
Effects of Habitat Fragmentation on the Population Structure and Genetic Diversity of Erythroneurini in the Typical Karst Rocky Ecosystem, Southwest China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
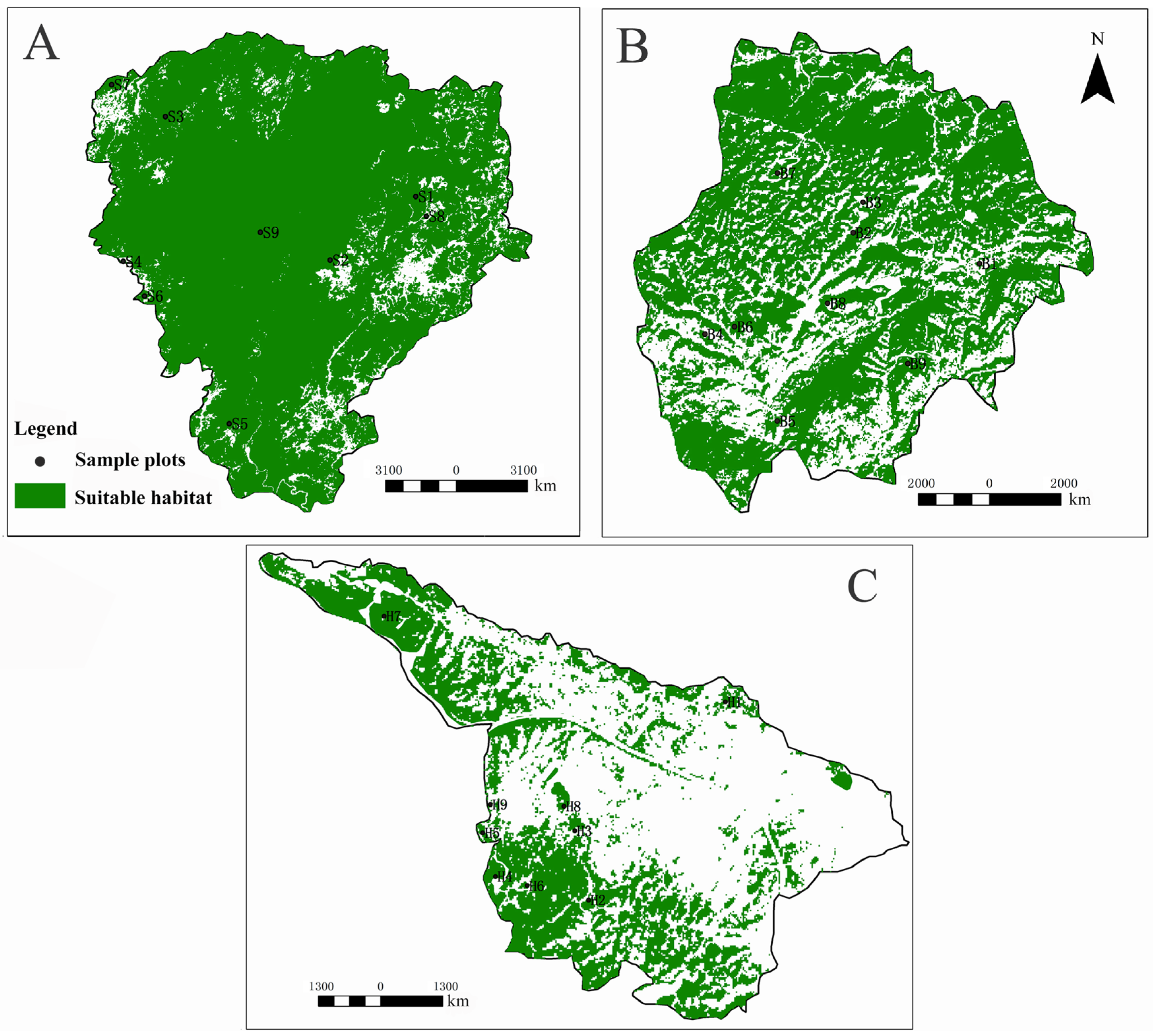
2.1. Studied Areas
2.2. Sample Plot Establishment
2.3. Determination of the Degree of Habitat Fragmentation
2.4. Species Diversity Analysis
2.5. Genetic Diversity Analysis
2.6. Correlation Analysis
3. Results and Analysis
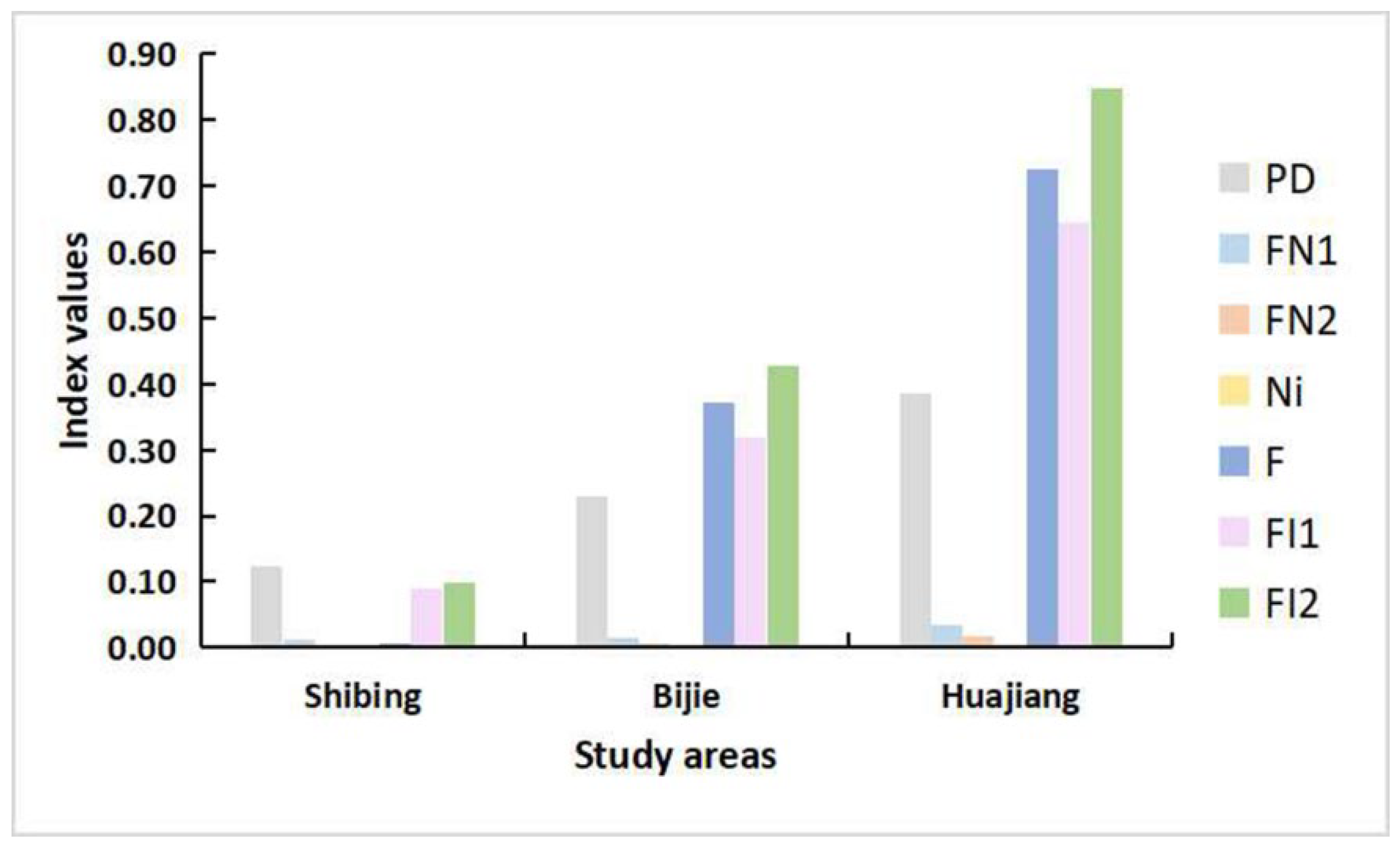
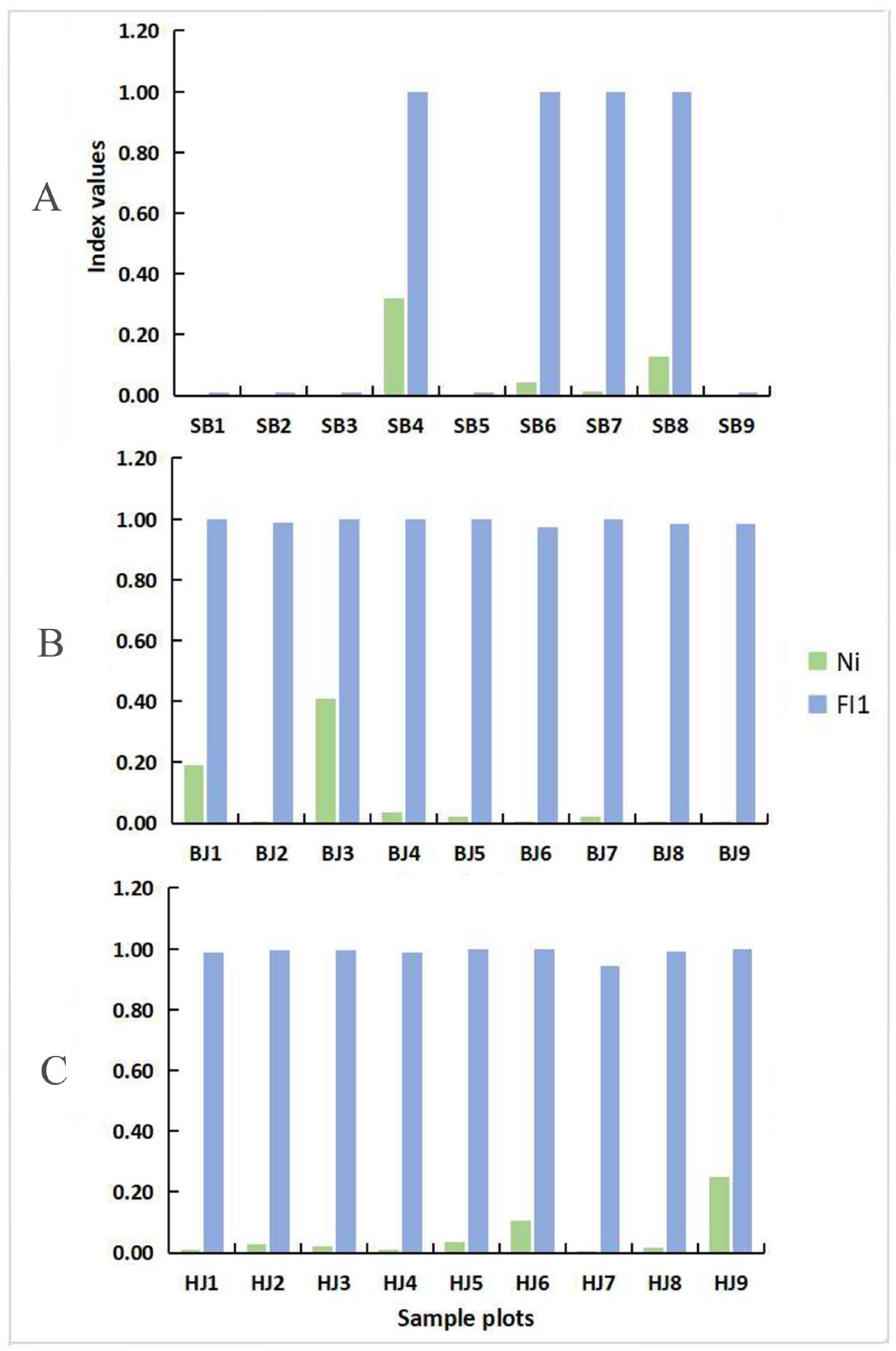
3.1. Determination of the Degree of Habitat Fragmentation
3.1.1. Habitat Spatial Distribution of Erythroneurine Leafhoppers in the Study Area
3.1.2. Habitat Fragmentation of Three Different Study Areas
3.1.3. Habitat Fragmentation of the Sample Plots
3.2. Effects of Habitat Fragmentation on the Species Diversity of Erythroneurine Leafhoppers
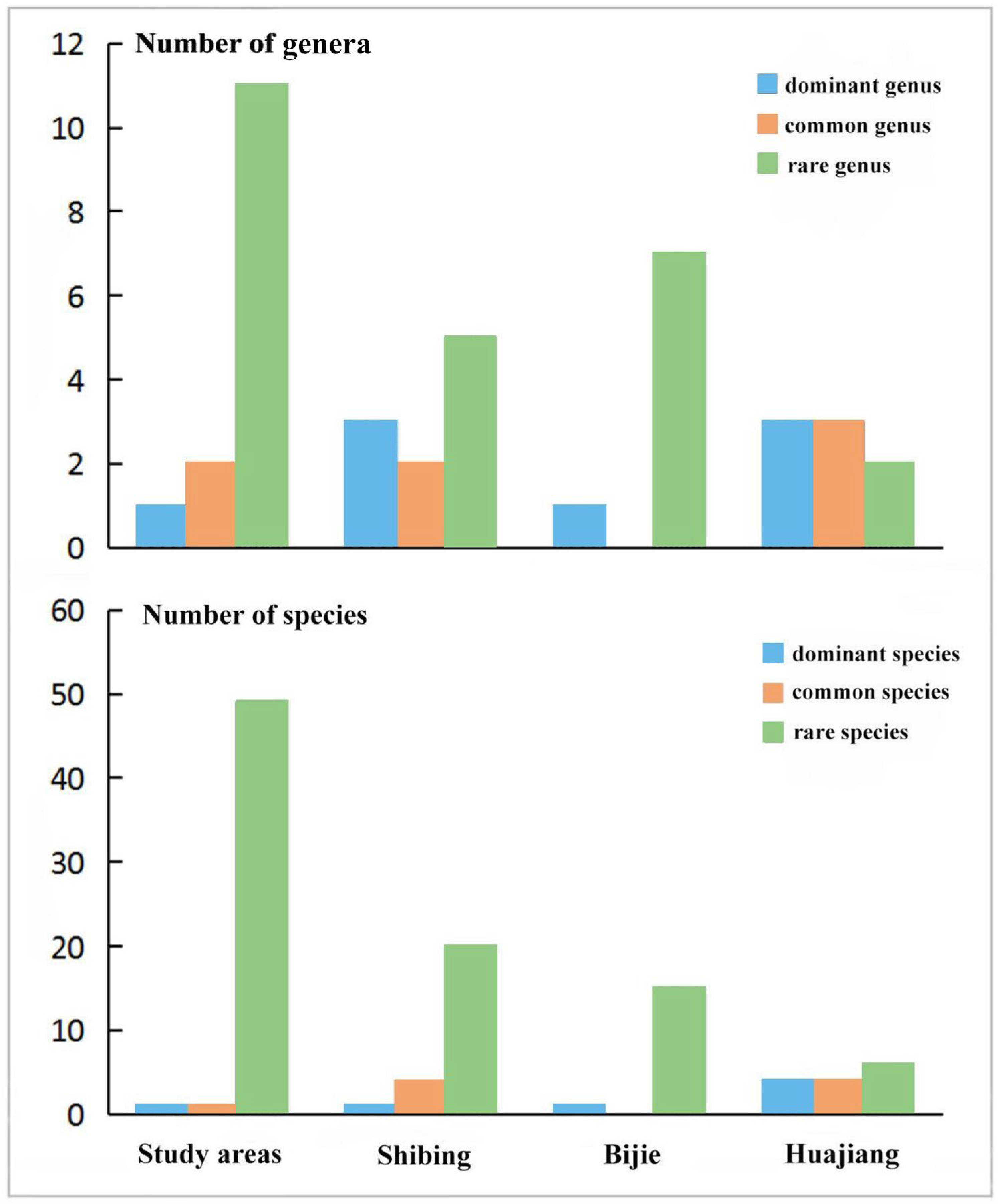
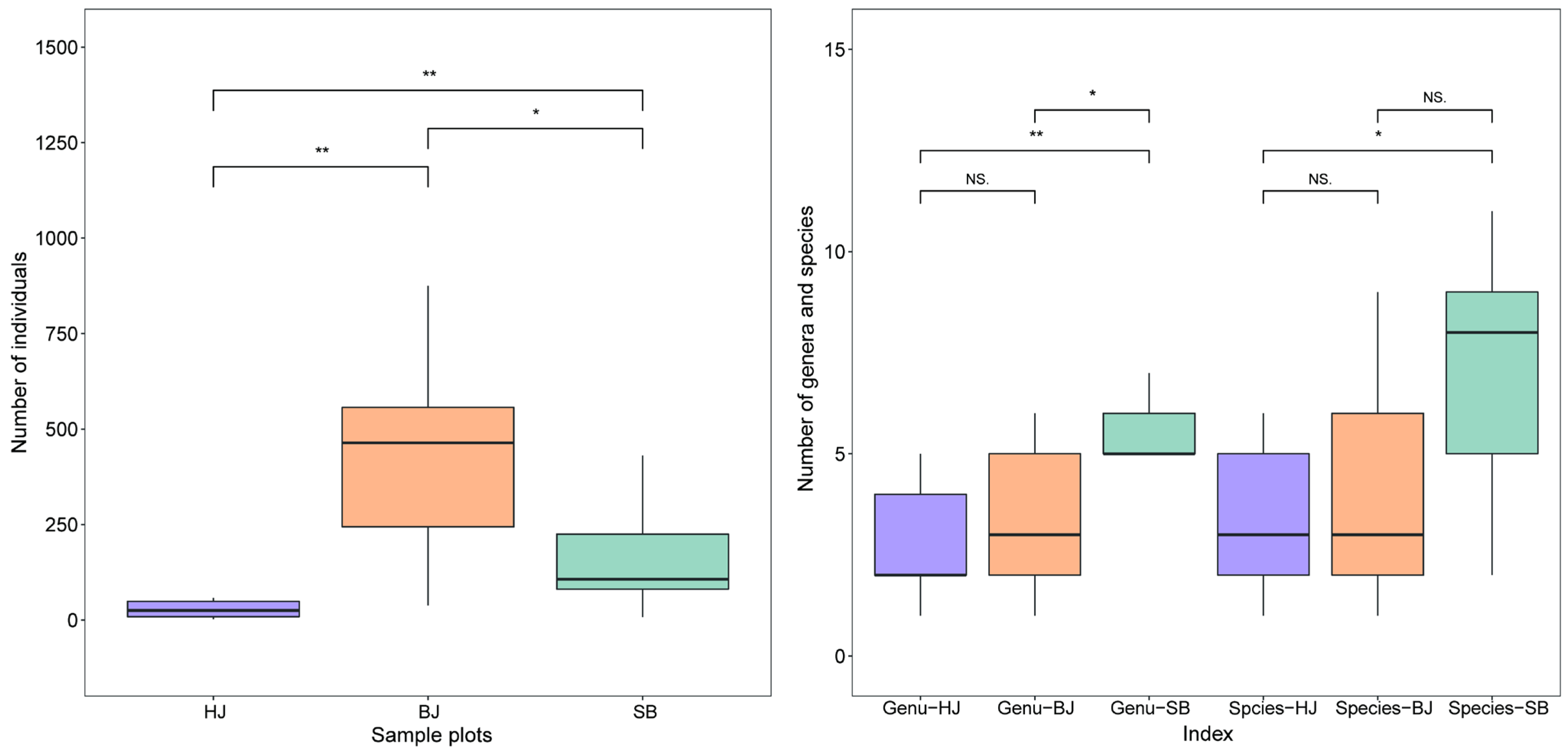
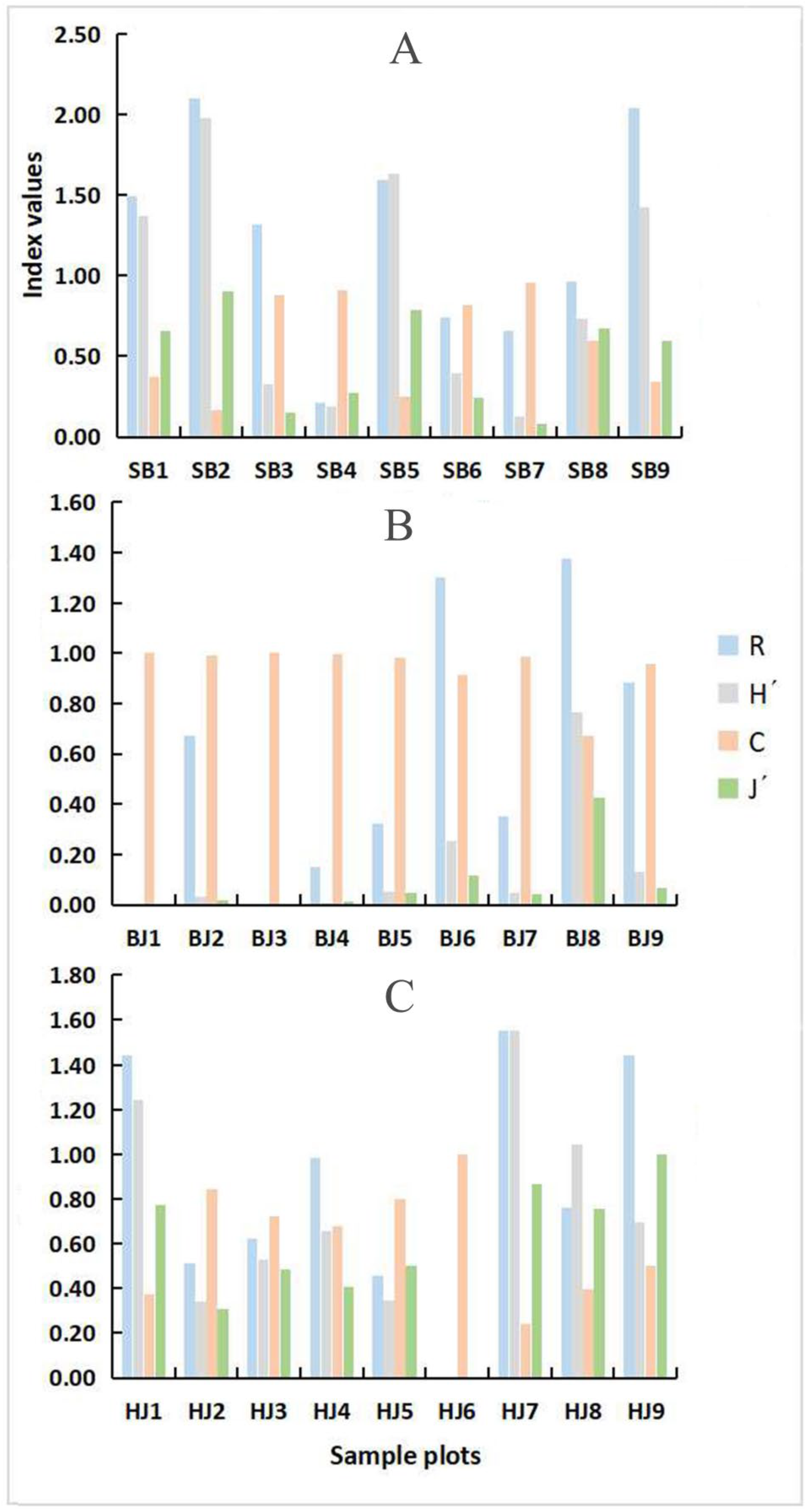
3.2.1. Characteristics of Spatial Distribution and Dominance of Community
3.2.2. Distribution Differences of Community Composition
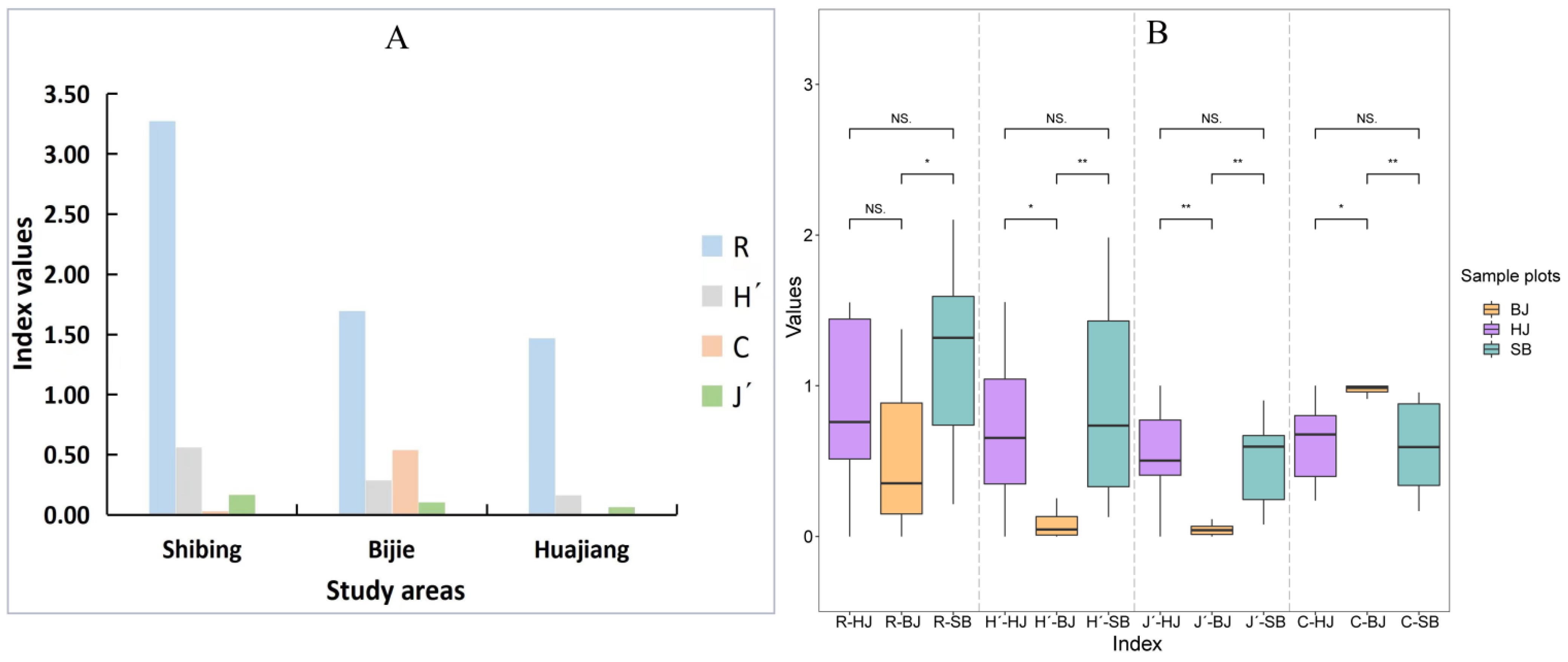
3.2.3. Diversity of Spatial Distribution of Erythroneurine Community
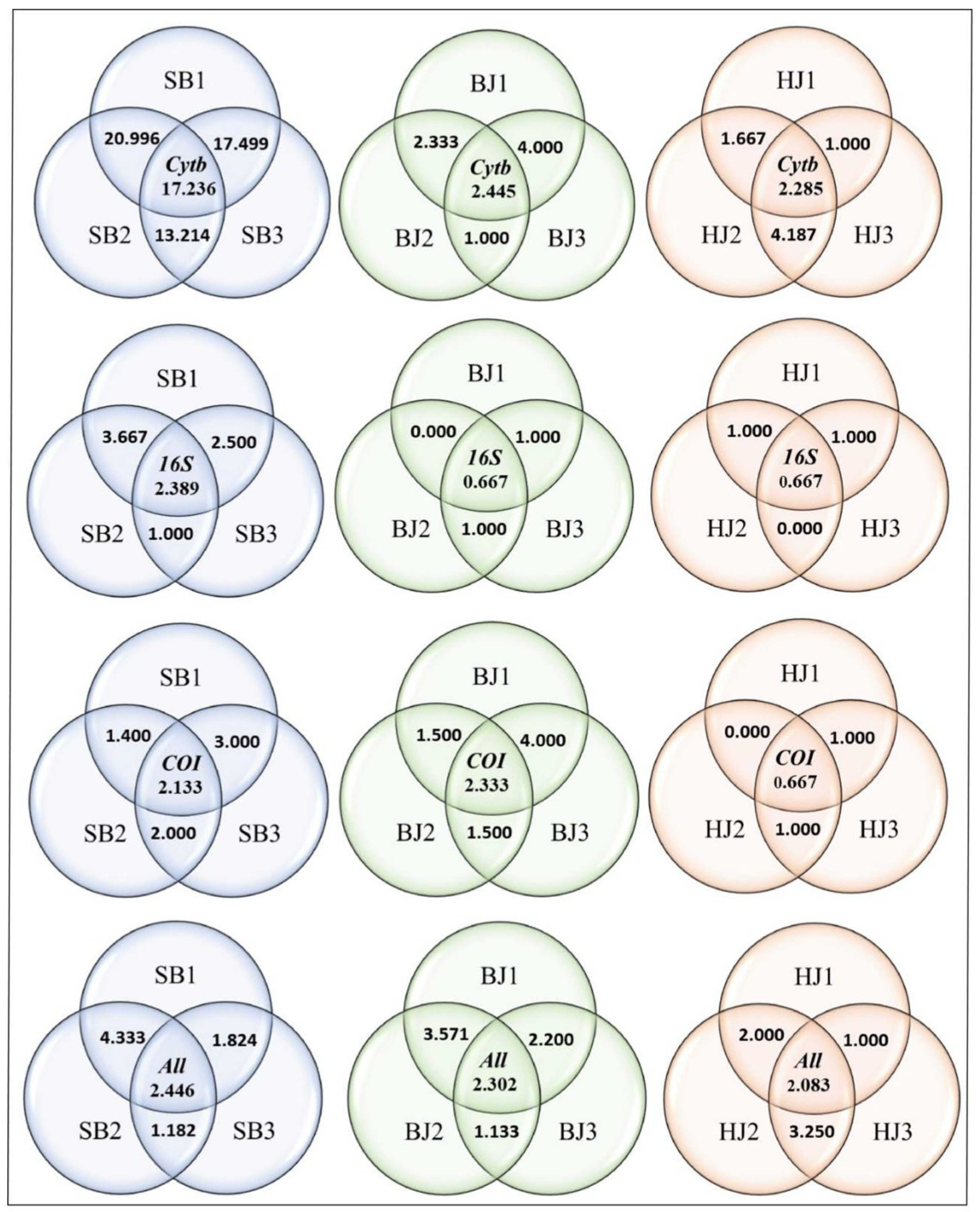
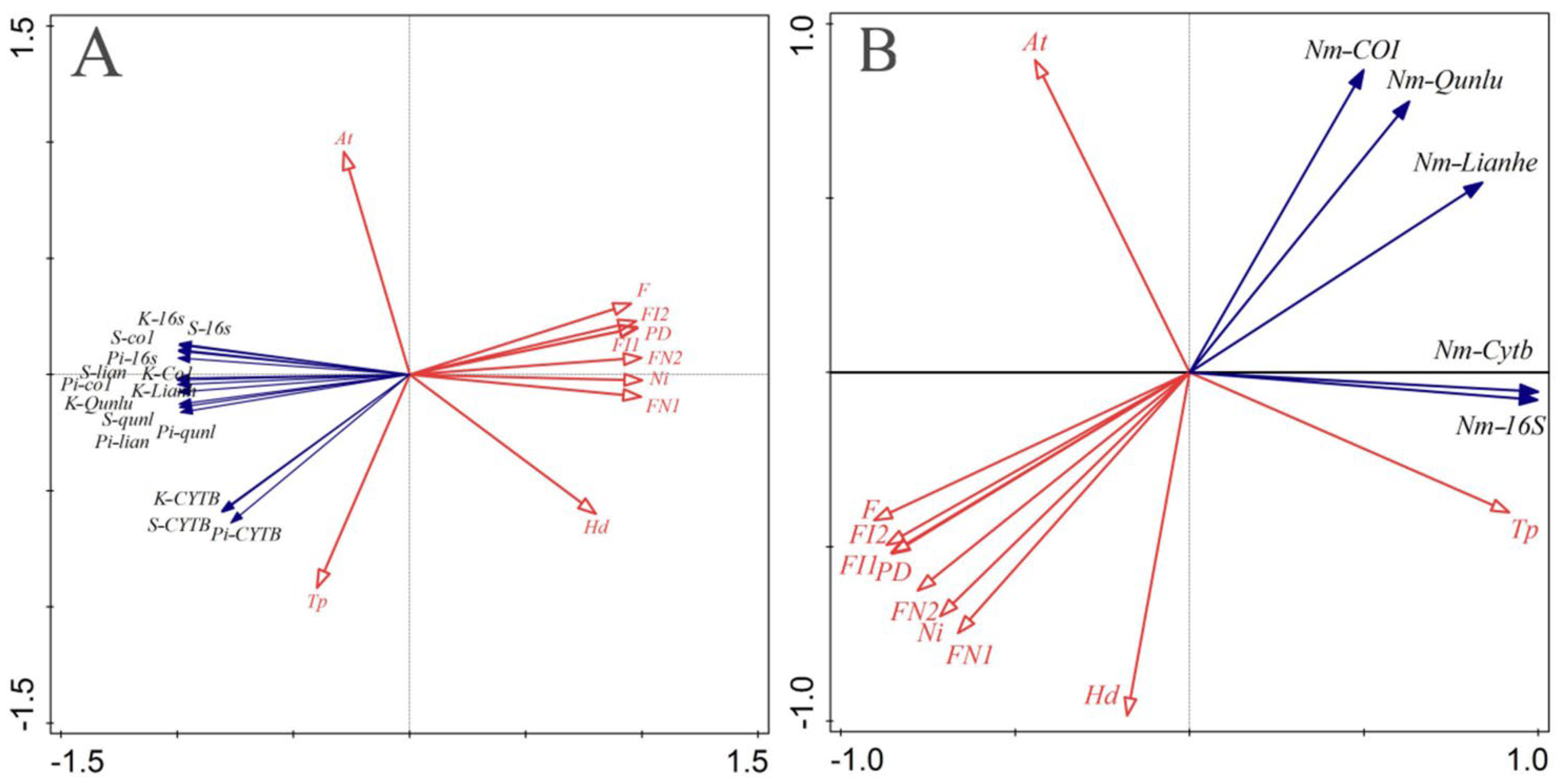
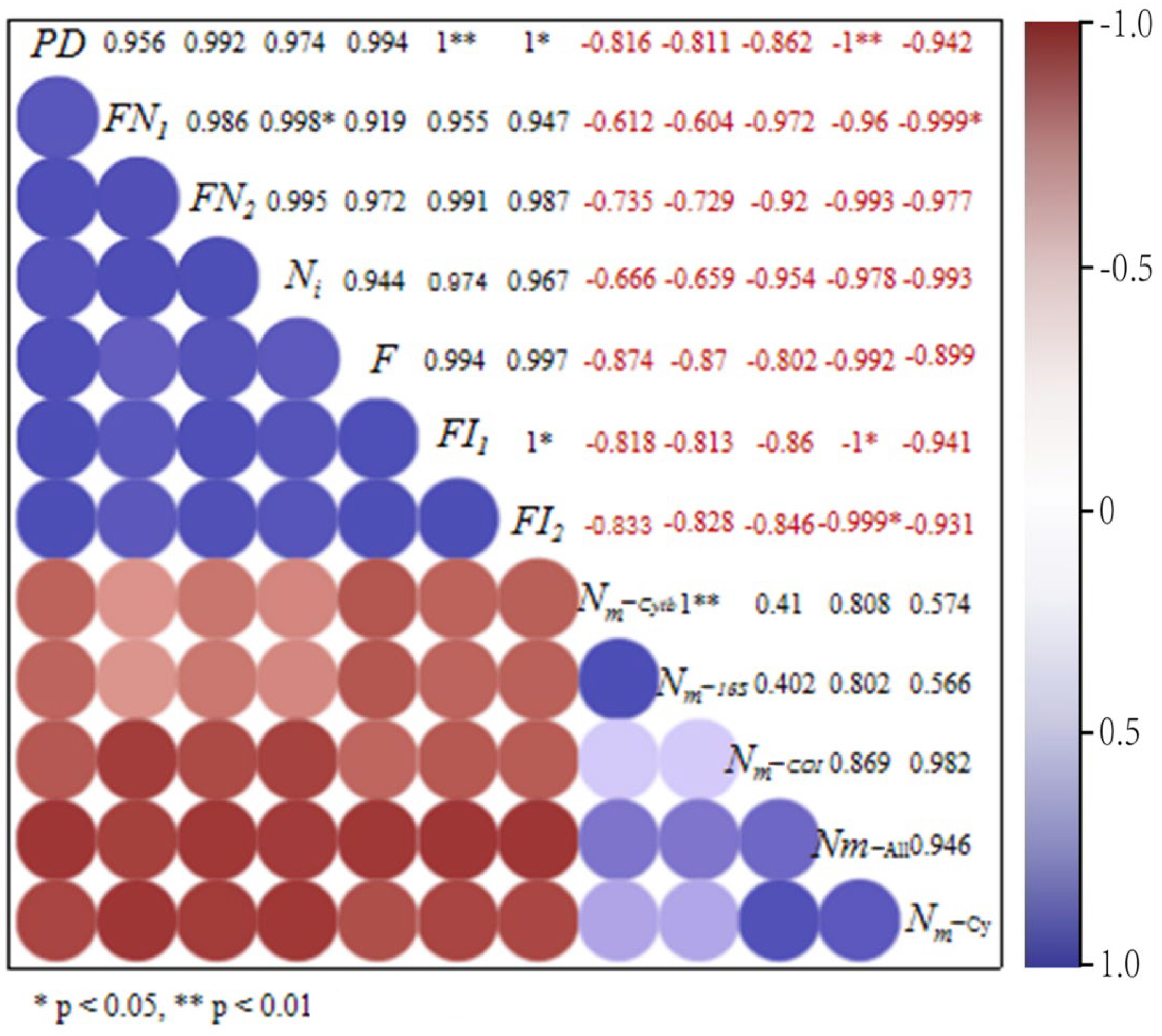
3.3. The Impact of Habitat Fragmentation on Population Gene-Flow
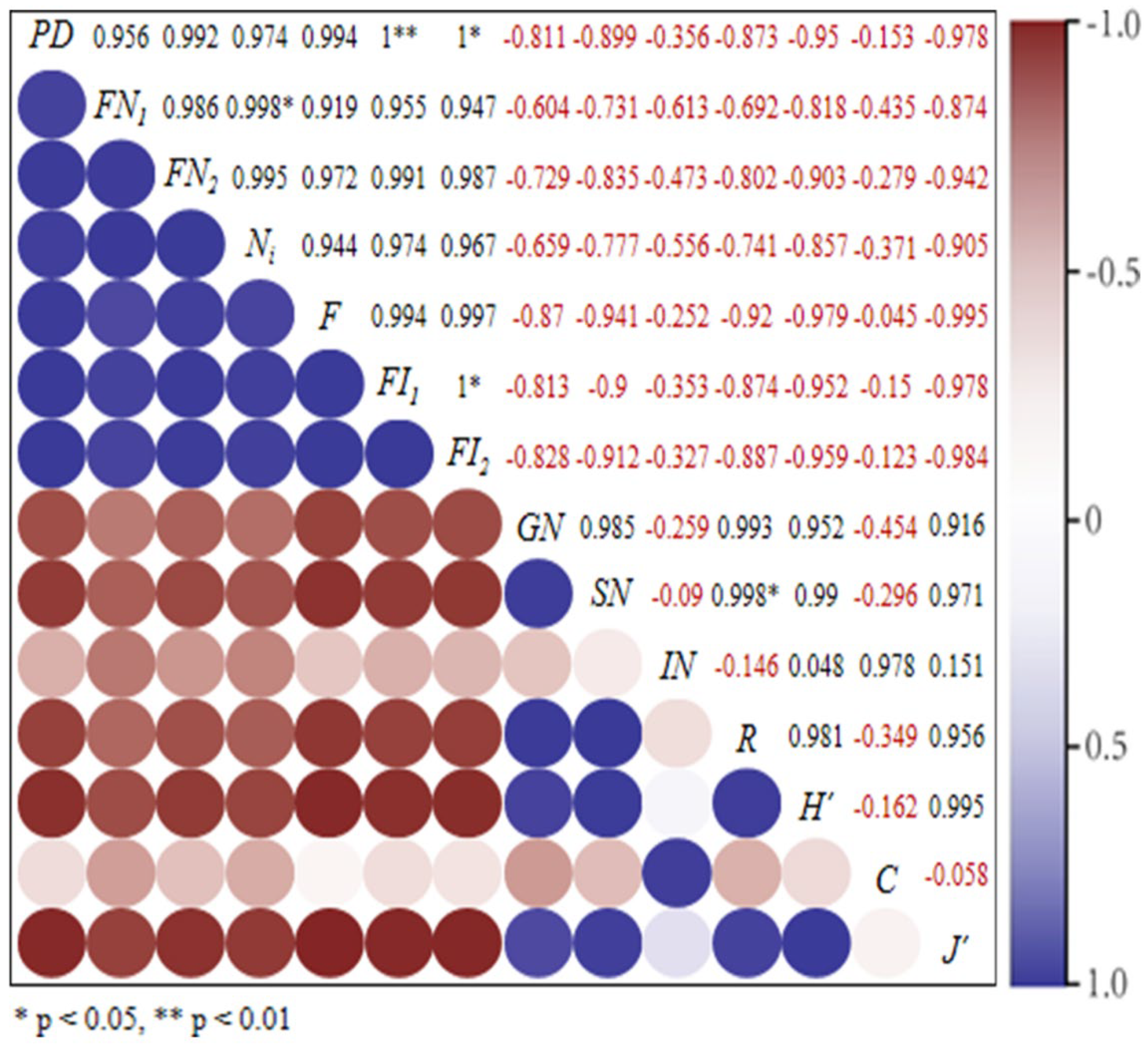
3.4. Correlation Analysis of Erythroneurine Biodiversity and Environmental Factors
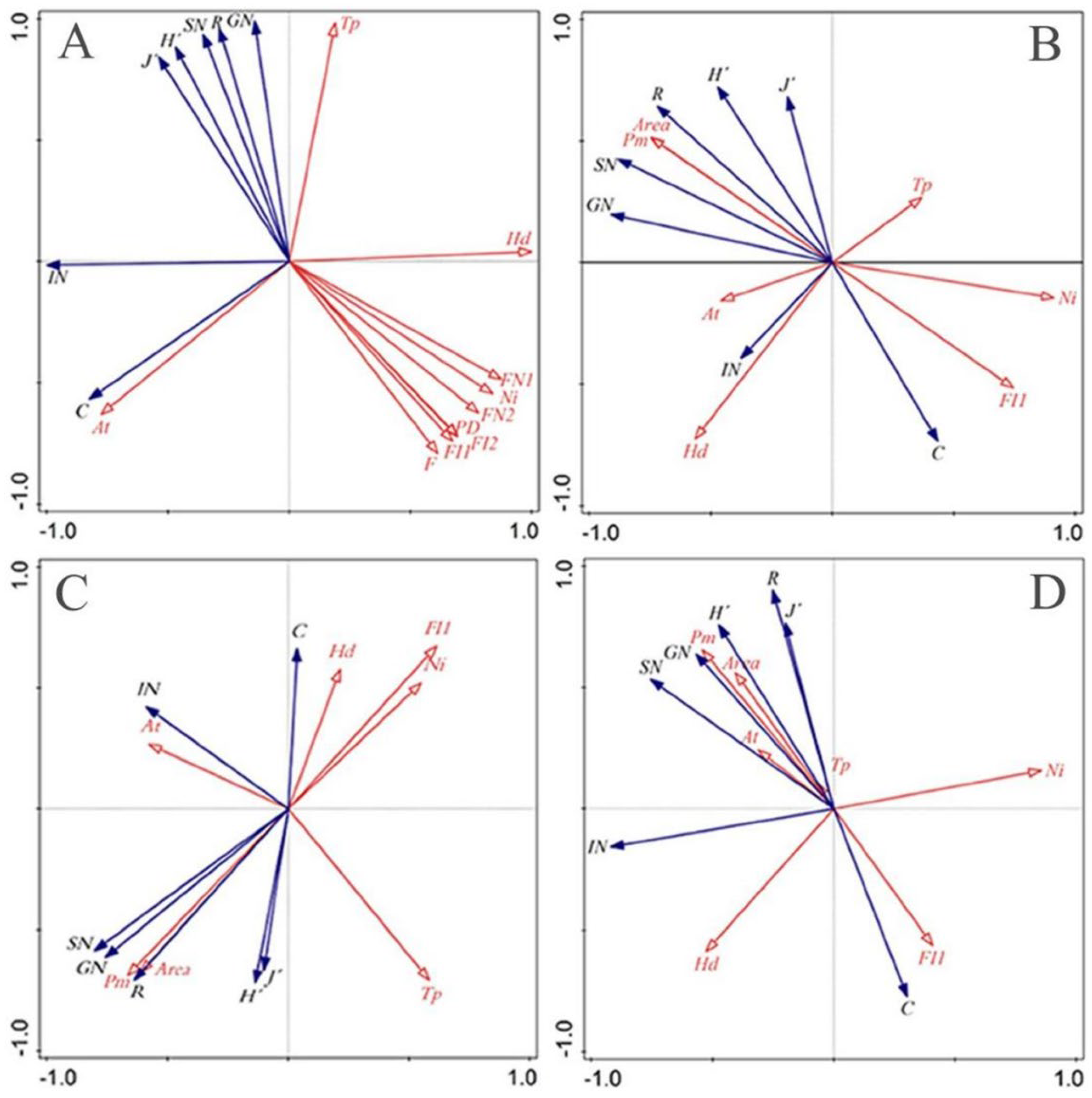
3.4.1. Redundancy Analysis (RDA) of Species Diversity of Erythroneurine and Environmental Factors
3.4.2. Redundant Analysis (RDA) of Genetic Diversity of Erythroneurine and Environmental Factors
3.4.3. Correlation Analysis of Habitat Fragmentation and Erythroneurine Leafhoppers’ Biodiversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berger, J. Persistence of differed-sized populations:an empirical assessment of rapid extinctions in bighorn sheep. Conserv. Biol. 1990, 4, 91–98. [Google Scholar] [CrossRef]
- Rodriguez, A.; Delibes, M. Population fragmentation and extinction in Iberian lynx. Biol. Conserv. 2003, 109, 321–331. [Google Scholar] [CrossRef]
- Gentil, R.; Abeli, T.; Parolo, G.; Ciapeta, S.; Montagnani, C.; Muller, J.V.; Rossi, G.; Citterio, S. Genetic structure of Leucojum aestivum L. in the Po Valley (N-Italy) drives conservation management actions. Conserv. Genet. 2018, 19, 827–838. [Google Scholar] [CrossRef]
- Wu, Z.J.; Li, Y.M. Effects of habitat fragmentation on survival of animal populations. Acta Ecol. Sin. 2003, 11, 2424–2435. [Google Scholar]
- Yang, F.; He, D.H. Effects of habitat fragmentation on plant and insect-insect interactions. Chin. J. Appl. Entomol. 2007, 4, 642–646. [Google Scholar]
- José, A.L.F.; Raphael, J.A.G.V.; Carlos, A.M.; Ferreira, F.F.; de Oliveira, V.M. Effects of habitat fragmentation on biodiversity patterns of ecosystems with resource competition. Phys. A Stat. Mech. Appl. 2021, 564, 125497. [Google Scholar]
- Didham, R.K.; Ghazoul, J.; Stork, N.E.; Davis, A.J. Insects in fragmented forests: A functional approach. Trends Ecol. Evol. 1996, 11, 255–260. [Google Scholar] [CrossRef]
- Heinken, T.; Weber, E. Consequences of habitat fragmentation for plant species: Do we know enough? Perspectives in Plant Ecology. Evol. Syst. 2013, 15, 205–216. [Google Scholar]
- Debinski, D.M.; Holt, R.D. A survey and overview of habitat fragmentation experiments. Conserv. Biol. 2000, 14, 342–355. [Google Scholar] [CrossRef]
- Qin, F.F.; An, S.Q.; Zuo, Y.W.; Xiang, H.J.; Zheng, J.W.; Chen, X.L. Effect of landscape fragmentation on plant populations. Chin. J. Ecol. 2003, 22, 43–48. [Google Scholar]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.H.; Cai, X.W.; Huang, C.M. Habitat Selection and Use of Fran Dois’s Langurs (Trachypithecus francoisi) in Guangxi Province, Fusui Area. Zool. Res. 2010, 31, 421–427. [Google Scholar]
- Cao, Y.S.; Fu, L.S.; Zhou, B.; Hu, W.J.; Li, H. The study of plant diversity in three fragmented habitats in Jian city. J. Jinggangshan Univ. Nat. Sci. 2011, 32, 117–123. [Google Scholar]
- Wang, Y.M.; Xue, Y.D.; Xia, F.Y. Landscape Pattern and Its Fragmentation Evaluation of Habitat of Rhinopithecus bieti in Northwest Yunnan. For. Inventory Plan. 2011, 36, 34–37. [Google Scholar]
- Katherine, E.H.; John, D.A.; Adam, S.H.; Stephens, J.L.; Yang, Z.; Betts, M.G. Using a species-centered approach to predict bird community responses to habitat fragmentation. Landsc. Ecol. 2019, 34, 1919–1935. [Google Scholar]
- Zabel, J.; Tscharntke, T. Does fragmentation of Urtica habitats affect phytophagous and predatory insects differentially? Oecologia 1998, 116, 419–425. [Google Scholar] [CrossRef]
- Liu, C.M.; Lian, Z.M. Comparisons of diversity of grasshopper community in fragmentary forest of Nanniwan. Acta Ecol. Sin. 2003, 23, 1222–1229. [Google Scholar]
- Inara, R.; Bruno, K.C.; Filgueiras, J.P.; Gomes, J.P.; Iannuzzi, L.; Andersen, A.N. Effects of habitat fragmentation on ant richness and functional composition in Brazilian Atlantic forest. Biodivers. Conserv. 2012, 21, 1687–1701. [Google Scholar]
- Yang, H.Y.; Liu, Y.M.; Sang, J.P. Impact of Habitat Fragmentation on Butterfly Species Diversity in Xiaolongshan Forest Region, Gansu. For. Sci. Technol. 2016, 5, 3–7. [Google Scholar]
- Danny, A.P.; Hooftman, R.C.; Billeter, B.S.; Schmid, B.; Diemer, M. Genetic effects of habitat fragmentation on common species of Swiss Fen Meadows. Conserv. Biol. 2004, 18, 1043–1051. [Google Scholar]
- Wang, Z.F.; Gao, S.H.; Tian, S.N.; Fu, S.L.; Ren, H.; Pen, S.L. Genetic structure of Cryptocarya chinensis in fragmented lower subtropical forests in China based on ISSR markers. Biodivers. Sci. 2005, 4, 324–331. [Google Scholar] [CrossRef]
- Samuel, A.C. Effects of habitat loss and fragmentation on amphibians: A review and prospectus. Biol. Conserv. 2006, 128, 231–240. [Google Scholar]
- Hu, S.J.; Yan, X.H.; He, p.; Zhang, C.P. Effects of Habitat Fragmentation on Population Genetic Diversity of Endangered Plant Euonymus chloranthoides Yang. J. Chongqing Norm. Univ. Nat. Sci. 2013, 30, 26–29. [Google Scholar]
- Chen, T. Effects of Landscape Variables on the Genetic Structure and Gene Flow among Plutella xylostella Populations. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2015. [Google Scholar]
- Zhu, S.H. Analysis on the Landscape Pattern Change and Influencing Factors of the Yun Taishan World Natural Heritage Site in Shibing County, Guizhou Province from 2005 to 2015. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2019. [Google Scholar]
- Jaipal, S.C.; Chandra, S.P.; Naiyar, N.; Lemtur, M.; Das, B.; Kumar, S.; Bhatt, B.P. Genetic diversity of mango leafhopper, Amritodus atkinsoni (Hemiptera: Cicadellidae) based on mtCOI gene sequences from India. Mitochondrial DNA Part B 2019, 4, 261–264. [Google Scholar]
- Wang, S.J. The Most Serious Eco-geologically environmental Problem in Southwestern China—Karst Rocky Desertification. Bull. Mineral. Petrol. Geochem. 2003, 2, 120–126. [Google Scholar]
- Wang, S.J. Concept deduction and its connotation of larst rocky desertification. Carsologica Sin. 2002, 21, 31–35. [Google Scholar]
- Yang, X.Y.; Zhou, Z.F.; Zou, C.H. Spatial pattern analysis of karst rocky desertification based on landscape level index. J. Guizhou Norm. Univ. Nat. Sci. 2010, 28, 23–27. [Google Scholar]
- Cao, J.H.; Yuan, D.X.; Tong, L.Q.; Malik, A.; Hui, Y.; Fen, H. An overview of karst ecosystem in southwest China: Current state and future management. J. Resour. Ecol. 2015, 6, 247–256. [Google Scholar]
- Dietrich, C.H. Keys to the families of Cicadomorpha and subfamilies and tribes of Cicadellidae (Hemiptera: Auchenorrhyncha). Fla. Entomol. 2005, 88, 502–517. [Google Scholar] [CrossRef]
- Morris, M.G. Differences between the invertebrate faunas of grazed and ungrazed chalk grassland, IV. Abundance and diversity of Homoptera-Auchenorrhyncha. J. Appl. Entomol. 1971, 8, 37–52. [Google Scholar]
- Guo, H.F. Major tea pests—Advances in research on pseudo-eye leafhopper. Jiangshu Nongye Kexue Jiangsu Agric. Sci. 2011, 1, 132–134. [Google Scholar]
- Chen, X.X.; Gao, Y.R.; Xiong, K.N.; Song, Y.H. Structure characteristics of leafhopper community in Fanjingshan National Nature Reserve. J. Environ. Entomol. 2000, 43, 883–898. [Google Scholar]
- Wu, X.Z. Floristic analysis of pteridophyte in karst forest area of Guizhou Shibing Yuntaishan Nature Reserve. J. Anhui Agric. Sci. 2012, 40, 869–870. [Google Scholar]
- Yang, S.L.; Luo, Q.H. New records and species richness of the family cerambycidae in Yuntaishan Area of Shibing, Guizhou. Guizhou Agric. Sci. 2013, 41, 103–106. [Google Scholar]
- Liu, P.P.; Xiao, H.; Chen, H. Temporal and spatial evolution characteristics of rocky desertification control landscape pattern in Salaxi Demonstration Area in Bijie City. Southwest China J. Agric. Sci. 2020, 33, 2316–2324. [Google Scholar]
- Zhang, Y.Y.; Chen, Q.W. On ecological benefit monitoring of rocky desertification control project in typical demonstration areas: A case study of Salaxi in Bijie and Huajiang in Guanling. J. Leshan Norm. Univ. 2018, 33, 58–67. [Google Scholar]
- Cui, L.; Xiong, K.N.; Guan, Z.H.; Chen, Y.B.; Liu, Z.Q. The model of karst land management based on plant diversity restoration and protection: Examples from Salaxi and Huajiang areas of Guizhou. Carsologica Sin. 2016, 35, 513–524. [Google Scholar]
- Gu, Z.F.; Liu, Q.; Lu, Y.R. Relationships between the rock and soil chemical element contents and rocky desertification difference in karst rocky desertification process: A case study on the Zhenfeng-Guanling Huajiang area of Guizhou Province. Carsologica Sin. 2016, 35, 533–538. [Google Scholar]
- Guo, M.; Ma, M.G.; Xiao, D.N.; Wenbing, Y. Study on landscape fragmentation of arid region oasis based on remote sensing and GIS-Acase study of Jintaoasis. J. Desert Res. 2004, 24, 201–206. [Google Scholar]
- Chen, H.Y.; Bao, Y.X.; Chen, L.; Hu, Z.Y.; Ge, B.M. Effects of habitat fragmentation on gene flow of the black muntjac (Muntiacus crinifrons). Acta Ecol. Sin. 2008, 3, 1109–1119. [Google Scholar]
- Zhang, C.; Wang, D.D.; Guo, L.; Zhao, W.Q.; Wang, S.M.; Zhang, X. Habitat Fragmentation of Distribution of Gagea bulbifera (pall.) Roem. et schult in the north of Tianshan mountains. J. Anhui Agric. Sci. 2015, 43, 189–191. [Google Scholar]
- Zhang, D.Z.; Zhang, S.X.; Ma, Z.F.; Wu, G.S. Effects of habitat fragmentation on seed pest populations and damage rate of Sophora alopecuroides. Ecol. Sci. 2019, 38, 31–35. [Google Scholar]
- Li, Z.Z.; Li, H.; Xing, J.C. Insects (Cephalus) Type Specimens in the Collection of Guizhou University; Guizhou Science and Technology Press: Guiyang, China, 2014. [Google Scholar]
- Song, Y.H.; Li, Z.Z. Erythroneurini and Zyginellini from China (Hemiptera: Cicadellidae: Typhlocybinae); Science and Technology Publishing House: Guiyang, China, 2014; pp. 12–209. [Google Scholar]
- Hong, X.M.; Xin, Y.; Li, J.L. Butterfly diversity and its influencing factors in Saihanwula Nature Reserve. Biodivers. Sci. 2018, 6, 590–600. [Google Scholar] [CrossRef]
- Peet, R.K. The measurement of species of diversity. Annu. Rev. Ecol. Syst. 2014, 5, 285. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, B.Y.; Gao, Y.X.; Ma, Z.X. A study on community characteristics of geometridae and insect faunal in Baishuijiang Nature Reserve of Gansu Province. J. Gansu Sci. 2010, 22, 58–64. [Google Scholar]
- Hu, B.B.; Li, H.H.; Liang, Z.P.; Zhao, T.J.; Ren, X.B. Diversity and fauna of butterflies in Baxian Mountain State Nature Reserves. Acta Ecol. Sin. 2010, 30, 3226–3238. [Google Scholar]
- Ma, Y. Effects of Desert Landscape Habitat Fragmentation on the Ground-Dwelling Beetle Diversity in Ningxia Lingwu Baijitan National Nature Reserve. Master’s Thesis, Ningxia University, Yinchuan, China, 2015. [Google Scholar]
- Hisatomo, I.; Takenari, I.; Hiroshi, T.; Hiroshi, M.; Sueyoshi, M.; Isono, M.; Okabe, K. Responses of community structure, diversity, and abundance of understory plants and insect assemblages to thinning in plantations. For. Ecol. Manag. 2010, 259, 607–613. [Google Scholar]
- Jiang, J.X.; Wang, N.F.; Ji, X.Y.; Dan, J.G. Diversity and stability of arthropod community in peach orchard under effects of ground cover vegetation. Chin. J. Appl. Ecol. 2011, 22, 2303–2308. [Google Scholar]
- Li, L.L. Study of the Relation between Grasshopper and Vegetation Community in the Upper Reaches of Hei River. Master’s Thesis, Northwest Normal University, Lanzhou, China, 2012. [Google Scholar]
- Gao, Y.R. Research on the Relationship between Fanjing Mountain Leafhopper Species Diversity and Its Host Plant. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2019. [Google Scholar]
- Zhao, Z.Q.; Xiong, K.N.; Song, Y.H.; Luo, D.; Zhao, P.D. The community structure and its diversity dynamic change of Cicadellidae in the karst plateau mountainous rocky desertification areas. J. Environ. Entomol. 2015, 37, 498–506. [Google Scholar]
- Collinge, S.K. Effects of grassland fragmentation on insect species loss, colonization, and movement patterns. Ecology 2000, 81, 2211–2226. [Google Scholar] [CrossRef]
- Krauss, J.; Steffan, D.I.; Tscharntke, T. How does landscape context contribute to effects of habitat fragmentation on diversity and population density of butterflies? J. Biogeogr. 2003, 30, 889–900. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.H.; Wang, Y.; He, H.D.; Zhang, R.; Zhu, M.M.; Dong, F.L. Effects of habitat loss and fragmentation on species loss and colonization of insect communities in experimental alfalfa landscapes. Biodivers. Sci. 2011, 19, 453–462. [Google Scholar]
- Gu, X.Y. Impacts of Habitat Fragmentation on Genetic Diversity and Differentiation of Liquidambar formosana Populations. Master’s Thesis, East China Normal University, Shanghai, China, 2016. [Google Scholar]
- Meher, A.O.; Marcin, N.; Sarah, L.B.; Klingeman, W.E.; Zobel, J.M.; Trigiano, R.N.; Hadziabdic, D. Habitat fragmentation influences genetic diversity and differentiation: Fine-scale population structure of Cercis canadensis (eastern redbud). Ecol. Evol. 2020, 10, 3655–3670. [Google Scholar]
- Yi, W.Y.; Su, W.C.; Wu, K.H.; Zhao, W.Q. Ecological health diagnostic of Yuntai Mountain area, Shibing County, Guizhou Province, China based on the view of land use. Earth Environ. 2015, 43, 552–557. [Google Scholar]
- Zhu, Y.; Li, J.; Pang, H.Y.; Wang, Z.; Du, L.; Xiong, S.L.; Cai, T. Genetic differentiation of 11 geographical populations of Empoasca vitis (Gothe) (Hemiptera:Cicadellidae) based on mtDNA cyt b gene. Biot. Resour. 2019, 41, 228–235. [Google Scholar]
- Wang, C.L.; Chen, H.; Xiao, H.; Zhang, H.M.; Li, N.Z.; Guo, C.; Chen, J.; Wei, Q. Diversity and habitat selection of amphibians in rocky desertification area in northwestern Guizhou. Biodivers. Sci. 2020, 28, 485–495. [Google Scholar] [CrossRef]
- Guan, Z.H.; Xiong, K.N.; Chen, Y.B.; Gu, Z.K.; Cui, L.; Zhao, P.D. Studies on land arrangement in karst rocky desertification area in basis of biodiversity conservation with a special refrence to Salaxi Demonstration Area, Bijie, Guizhou. J. Xinyang Norm. Univ. Nat. Sci. Ed. 2015, 28, 362–367. [Google Scholar]
- Chen, L.S.; Xiong, K.N.; Chen, Q.W.; Shu, T.; Wu, J. Response mechanism of soil conservation function to rocky desertification under eco-environmental harness of karst areas. Resour. Environ. Yangtze Basin 2020, 29, 499–510. [Google Scholar]
- Wang, X.Y. Evaluation of Ecological Environment Quality in Rocky Desertification Area Based on Remote Sensing Ecological Index-a Case Study of Huajiang Demonstration Area in Guizhou. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2019. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Study Areas | Total | Abundance | |||||
|---|---|---|---|---|---|---|---|---|---|
| SB (1580) | BJ (5245) | HJ (239) | Ra | ||||||
| Anufrievia | Anufrievia sp. nov.-1 | 1 | + | 0 | 0 | 1 | + | ||
| Anufrievia parisakazu | 1 | + | 0 | 0 | 1 | + | |||
| Anufrievia sp.nov.-2 | 0 | 2 | + | 0 | 2 | + | |||
| Arboridia | Arboridia sp. fm-1 | 1 | + | 0 | 0 | 1 | + | ||
| Arboridia sp. fm-2 | 1 | + | 0 | 0 | 1 | + | |||
| Arboridialunula | 16 | + | 0 | 0 | 16 | + | |||
| Arboridia sp. nov.-1 | 9 | + | 0 | 0 | 9 | + | |||
| Arboridia sp. fm-3 | 0 | 1 | + | 0 | 1 | + | |||
| Arboridiaechinata | 0 | 1 | + | 0 | 1 | + | |||
| Arboridia sp. fm-4 | 0 | 0 | 2 | + | 2 | + | |||
| Diomma | Diomma pincersa | 1 | + | 0 | 0 | 1 | + | ||
| Elbelus | Elbelus tripunctatus | 0 | 0 | 44 | +++ | 44 | + | ||
| Empoascanara | Empoascanara sp. fm-1 | 1 | + | 0 | 0 | 1 | + | ||
| Empoascanara sp. fm-2 | 1 | + | 0 | 0 | 1 | + | |||
| Empoascanara sp. fm-3 | 1 | + | 0 | 0 | 1 | + | |||
| Empoascanara sp. fm-4 | 1 | + | 0 | 0 | 1 | + | |||
| Empoascanara sp. fm-5 | 0 | 2 | + | 0 | 2 | + | |||
| Empoascanara sipra | 1267 | +++ | 5185 | +++ | 13 | +++ | 6465 | +++ | |
| Empoascanara dwalata | 4 | + | 0 | 0 | 4 | + | |||
| Empoascanara gracilis | 9 | + | 0 | 0 | 9 | + | |||
| Empoascanara sp. fm-6 | 0 | 0 | 4 | ++ | 4 | + | |||
| Empoascanara mai | 0 | 0 | 1 | + | 1 | + | |||
| Empoascanara sp. nov.-1 | 0 | 0 | 1 | + | 1 | + | |||
| Kapsa | Kapsa sp. fm-1 | 1 | + | 0 | 0 | 1 | + | ||
| Kapsa alba | 0 | 3 | + | 0 | 3 | + | |||
| Kapsa arca | 0 | 1 | + | 0 | 1 | + | |||
| Kapsa dolka | 0 | 12 | + | 0 | 12 | + | |||
| Kapsa sp. nov.-1 | 0 | 0 | 5 | ++ | 5 | + | |||
| Kapsa dolka | 0 | 0 | 2 | + | 2 | + | |||
| Mitjaevia | Mitjaeviadworakowskae | 33 | ++ | 0 | 0 | 33 | + | ||
| Mitjaevia aurantiaca | 6 | + | 0 | 0 | 6 | + | |||
| Mitjaevia sp. fm-1 | 1 | + | 0 | 0 | 1 | + | |||
| Mitjaevia sp. fm-2 | 2 | + | 0 | 0 | 2 | + | |||
| Mitjaevia shibingensis | 69 | ++ | 0 | 0 | 69 | + | |||
| Mitjaevia protuberanta | 19 | ++ | 0 | 0 | 19 | + | |||
| Mitjaevia diana | 0 | 12 | + | 0 | 12 | + | |||
| Mitjaevia sp. nov.-1 | 0 | 15 | + | 0 | 15 | + | |||
| Mitjaevia sp. nov.-2 | 0 | 1 | + | 0 | 1 | + | |||
| Mitjaevia sp. nov.-3 | 0 | 1 | + | 0 | 1 | + | |||
| Mitjaevia sp. fm-3 | 0 | 3 | + | 0 | 3 | + | |||
| Mitjaevia sp. fm-4 | 0 | 0 | 1 | + | 1 | + | |||
| Salka | Salka sawna | 19 | ++ | 4 | + | 0 | 23 | + | |
| Seriana | Seriana bacilla | 100 | ++ | 0 | 142 | +++ | 242 | ++ | |
| Seriana ochrata | 0 | 0 | 12 | +++ | 12 | + | |||
| Tautoneura | Tautoneura albida | 0 | 1 | + | 0 | 1 | + | ||
| Thaia | Thaia sp. fm-1 | 0 | 1 | + | 0 | 1 | + | ||
| Thaia sp. fm-2 | 0 | 0 | 3 | ++ | 3 | + | |||
| Thaia sp. fm-3 | 0 | 0 | 1 | + | 1 | + | |||
| Ziczacella | Ziczacella steggerdai | 7 | + | 0 | 0 | 7 | + | ||
| Erythroneurini new-1 | Erythroneurini Gen. new-1. sp. nov. | 9 | + | 0 | 0 | 9 | + | ||
| Erythroneurini new-2 | Erythroneurini Gen. new-2. sp. nov. | 0 | 0 | 8 | ++ | 8 | + | ||
| Genes | Study Areas | Length bp | Nucleotide Diversity Pi | Average Number of Nucleotide Differences K | Number of Polymorphic (Segregating) Sites S | Haplotypes H |
|---|---|---|---|---|---|---|
| Cytb | SB | 659 | 0.0111 | 7.333 | 11 | 3 |
| BJ | 641 | 0.0052 | 3.333 | 5 | 3 | |
| HJ | 630 | 0.0042 | 2.667 | 4 | 3 | |
| 16S | SB | 493 | 0.0095 | 4.667 | 7 | 3 |
| BJ | 486 | 0.0082 | 4.000 | 6 | 2 | |
| HJ | 476 | 0.0014 | 0.667 | 1 | 2 | |
| COI | SB | 683 | 0.0059 | 4.000 | 6 | 3 |
| BJ | 714 | 0.0047 | 3.333 | 5 | 2 | |
| HJ | 683 | 0.0010 | 0.667 | 1 | 2 | |
| Cytb +16S + COI | SB | 1835 | 0.0288 | 16.008 | 24 | 3 |
| BJ | 1841 | 0.0192 | 10.672 | 16 | 3 | |
| HJ | 1789 | 0.0072 | 4.002 | 6 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Jiang, J.; Zhang, N.; Yang, X.; Chi, Y.; Song, Y. Effects of Habitat Fragmentation on the Population Structure and Genetic Diversity of Erythroneurini in the Typical Karst Rocky Ecosystem, Southwest China. Insects 2022, 13, 499. https://doi.org/10.3390/insects13060499
Chen X, Jiang J, Zhang N, Yang X, Chi Y, Song Y. Effects of Habitat Fragmentation on the Population Structure and Genetic Diversity of Erythroneurini in the Typical Karst Rocky Ecosystem, Southwest China. Insects. 2022; 13(6):499. https://doi.org/10.3390/insects13060499
Chicago/Turabian StyleChen, Xiaoxiao, Jia Jiang, Ni Zhang, Xiao Yang, Yongkuan Chi, and Yuehua Song. 2022. "Effects of Habitat Fragmentation on the Population Structure and Genetic Diversity of Erythroneurini in the Typical Karst Rocky Ecosystem, Southwest China" Insects 13, no. 6: 499. https://doi.org/10.3390/insects13060499
APA StyleChen, X., Jiang, J., Zhang, N., Yang, X., Chi, Y., & Song, Y. (2022). Effects of Habitat Fragmentation on the Population Structure and Genetic Diversity of Erythroneurini in the Typical Karst Rocky Ecosystem, Southwest China. Insects, 13(6), 499. https://doi.org/10.3390/insects13060499