
High-Quality Queens Produce High-Quality Offspring Queens




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and F1 Generation Queens Rearing
2.2. Gene Expression Analysis
2.3. F2 Generation Queens Rearing
2.4. Physiological Indexes of F2 Queens
2.5. Statistical Analysis
3. Results
3.1. The Physiological Indexes of Reared F1 Queens
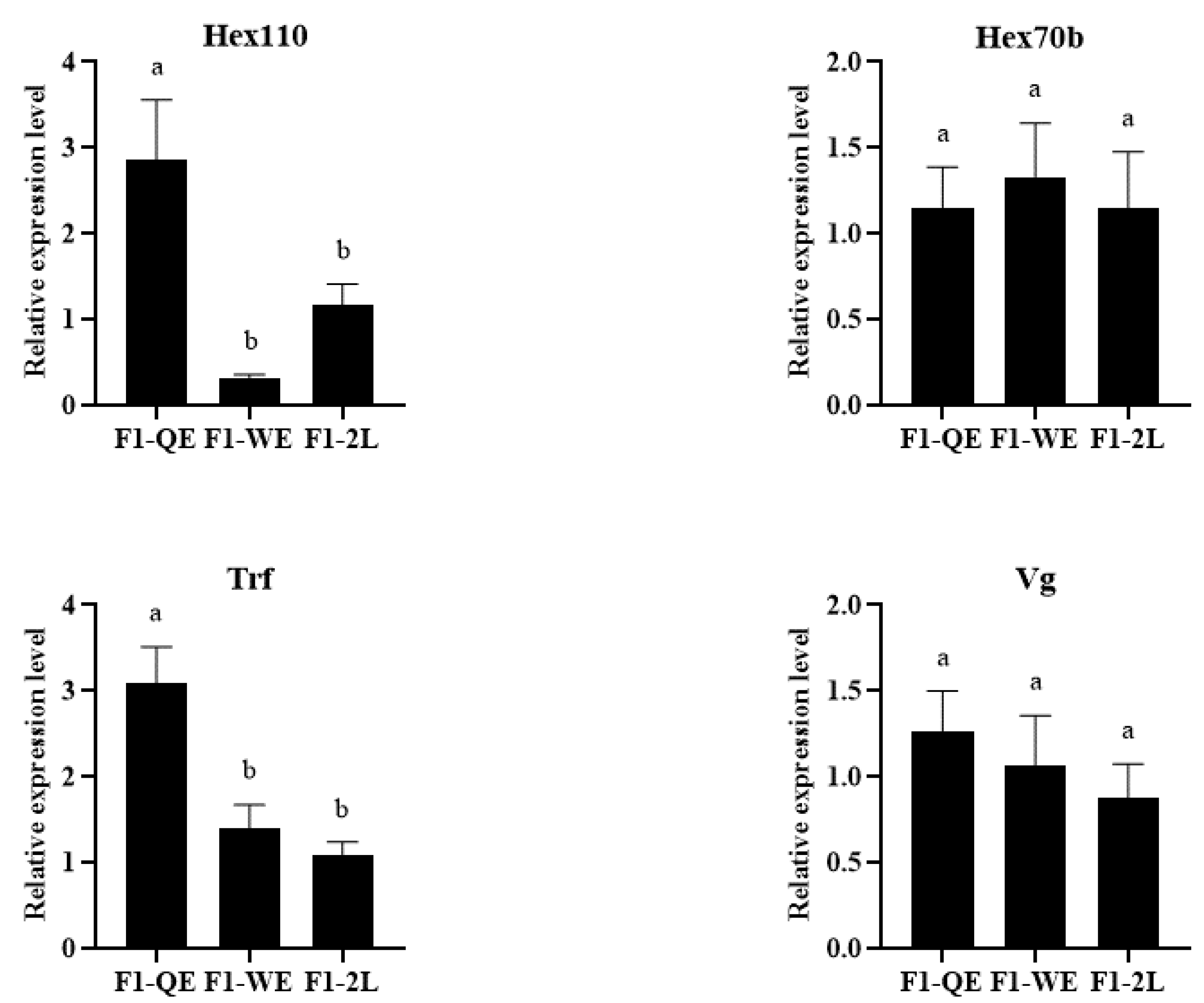
3.2. The Expression of Development-Related Genes of F1 Queens
3.3. The Weight and Ovarioles Number of F2 Queens
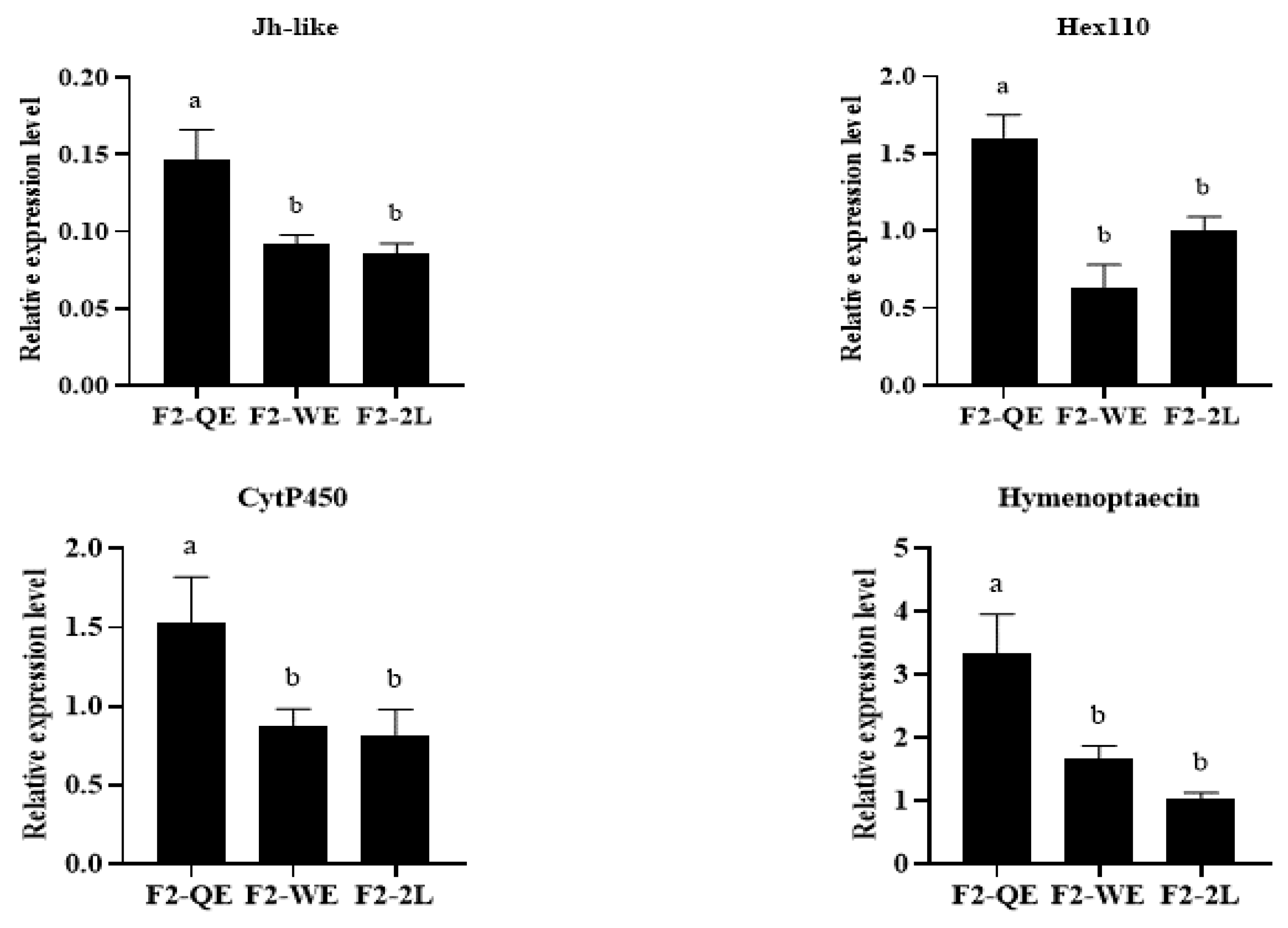
3.4. The Relative Expression of the Development-Related Genes of F2 Queens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, UK, 1987. [Google Scholar]
- Adgaba, N.; Al-Ghamdi, A.; Tadesse, Y.; Tadesse, Y.; Sarhan, R.; Single, A.; Mohammed, S.E.; Khan, K.A. The responses of Apis mellifera jemenitica to different artificial queen rearing techniques. Saudi. J. Biol. Sci. 2018, 26, 1649–1654. [Google Scholar] [CrossRef] [PubMed]
- Stankus, T. A review and bibliography of the literature of honey bee colony collapse disorder: A poorly understood epidemic that clearly threatens the successful pollination of billions of dollars of crops in America. J. Agric. Food Inf. 2008, 9, 115–143. [Google Scholar] [CrossRef]
- Kulhanek, K.; Steinhauer, N.; Rennich, K.; Caron, D.M.; Sagili, R.R.; Pettis, J.S.; Ellis, J.D.; Wilson, M.E.; Wilkes, J.T.; Tarpy, D.R.; et al. A national survey of managed honey bee 2015–2016 annual colony losses in the USA. J. Apic. Res. 2017, 56, 328–340. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Liu, Y.B.; Barron, A.B.; Zeng, Z.J. Effects of commercial queen rearing methods on queen fecundity and genome methylation. Apidologie 2020, 52, 282–291. [Google Scholar] [CrossRef]
- Tarpy, D.R.; Keller, J.J.; Caren, J.R.; Delaney, D.A. Assessing the mating ‘health’ of commercial honey bee queens. J. Econ. Entomol. 2012, 105, 20–25. [Google Scholar] [CrossRef]
- Amiri, E.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Queen quality and the impact of honey bee diseases on queen health: Potential for interactions between two major threats to colony health. Insects 2017, 8, 48–66. [Google Scholar] [CrossRef]
- Tarpy, D.R. Genetic diversity within honeybee colonies prevents severe infections and promotes colony growth. Proc. R. Soc. B Sci. 2003, 270, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Woyke, J. Correlations between the age at which honeybee brood was grafted, characteristics of the resultant queens, and results of insemination. J. Apic. Res. 1971, 10, 45–55. [Google Scholar] [CrossRef]
- Wang, W.X.; Tian, L.Q.; Huang, Q.; Wu, X.B.; Zeng, Z.J. Effects of 10-Hydroxy-2-decenoic acid on the development of honey bee (Apis mellifera) larvae. J. Apic. Res. 2014, 53, 171–176. [Google Scholar] [CrossRef]
- Kahya, Y.; Gençer, H.V.; Woyke, J. Weight at emergence of honey bee (Apis Mellifera Caucasica) Queens and its effect on live weights at the pre and post mating periods. J. Apic. Res. 2008, 47, 118–125. [Google Scholar] [CrossRef]
- Delaney, D.A.; Keller, J.J.; Caren, J.R.; Tarpy, D.R. The physical, insemination, and reproductive quality of honey bee queens (Apis Mellifera L.). Apidologie 2011, 42, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hatjina, F.; Bieńkowska, M.; Charistos, L.; Chlebo, R.; Costa, C.; Dražić, M.M.; Filipi, J.; Gregorc, A.; Ivanova, E.N.; Kezić, N.; et al. A review of methods used in some European countries for assessing the quality of honey bee queens through their physical characters and the performance of their colonies. J. Apic. Res. 2014, 53, 337–363. [Google Scholar] [CrossRef] [Green Version]
- Niño, E.L.; Tarpy, D.R.; Grozinger, C.M. Differential effects of insemination volume and substance on reproductive changes in honey bee queens (Apis mellifera L.). Insect Mol. Biol. 2013, 22, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Rangel, J.; Keller, J.J.; Tarpy, D.R. The effects of honey bee (Apis mellifera L.) queen reproductive potential on colony growth. Insectes Sociaux 2013, 60, 65–73. [Google Scholar] [CrossRef]
- Den Boer, S.P.A.; Boomsma, J.J.; Baer, B. Honey bee males and queens use glandular secretions to enhance sperm viability before and after storage. J. Insect Physiol. 2009, 55, 538–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.R.; Nunes, F.M.F.; Simões, Z.L.P.; Bitondi, M.M.G. A honeybee storage protein gene, hex70a, expressed in developing gonads and nutritionally regulated in adult fat body. J. Insect Physiol. 2008, 54, 867–877. [Google Scholar] [CrossRef]
- Martins, J.R.; Nunes, F.M.; Cristino, A.S.; Bitondi, M.M.G. The four hexamerin genes in the honey bee: Structure, molecular evolution and function deduced from expression patterns in queens, workers and drones. BMC Mol. Biol. 2010, 11, 23–43. [Google Scholar] [CrossRef] [Green Version]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran, L.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef] [Green Version]
- Koywiwattrakul, P.; Sittipraneed, S. Expression of vitellogenin and transferrin in activated ovaries of worker honey bees, Apis mellifera. Biochem. Genet. Biol. 2009, 47, 19–26. [Google Scholar] [CrossRef]
- Amdam, G.V.; Simões, Z.L.P.; Hagen, A.; Norberg, K.; Schrøder, K.; Mikkelsen, Ø.; Kirkwood, T.B.; Omholt, S.W. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 2004, 39, 767–773. [Google Scholar] [CrossRef]
- Kuterbach, D.A.; Walcott, B.; Reeder, R.J.; Frankel, R.B. Iron-containing cells in the honey bee (Apis mellifera). Science 1982, 218, 695–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharski, R.; Maleszka, R. Transcriptional profiling reveals multifunctional roles for transferrin in the honeybee, Apis mellifera. J. Insect Sci. 2003, 3, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddiford, L.M. Cellular and molecular actions of juvenile hormone I. General considerations and premetamorphic actions. In Advances in Insect Physiology; Academic Press: Cambridge, MA, USA, 1994; pp. 213–274. [Google Scholar]
- Robinson, G.E. Regulation of honey bee age polyethism by Juvenile hormone. Behav. Ecol. Sociobiol. 1987, 20, 329–338. [Google Scholar] [CrossRef]
- Huang, Z.Y.; Robynson, G.E.; Tobe, S.S.; Yagi, K.J.; Strambi, C.; Strambi, A.; Stay, B. Hormonal regulation of behavioral development in the honey bee is based on changes in the rate of juvenile hormone biosynthesis. J. Insect Physiol. 1991, 37, 733–741. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Riviere, L.; Damme, J.; Elicone, C.; Fleming, M.; Jacobs, F.; Tempst, P. Isolation and characterization of abaecin, a major antibacterial response peptide in the honeybee (Apis mellifera). FEBS J. 2010, 187, 381–386. [Google Scholar] [CrossRef]
- Wei, H.; He, X.J.; Liao, C.H.; Wu, X.B.; Jiang, W.J.; Zhang, B.; Zhou, L.B.; Zhang, L.Z.; Barron, A.B.; Zeng, Z.J. A Maternal Effect on Queen Production in Honeybees. Curr. Biol. 2019, 29, 2208–2213.e3. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Vaeck, M.; Tempst, P. Apidaecins: Antibacterial peptides from honeybees. EMBO J. 1989, 8, 2387–2391. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Tempst, P. Functional and chemical characterization of Hymenoptaecin, an antibacterial polypeptide that is infection-inducible in the honeybee (Apis mellifera). J. Biol. Chem. 1993, 268, 7044–7054. [Google Scholar] [CrossRef]
- Qiu, Y.; Tittiger, C.; Wicker-Thomas, C.; Goff, G.L.; Young, S.; Wajnberg, E.; Fricaux, T.; Taquet, N.; Blomquist, T.G.; Feyereisen, R. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14858–14863. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D.; Wheeler, D.E. Differential gene expression between developing queens and workers in the honey bee, Apis mellifera. Proc. Natl. Acad. Sci. USA 1999, 96, 5575–5580. [Google Scholar] [CrossRef] [Green Version]
- Shell, W.A.; Rehan, S.M. Behavioral and genetic mechanisms of social evolution: Insights from incipiently and facultatively social bees. Apidologie 2018, 49, 13–30. [Google Scholar] [CrossRef]
- He, X.J.; Wei, H.; Jiang, W.J.; Liu, Y.B.; Zeng, Z.J. Honeybee (Apis mellifera) maternal effect causes alternation of DNA methylation regulating queen development. Sociobiology 2021, 68, e5935. [Google Scholar] [CrossRef]
- Bernardo, J. Maternal Effects in Animal Ecology. Integr. Comp. Biol. 1996, 36, 83–105. [Google Scholar] [CrossRef] [Green Version]
- Roach, D.A.; Wulff, R.D. Maternal Effects in Plants. Annu. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Schwabl, H.; Groothuis, G.G.T. Maternal Effects on Behavior. Encycl. Anim. Behav. 2010, 128, 399–411. [Google Scholar]
- Cunningham, E.; Russell, A.F. Egg investment is influenced by male attractiveness in mallards. Nature 2000, 404, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Passera, L. The laying of biased eggs by the ant Pheidole pallidula (Nyl,)(Hymenoptera, Formicidae). Insectes Sociaux 1980, 27, 79–95. [Google Scholar] [CrossRef]
- Schwander, T.; Humbert, J.Y.; Brent, C.S.; Cahan, S.H.; Chapuis, L.; Renai, E.; Keller, L. Maternal effect on female caste determination in a social insect. Curr. Biol. Soc. 2008, 18, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiri, E.; Le, K.; Melendez, C.V.; Strand, M.K.; Trapy, D.R.; Rueppell, O. Egg-size plasticity in Apis mellifera: Honey bee queens alter egg size in response to both genetic and environmental factors. J. Evol. Biol. 2020, 33, 534–543. [Google Scholar] [CrossRef]
- Gan, H.Y.; Tian, L.Q.; Yan, W.Y. Paraffin section method of queen ovary. J. Bee 2012, 32, 9. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wilson, E.O. The Insect Societies; Harvard University Press: Cambridge, UK, 2017. [Google Scholar]
- Watson, L.R. Controlled mating in honeybees. Q. Rev. Biol. 1928, 3, 377–390. [Google Scholar] [CrossRef]
- Winston, M.L. Events following queen removal in colonies of Africanized honeybees in south America. Insectes Sociaux 1979, 26, 373–381. [Google Scholar] [CrossRef]
- Shi, Y.Y.; Huang, Z.Y.; Zeng, Z.J.; Wang, Z.L.; Wu, X.B.; Yan, W.Y. Diet and cell size both affect queen-worker differentiation through DNA methylation in honey bees (Apis mellifera, Apidae). PLoS ONE 2011, 6, e18808. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, L.; Zhang, W.; Cui, X.; Wang, H.; Xu, B. Comparison of the nutrient composition of royal jelly and worker jelly of honey bees (Apis mellifera). Apidologie 2016, 47, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Sagili, R.R.; Pankiw, T. Effect of brood pheromone modulated brood rearing behaviors on honey bee (Apis mellifera L.) colony growth. J. Insect Behav. 2009, 22, 339–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocks, C.; Ju, H.C.; Nehme, N.; Ulvila, J.; Pearson, A.M.; Meiter, M.; Strom, C.; Conto, S.L.; Hetru, C.; Stuart, L.M.; et al. Eater, a Transmembrane Protein Mediating Phagocytosis of Bacterial Pathogens in Drosophila. Cell 2005, 123, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Grozinger, C.M.; Fan, Y.; Hoover, S.E.R.; Winston, M.L. Genome-wide analysis reveals differences in brain gene expression patterns associated with caste and reproductive status in honey bees (Apis mellifera). Mol. Ecol. 2010, 16, 4837–4848. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward Primer 5′-3′ | Reverse Primer 5′-3′ |
|---|---|---|
| GAPDH | GCTGGTTTCATCGATGGTTT | ACGATTTCGACCACCGTAAC |
| -actin | TCCTGCTATGTATGTCGC | AGTTGCCATTTCCTGTTC |
| Hex110 | AACGTGCCAGGCGCAGTTGT | TTCACCAGCATGGAGGTTCTGGA |
| Hex70b | GAGGACGGTAGCGAGTCCTT | ATGTTGCGGCCCAATACAGG |
| Trf | AGCGGCATACTCCAGGGAC | CGTTGAGCCTGATCCATACGA |
| Jh-like | CACTGGCACCAGAGCCTGTC | GATTCCCATTGAACGAGCGA |
| Abeacin | TCTTCGCACTACTCGCCACG | TCAGGGACCATTCAATCCGA |
| CytP450 | CAAAATGGTGTTCTCCTTACCG | ATGGCAACCCATCACTGC |
| Hymenoptaecin | TCAAGCGGAATTGGAACCTG | CGAATCCTGCATGCTGTCGA |
| Vg | CGTGTTCCAGAGGACGTTGA | ACGCTCCTCAGGCTCAACTC |
| Groups | Weight of Newly Emerged Queens | Thorax Width |
|---|---|---|
| F1-QE | 267.50 ± 6.33 a | 4.94 ± 0.07 a |
| F1-WE | 235.40 ± 7.75 b | 4.88 ± 0.06 a |
| F1-2L | 177.28 ± 12.41 c | 4.66 ± 0.067 b |
| Groups | Weight of Newly Emerged Queens | Number of Ovarioles |
|---|---|---|
| F2-QE | 277.30 ± 6.33 a | 116.71 ± 4.70 a |
| F2-WE | 257.53 ± 6.16 b | 107.71 ± 3.60 ab |
| F2-2L | 262.88 ± 1.98 b | 103.43 ± 2.71 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Shi, X.; He, X.; Zeng, Z.; Yan, W.; Wu, X. High-Quality Queens Produce High-Quality Offspring Queens. Insects 2022, 13, 486. https://doi.org/10.3390/insects13050486
Yu L, Shi X, He X, Zeng Z, Yan W, Wu X. High-Quality Queens Produce High-Quality Offspring Queens. Insects. 2022; 13(5):486. https://doi.org/10.3390/insects13050486
Chicago/Turabian StyleYu, Longtao, Xinxin Shi, Xujiang He, Zhijiang Zeng, Weiyu Yan, and Xiaobo Wu. 2022. "High-Quality Queens Produce High-Quality Offspring Queens" Insects 13, no. 5: 486. https://doi.org/10.3390/insects13050486
APA StyleYu, L., Shi, X., He, X., Zeng, Z., Yan, W., & Wu, X. (2022). High-Quality Queens Produce High-Quality Offspring Queens. Insects, 13(5), 486. https://doi.org/10.3390/insects13050486