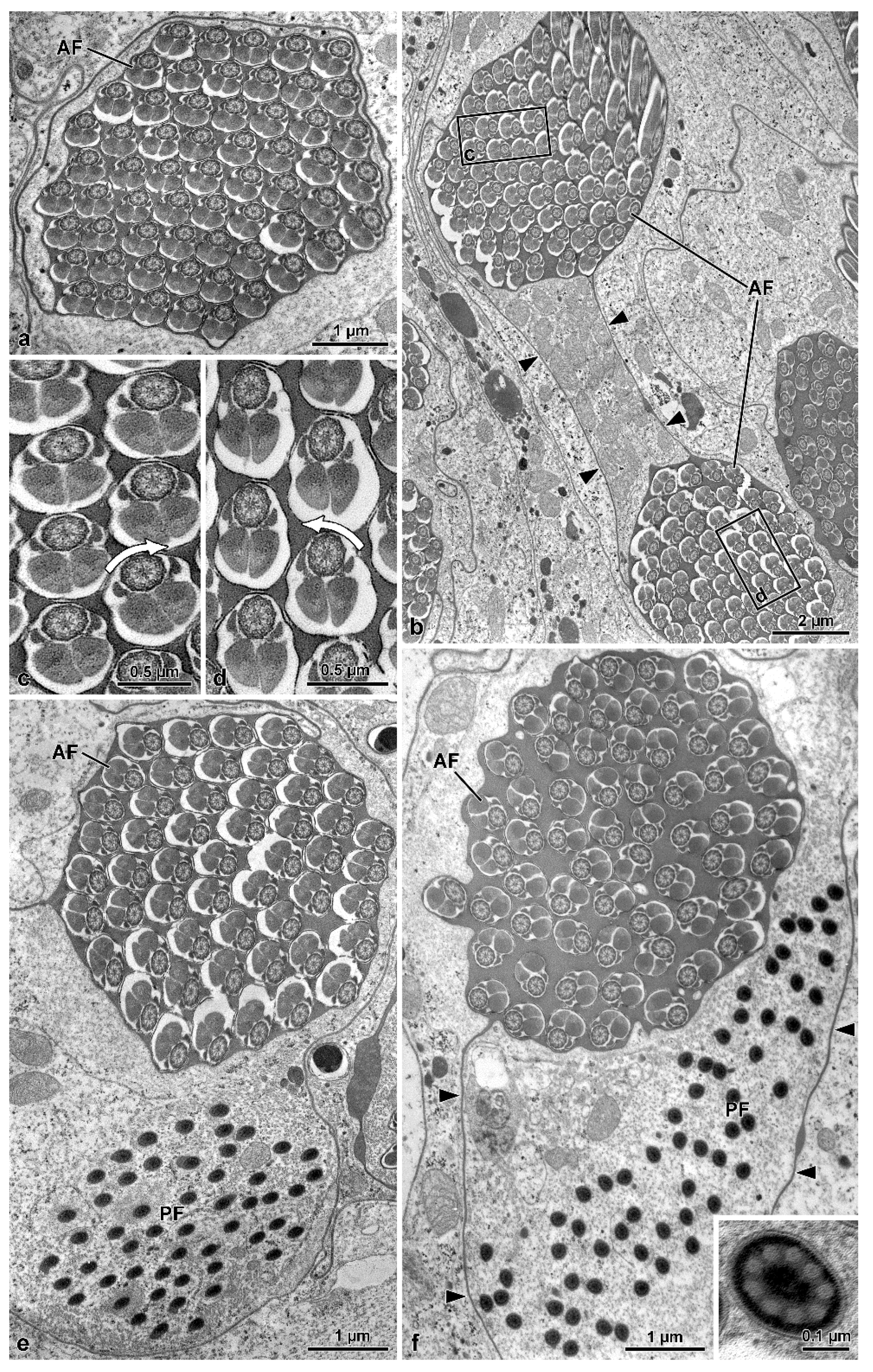
11]. They consist of a short two-layered acrosome, relatively long nucleus, and flagellum; in this latter, an axoneme with 9 + 9 + 2 microtubules, two thick similar mitochondrial derivatives, and two elliptical accessory bodies. However, the features supporting the positioning of this family together with advanced tenebrionoids are the presence of numerous sperm per testicular cyst and the antiparallel arrangement of the spermatozoa within the cyst. These two remarkable features are shared by all tenebrionoid families studied so far, except Mordellidae ([
9], this study) and Ripiphoridae [
9]. The unusual arrangement of sperm within the cysts was initially described in Tenebrionidae by Dias et al. [
8]. These authors demonstrated that this disposition begins in the first stages of spermiogenesis; as the flagella elongate, half of the nuclei migrate to one pole of the cyst and the other half to the opposite pole, ultimately forming two antiparallel sets of sperm per cyst, which are easily observed using DNA stains. Observation by transmission electron microscopy further demonstrated that this arrangement could be inferred from the clockwise and counter-clockwise orientations of axonemal microtubule pairs in cross-sections of cysts (see [
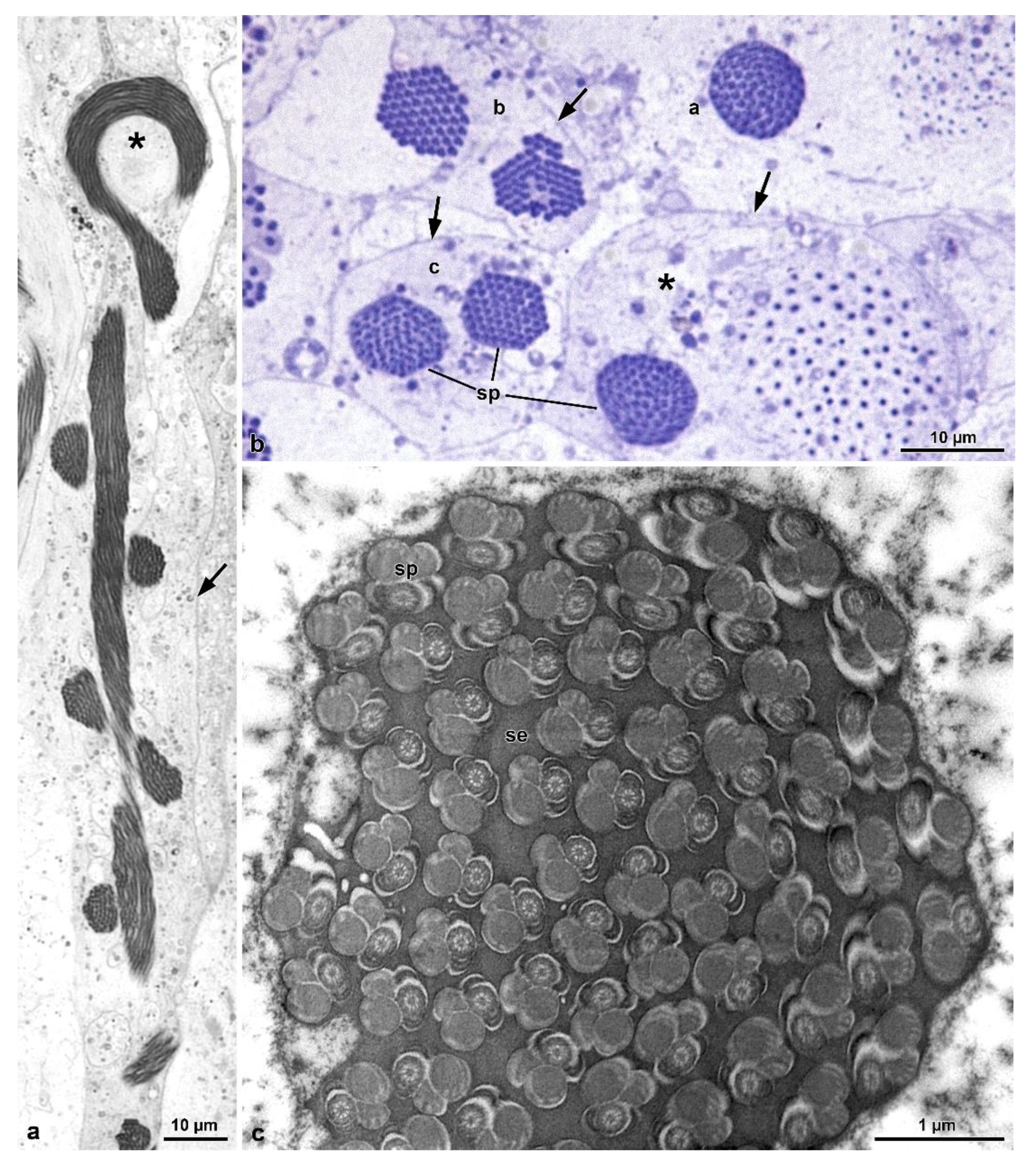
9]) and that this character is common to most Tenebrionoidea. The amount of sperm per cyst is a consequence of the number of division cycles that the initial spermatogonium undergoes in the early stages of spermatogenesis, a number considered constant for the species. In four species of Tenebrionidae [
8], as well as in
A. velikensis and
O. nobilis of this study, up to 512 (=2
9) sperm per cyst were observed, indicative of nine spermatogonial division cycles. On the other hand, in the Tenebrionidae
Lagria villosa (Fabricius, 1781), 1024 (=2
10) sperm were counted per cyst [
15]. Although Zhang et al. [
5] suggested that Ciidae was closely related to the Tenebrionidae, in
Ceracis cornifer (Melli, 1849), 256 sperm per cyst were observed [
11], and the same number was found in four species of Meloidae [
9]. In families of Tenebrionoidea that are considered basal, such as Ripiphoridae and Mordellidae, the number of sperm per cyst is lower, reaching 64 (=2
6) ([
9], this study). Commonly, the works on Oedemeridae, Pythidae, Meloidae, Ciidae, and Tenebrionidae placed these groups in all the main branches of the phylogenetic trees above the more basal branch of Mordellidae and Ripiphoridae [
4,
5,
6,
15,
16]. Thus, it is possible to suppose that the antiparallel disposition of the sperm inside the cyst, and the high number of them per cyst, are characteristics that arose in the common ancestor of all Tenebrionoidea, except Mordellidae and Ripiphoridae. Yet these data indicate that the proposition by Virkki [
17] and Lachaise and Joly [
18] that the occurrence of a relatively low number of sperm per cyst would indicate a derived character state, compared to cysts containing a relatively high number, cannot be applied to all Tenebrionoidea, as recent studies have indicated that Ripiphoridae and Mordellidae would rather form a sister group to all other Tenebrionoidea [
5,
6].
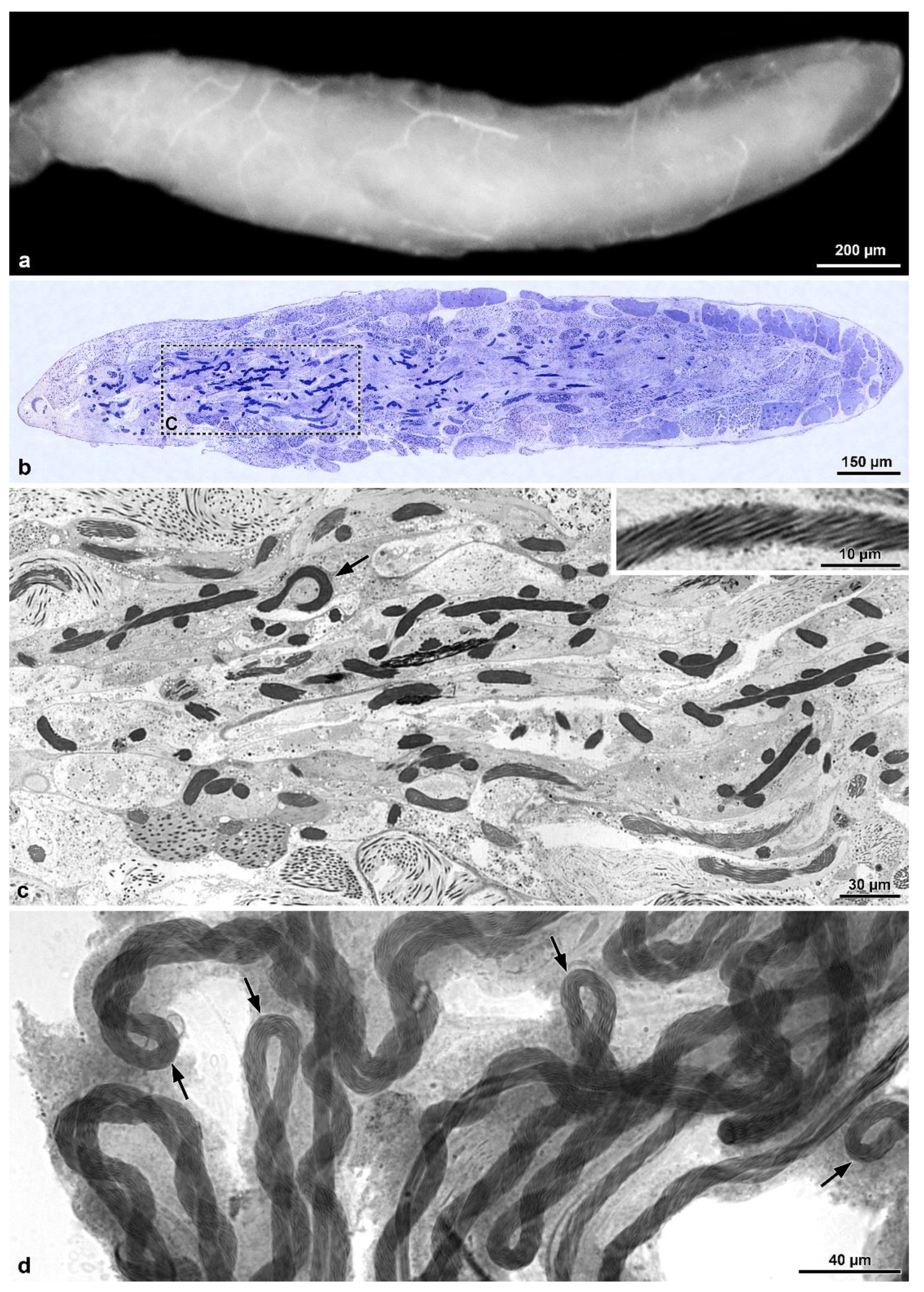
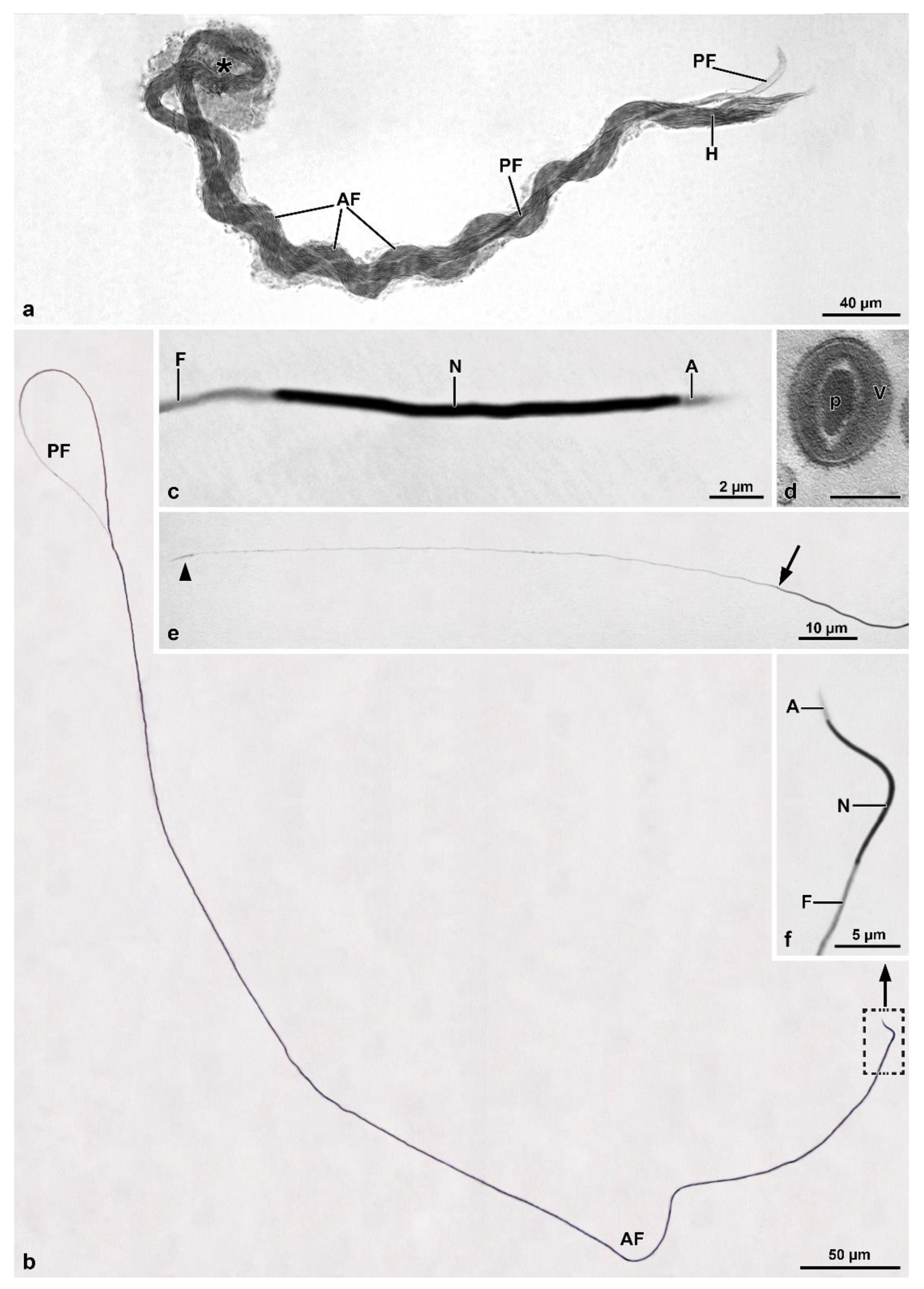
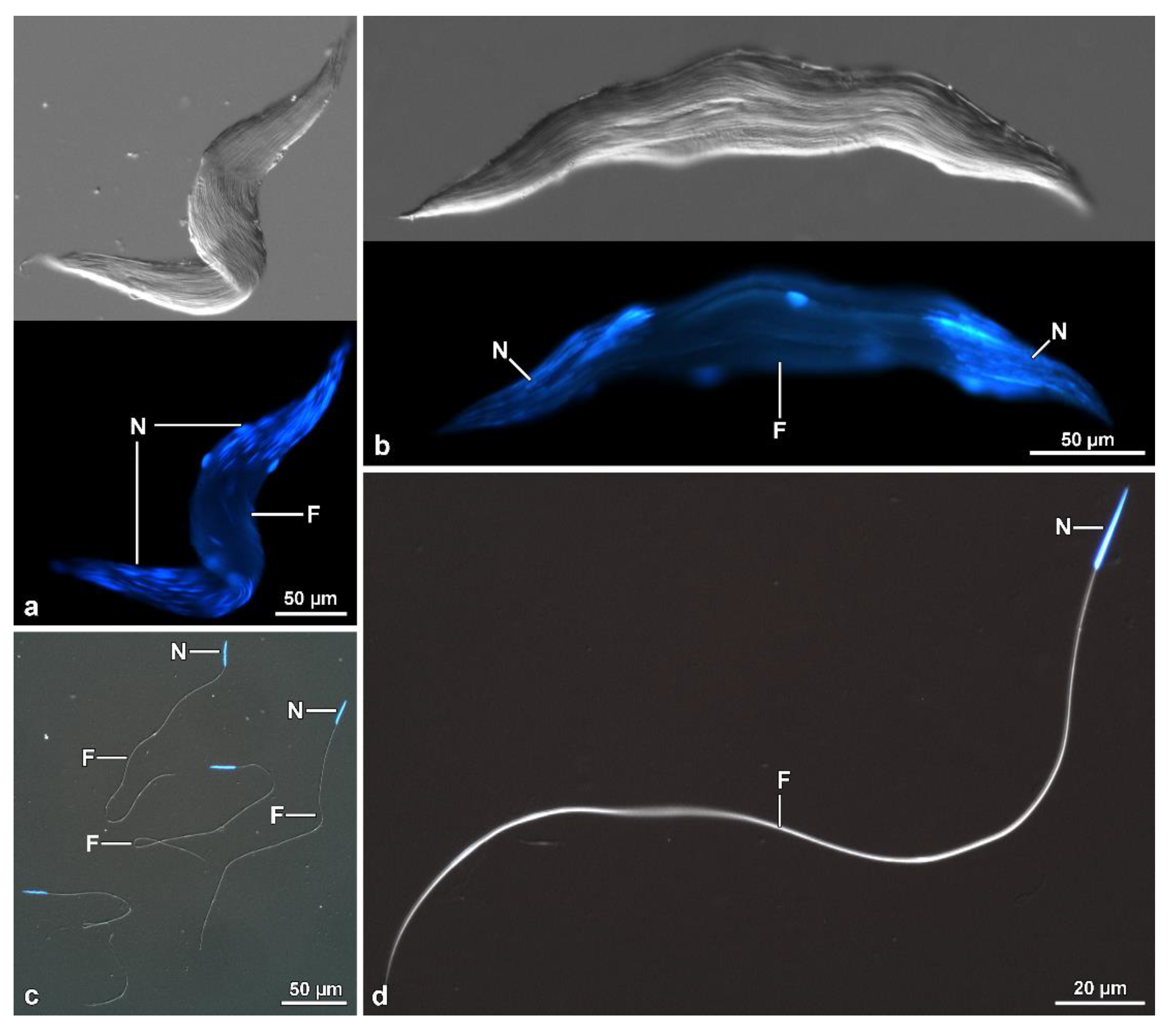
Within the Mordellidae family,
Mordellistena sp. [
9] and
Hoshihananomia sp. have giant sperm, 1230 µm and 1030 µm long, respectively, while those of
M. brevicauda, with about 290 µm, are comparatively very short. Based on molecular data, Batelka et al. [
19] showed, in one of the cladograms, the genera
Mordellistena and
Hoshihananomia in the most basal and most derived branches, respectively. Thus, it is possible to assume that giant sperm may be the synapomorphic condition for Mordellidae. In contrast, short sperm, as observed in
M. brevicauda, is a derived condition within the family. Among all the tenebrionoids studied so far, giant spermatozoa were observed only in Mordellidae and in Ripiphoridae ([
9,
20], this study), which is consistent with the proposal that both families are closely related and form a sister group of all Tenebrionoidea [
5,
6].
A new peculiar finding observed in the two Mordellidae species studied here and possibly in
Mordellistena sp. and the Ripiphoridae
Ptilophorus dufourii [
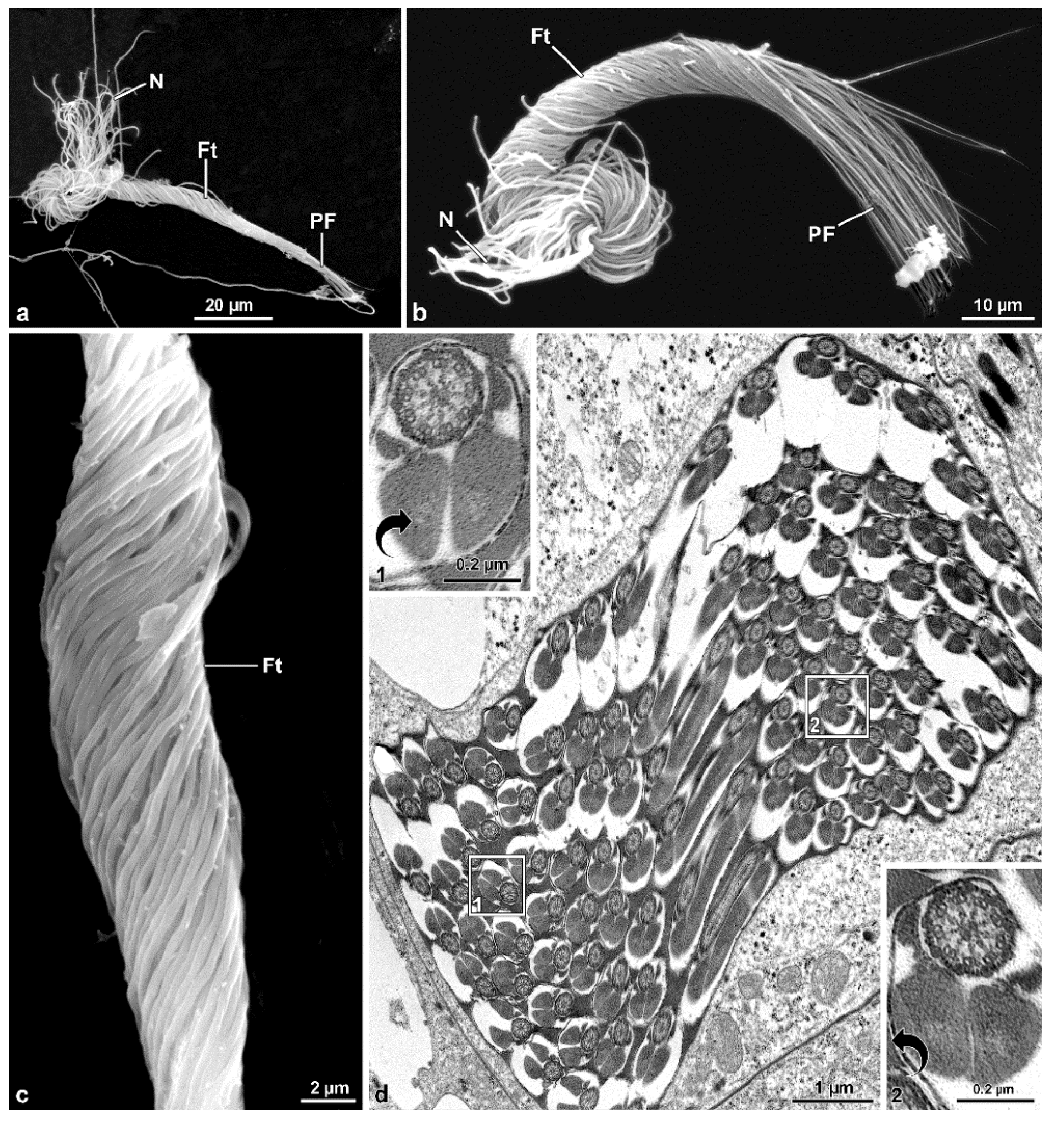
9], deserves to be discussed. The sperm bundles in these species are characterized by bending at half their length forming a loop at this point. In
Hoshihananomia sp., probably because the sperm are very long, it is easy to see that the two folded halves spiral over each other. In
M. brevicauda, this cystic organization is less evident, as the sperm length is only about one-quarter of that in H. sp. However, a twisting of the median region of the sperm bundle is also evident in this species. Thus, the cyst shows in both species a supercoiled organization. The folding of the sperm bundle forming a loop, the spiraling of the two halves, and the twisting of the sperm are probably spermatogenic mechanisms resulting from evolutionary innovations that enabled males with extremely long sperm to produce them in sufficient quantities to be competitive, even in relatively small testes. It was possible to observe that the folding of the sperm bundle within a cyst in the form of a loop begins in the early stages of spermiogenesis, and according to Syed et al. [
13], it is the result of asynchrony between the elongation of spermatid tails and increases in the plasma membrane of surrounding somatic cells. It is essential to point out that this peculiar cystic organization occurs in mordellids either with very long sperm, namely
Hoshihananomia sp. and
Mordellistena sp. [
9], as well as with very short sperm, such as
M. brevicauda. Therefore, this feature must be common to the entire family and it may be likely present in the common ancestor of Mordellidae and Ripiphoridae, sister taxa sharing this cystic organization.
Testicular follicles in H. sp. are elongated and approximately four times longer than those of M. brevicauda, which are ovoid. In tenebrionids the testicular follicles are oval, with the cysts in the different stages distributed in distinct transverse zones: the youngest in the distal zone and the most advanced (about 100 µm long) in the proximal zone to the efferent duct (unpublished data). Such a feature is different from what was observed in Hoshihananomia sp., in which the long cysts are distributed along almost the entire central region of the follicles. These differences seem to validate an existing correlation between sperm and testis lengths and the organization of cysts within follicles.
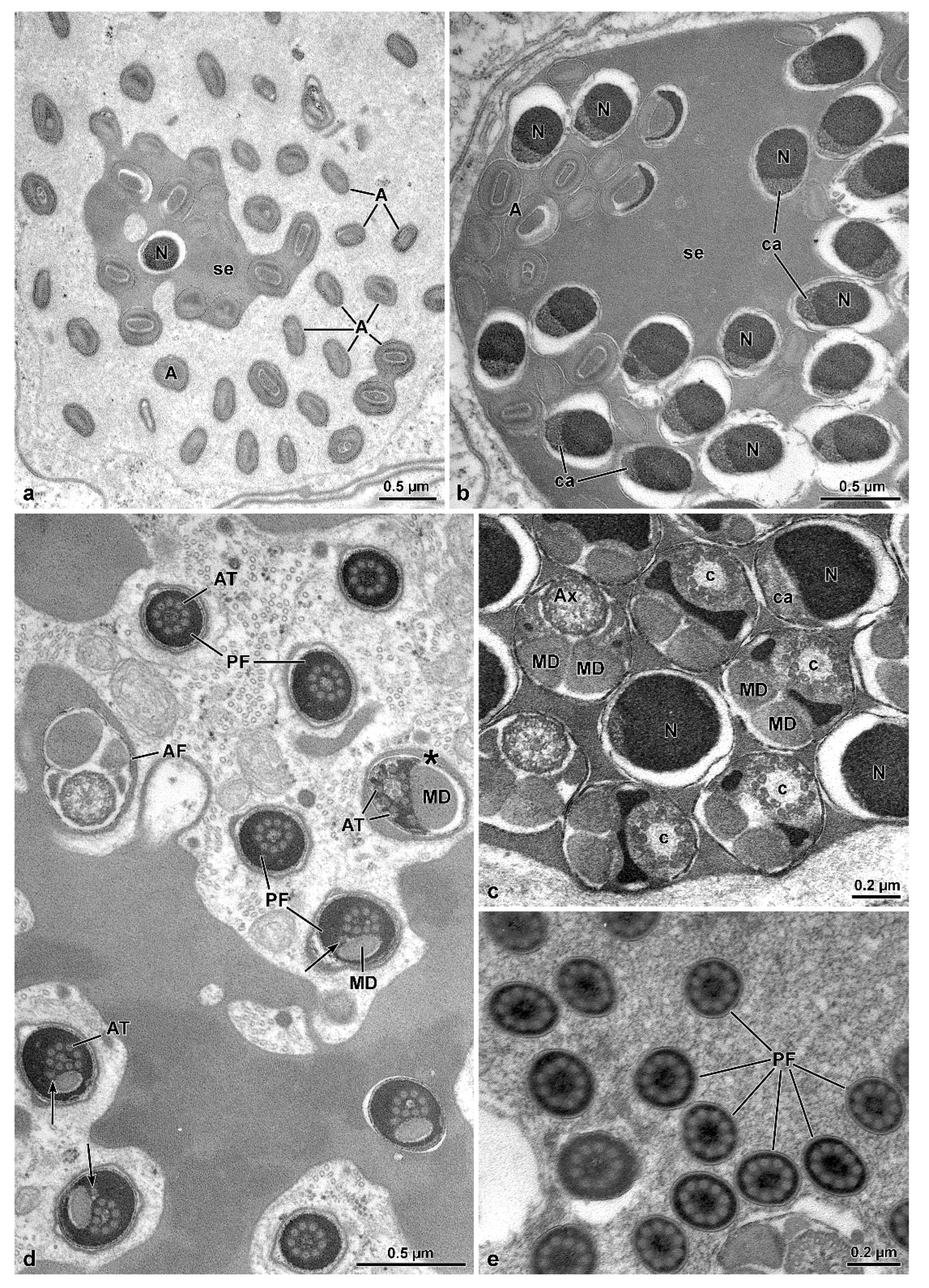
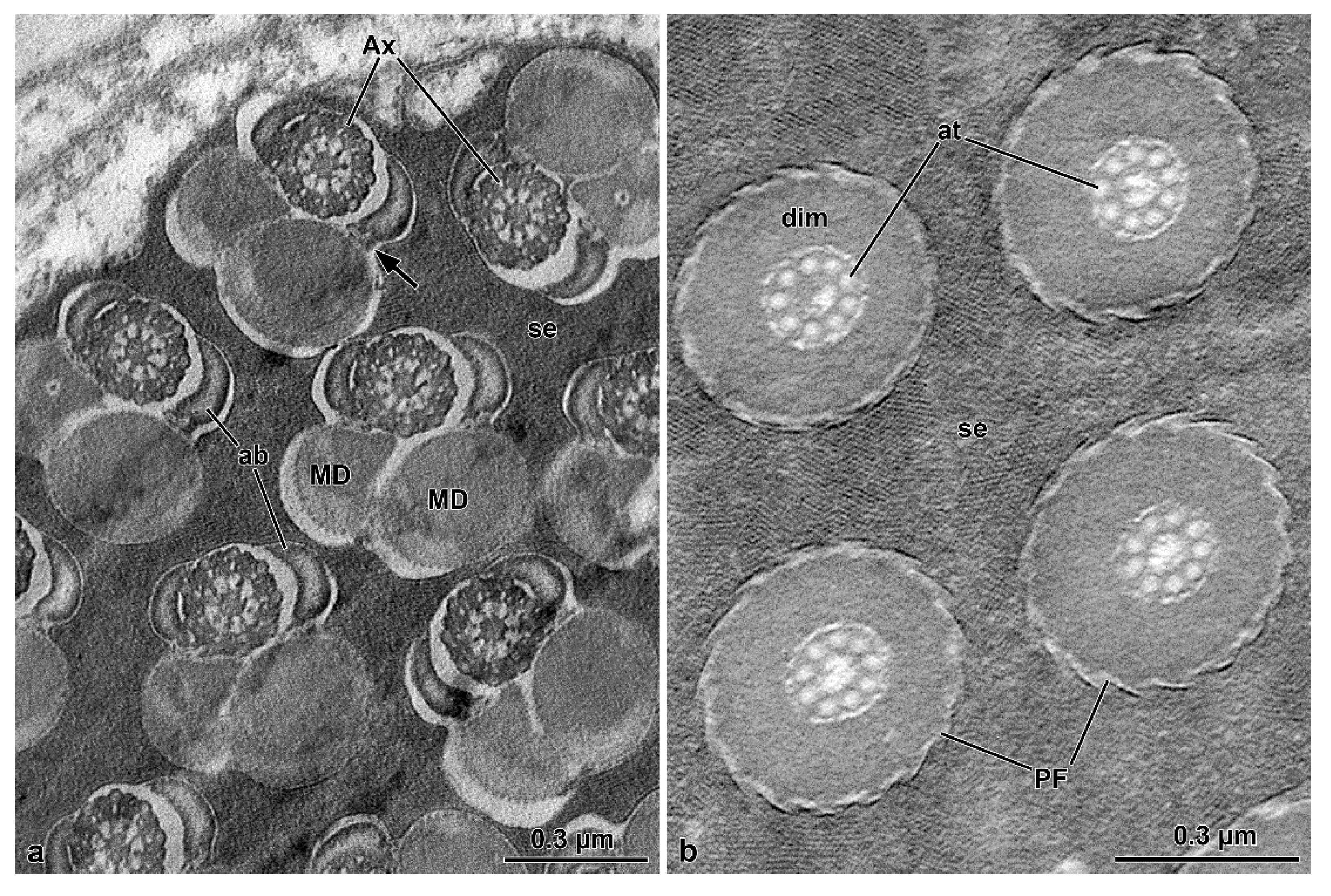
Mordellidae sperm ([
9], this study) are characterized by a long posterior flagellar tip. In many insects this region is affected by axoneme disorganization with the microtubule doublets becoming grossly irregular structures and lacking their dynein arms, as described in the orthopteran
Gryllotalpa gryllotalpa [
10,
21], the zorapteran
Zorotypus caudelli [
10], and in the dipterans
Drosophila melanogaster and
Bactrocera oleae [
10]. In the mordellids, however, axonemal degeneration occurs by the loss of the central 9 + 2 microtubule complex, while accessory tubules persist throughout this entire region. Due to the lack of microtubule pairs and consequently dynein arms, this region is stiff and immotile. Still in Mordellidae ([
9], this study), unlike the other Tenebrionoidea, there is a compact material among the sperm in the cysts; in the sperm, the centriole adjunct flanks the entire nucleus, and the tail posterior end is long, formed only by axonemal accessory tubules embedded in dense intracellular material. The absence of these features in ripiphorids, and the phylogenetic distance of
Mordellistena and
Hoshihananomia [
19], suggest that they may be unique traits (autopomorphies) of Mordellidae. Furthermore, these traits may constitute good phylogenetic signals to understand the relationships between the basal families of Tenebrionoidea. Alternatively, as suggested by Hunt et al. [
15], the clade at the base of Tenebrionoidea could be Ripiphorinae and Mordellinae together with two subfamilies of Lymexyloidea.
The sperm flagellar structure of insects is also characterized by the size and shape of the mitochondrial derivatives and the accessory bodies [
10]. All the species have their two mitochondrial derivatives with a similar shape. However, these structures are more developed and elliptical in
O. nobilis and
A. velikensis while in the two species of Mordellidae they are smaller and oval-shaped. The two accessory bodies, flanking the axoneme, are of primary importance to verify the relationship between the families of Tenebrionoidea. It has been already pointed out that the oval or elliptical shape of these structures in cross-section, is typical of various species of Tenebrionidae such as
Tenebrio molitor or
Tribolium castaneum [
7,
9] and also of
A. velikensis here studied. This shape, with few variations, is the commonest feature among the whole superfamily, and it was also found in Meloidae [
9,
20].
O. nobilis, a member of the new family here studied has elliptical accessory bodies with a pointed apical side. On the contrary Mordellidae species have smaller accessory bodies with an almost triangular shape ([
9], this study).
As quoted in the Introduction, Levkaničová [
3] and Bocak et al. [
4] considered Ripiphoridae, Mordellidae, and Meloidae closely related. However, the latter authors considered the clade with the three families the most derived, while Levkaničová considered it the most basal. From the testicular and sperm morphology ([
9,
20], this study) it is possible to assume that Mordellidae and Ripiphoridae, but not Meloidae, share a recent common ancestor, a condition also proposed by Zhang et al. [
5] and McKenna et al. [
6] from extensive gene sampling. They further suggested that the clade with these two families is at the base of the superfamily. A position that will possibly be supported by sperm and testicular morphology, but this type of data should be extended to other families and also to groups closely related to Tenebrionoidea, such as Lymexyloidea.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











