
Biological Control Options for the Golden Twin-Spot Moth, Chrysodeixis chalcites (Esper) (Lepidoptera: Noctuidae) in Banana Crops of the Canary Islands




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Natural Parasitism
2.2. Efficacy of T. achaeae as Biological Control Agent
2.2.1. Experimental Design
2.2.2. Sampling
2.3. Statistics
3. Results
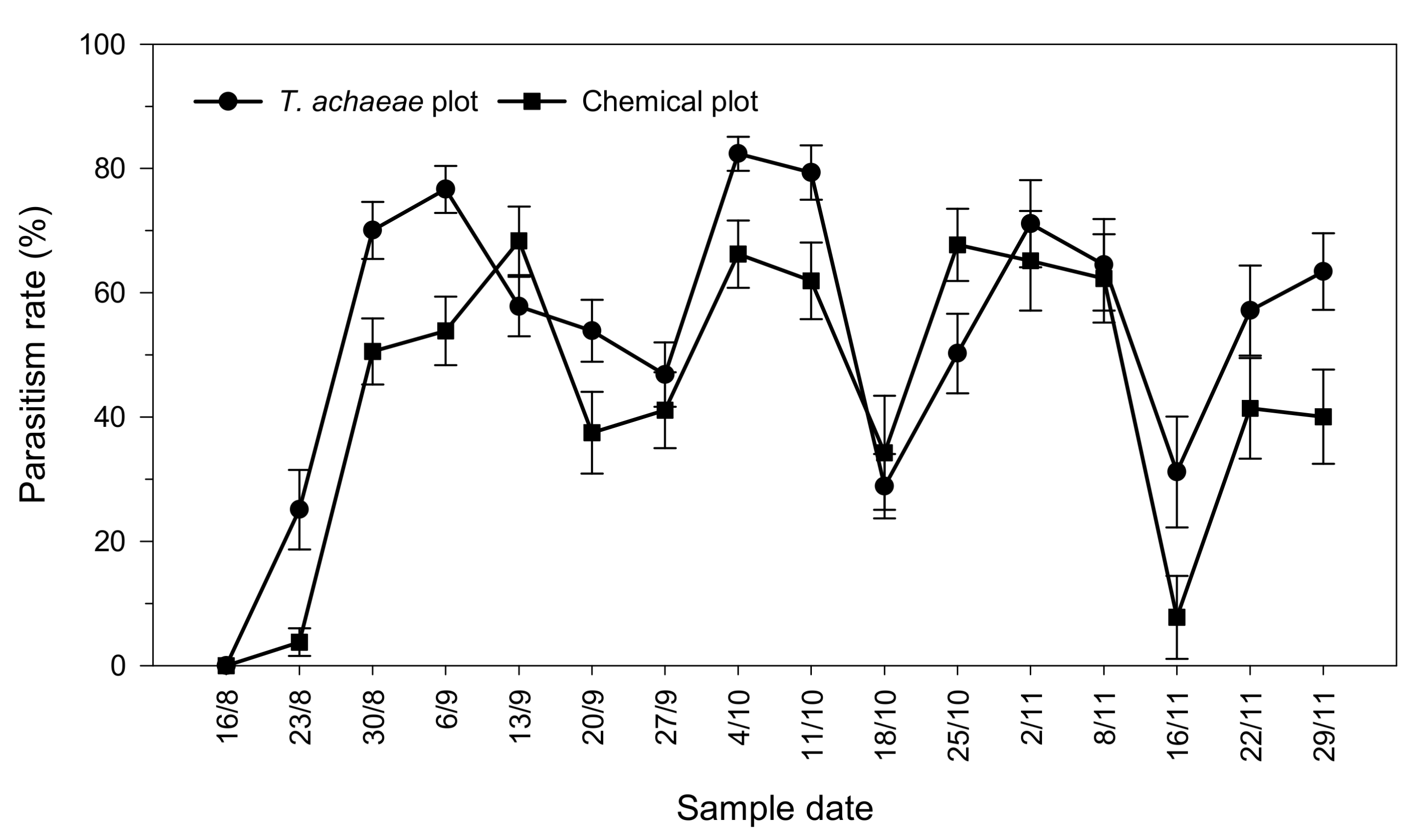
3.1. Natural Parasitism
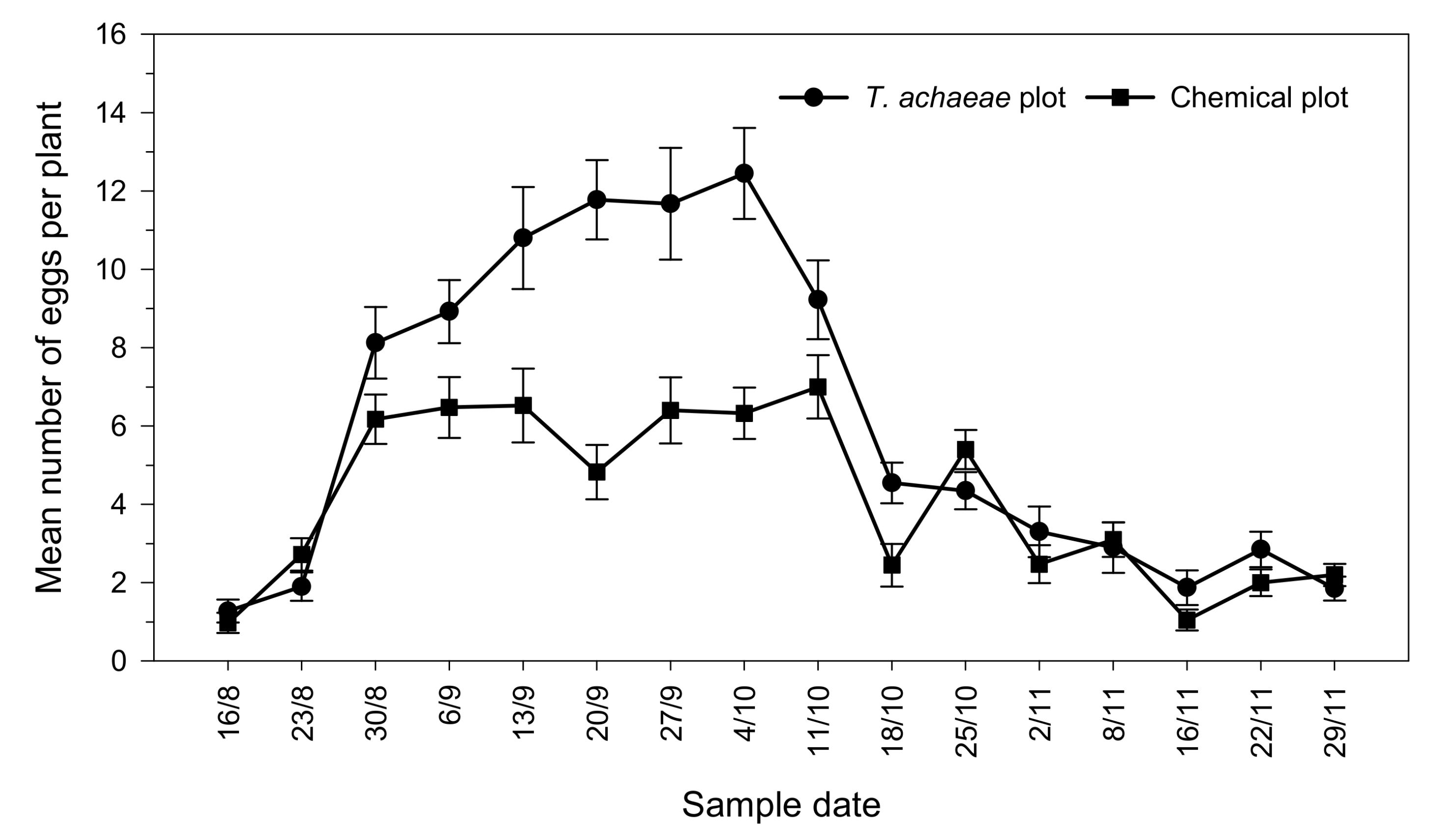
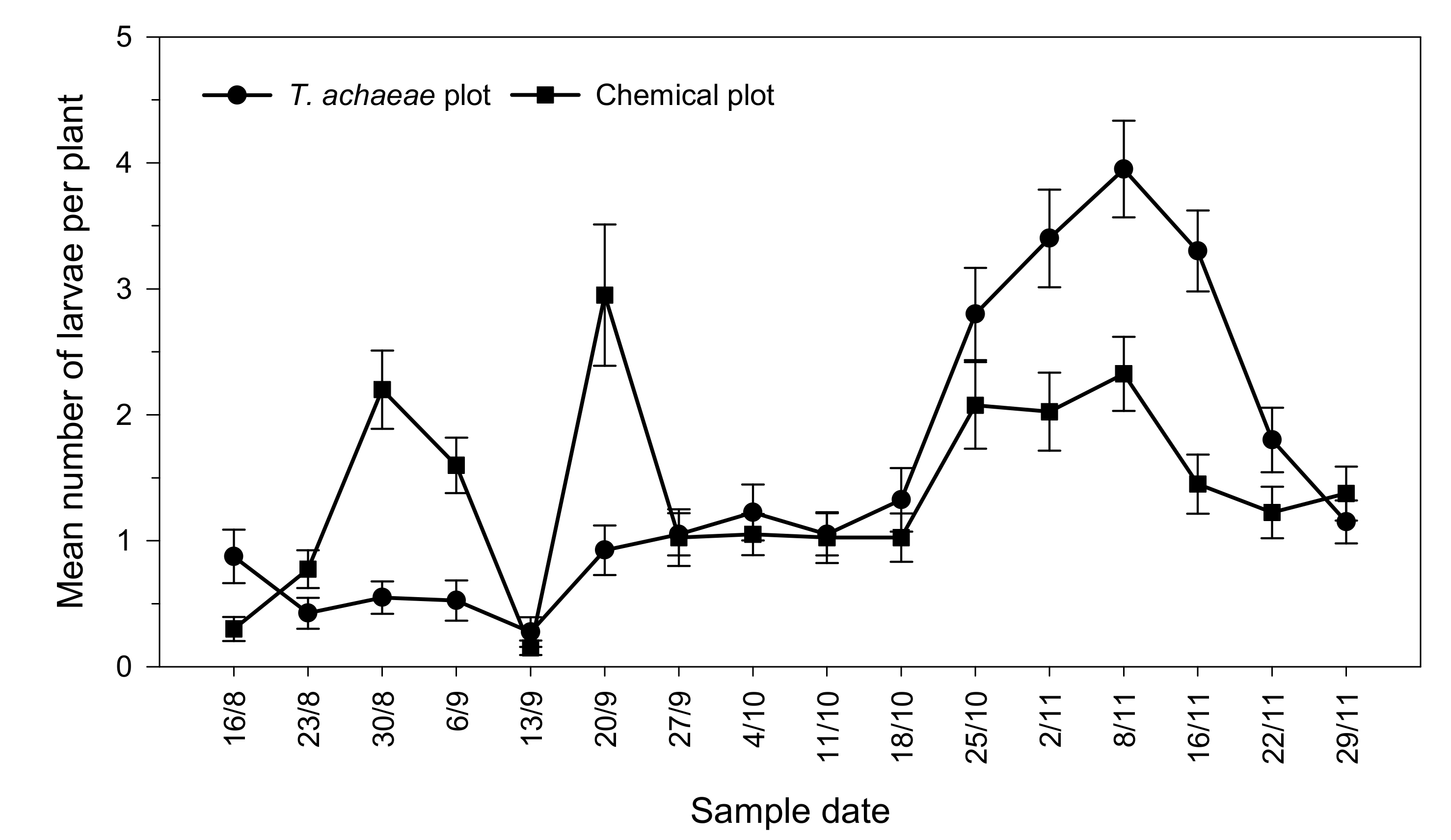
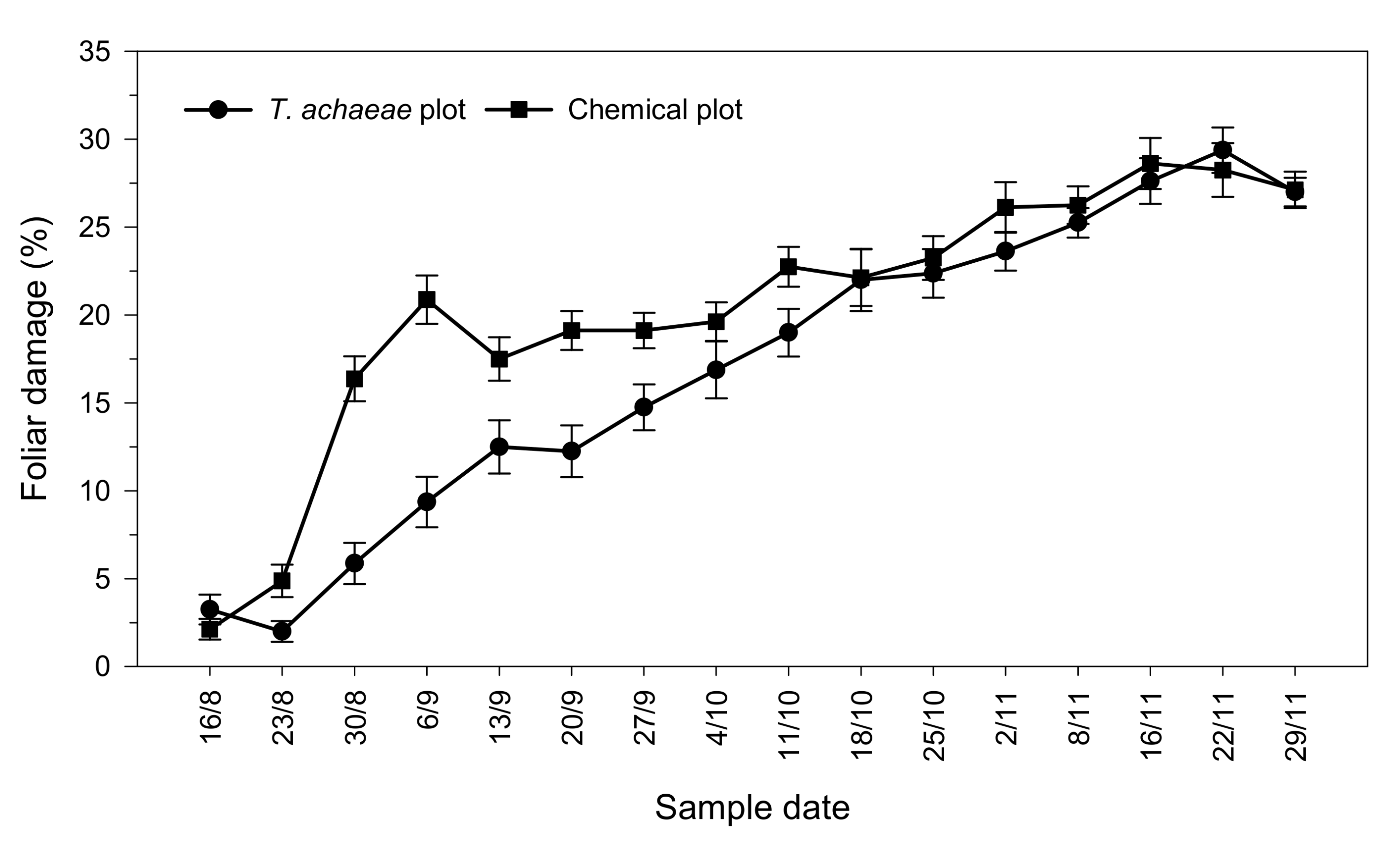
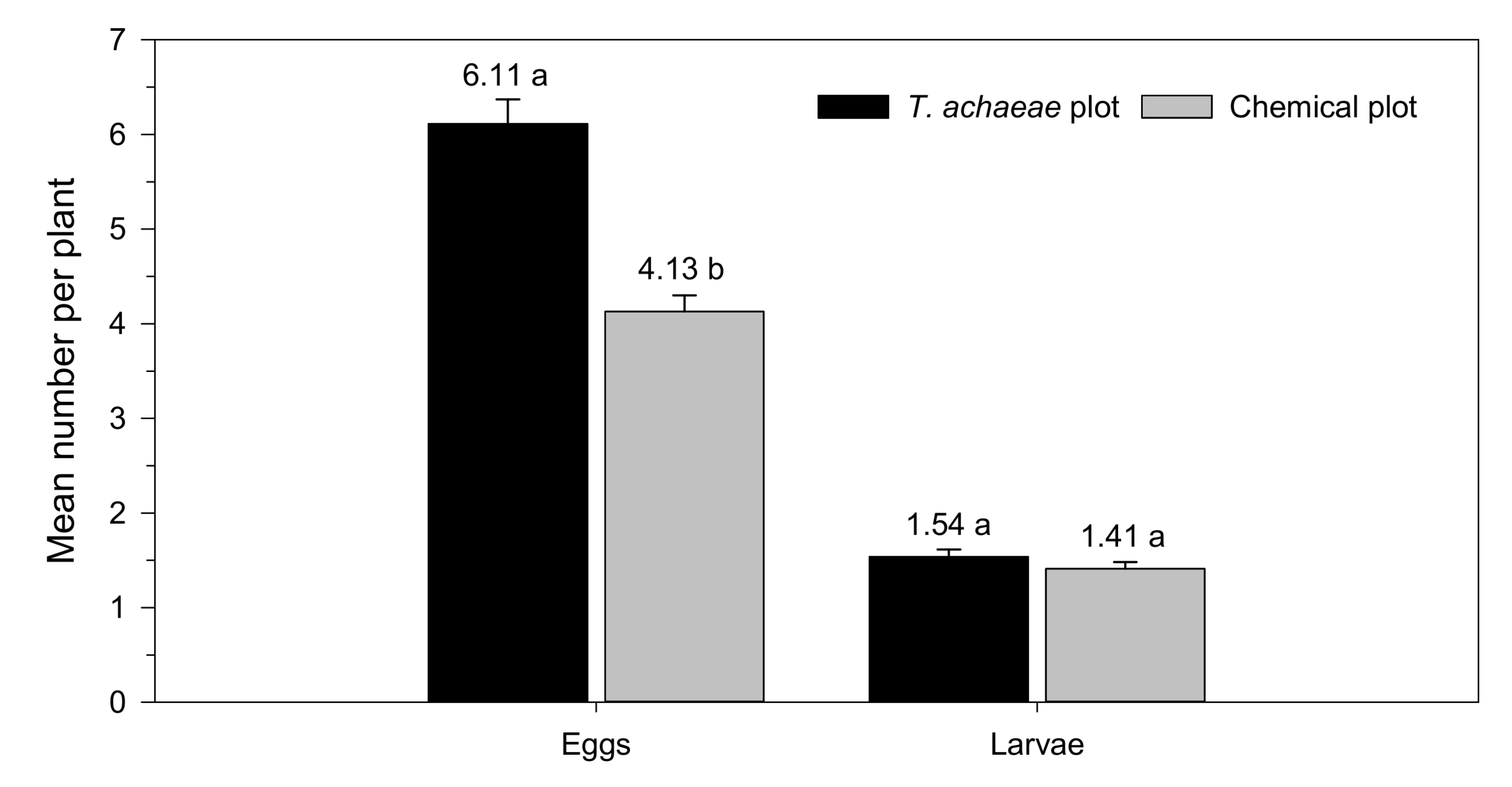
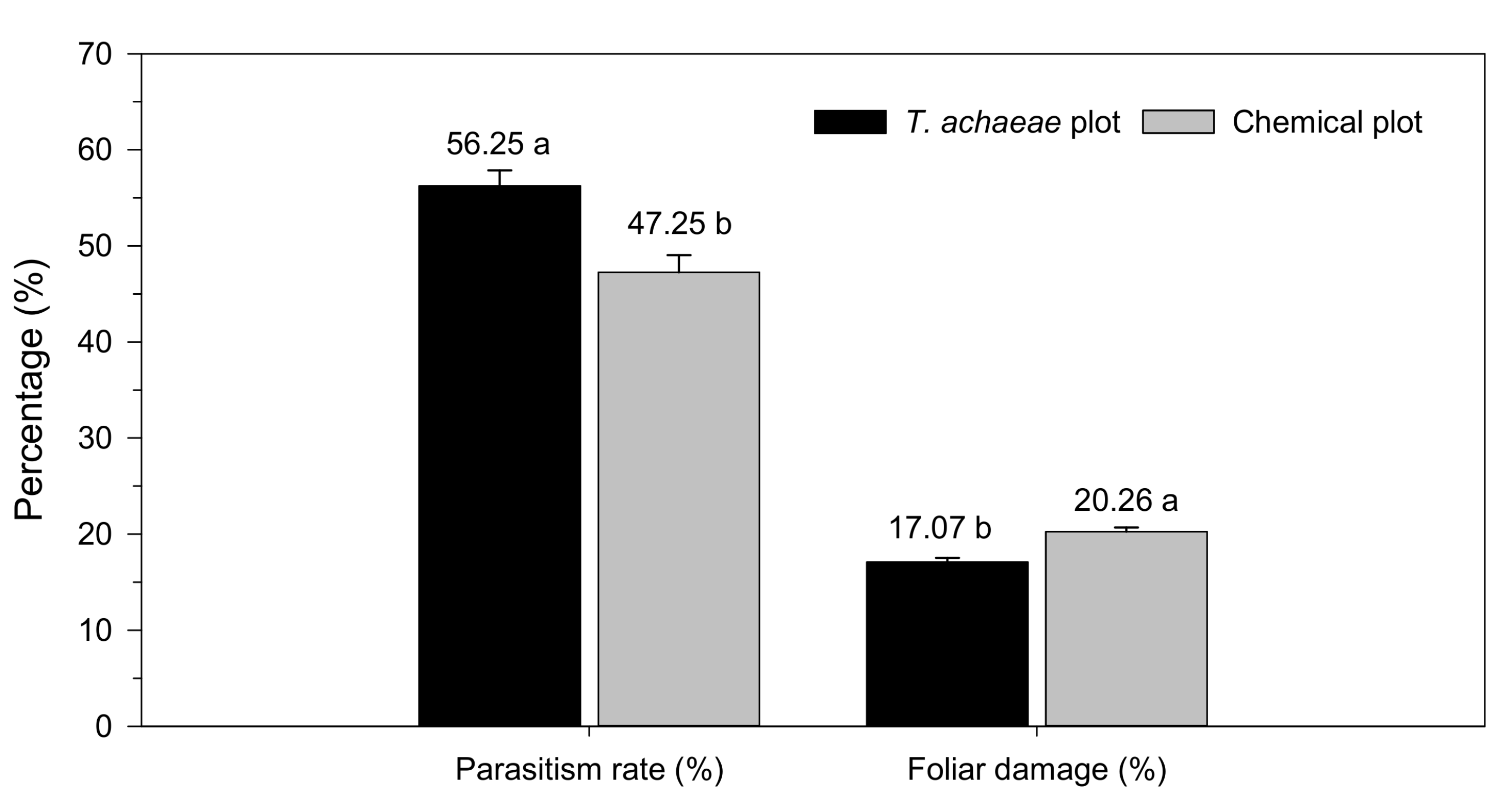
3.2. Efficacy of T. achaeae as Biological Control Agent
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, J.C.; Galán-Saúco, V. Bananas and Plantains, 2nd ed.; CAB International: London, UK, 2010. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization (FAO) of the United Nations. Available online: http://www.fao.org/faostat/es/#home (accessed on 29 June 2021).
- MAPA. Statistical Yearbook 2021; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2021.
- Perera, S.; Molina, M.J. Plagas y enfermedades en el cultivo ecológico de la platanera. In El Cultivo Ecológico de la Platanera en Canarias, 1st ed.; Nogueroles, C., Líbano, J., Eds.; Gabinete de Proyectos Agroecológicos S.L.: Tenerife, Spain, 1997; pp. 70–118. [Google Scholar]
- del Pino, M.; Cabello, T.; Hernández-Suárez, E. Age-Stage, Two-Sex Life table of Chrysodeixis chalcites (Lepidoptera: Noctuidae) at constant temperatures on semi-synthetic diet. Environ. Entomol. 2020, 49, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, E.G.; Hernández-Suárez, E.; Simón, O.; Williams, T.; Caballero, P. Chrysodeixis chalcites, a pest of banana crops on the Canary Islands: Incidence, economic losses and current control measures. Crop Prot. 2018, 108, 137–145. [Google Scholar] [CrossRef]
- CABI. Chrysodeixis chalcites (Esper). In Invasive Species Compendium; CAB International: Wallingford, UK; Available online: www.cabi.org/isc (accessed on 29 June 2021).
- del Pino, M.; Carnero, A.; Cabello, T.; Hernández-Suárez, E. La lagarta o bicho camello, Chrysodeixis chalcites (Esper, 1789), una plaga emergente en los cultivos de platanera de Canarias. Phytoma España 2011, 225, 21–24. [Google Scholar]
- López-Cepero, J. Análisis de residuos y gestión integrada de plagas en platanera. Phytoma España 2015, 271, 48–52. [Google Scholar]
- Horowitz, A.R.; Weintraub, P.G.; Ishaaya, I. Status of pesticide resistance in arthropod pests in Israel. Phytoparasitica 1998, 26, 231–240. [Google Scholar] [CrossRef]
- Gallego, J.R.; Guerrero-Manzano, J.; Fernández-Maldonado, F.J.; Cabello, T. Susceptibility of the egg parasitoid Trichogramma achaeae (Hymenoptera: Trichogrammatidae) to selected insecticides used in tomato greenhouses. Span. J. Agric. Res. 2019, 17, e1009. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Borges, J.; Cabrera, J.; Rodríguez-Delgado, M.A.; Hernández-Suárez, E.M.; Galán, V. Analysis of pesticide residues in bananas harvested in the Canary Islands (Spain). Food Chem. 2009, 113, 313–319. [Google Scholar] [CrossRef]
- Cabrera-Cabrera, J.; Hernández-Suárez, E.; Padilla-Cubas, A.; Jaizme-Vega, M.C.; López-Cepero, J. Banana Production under Integrated Pest Management and Organic Production Criteria: The Canary Islands Case Study. Guide Number 5. Endure Network. 2010. Available online: http://www.endure-network.eu/content/download/6427/47396/file/Banana%20Case%20Study%20Guide%20Number%205.pdf (accessed on 29 June 2021).
- del Pino, M.; Carnero, A.; Hernández-Suárez, E.; Cabello, T. Bases para la gestión integrada de Chrysodeixis chalcites (Lep.: Noctuidae) en cultivos de platanera de Canarias. Phytoma España 2015, 271, 40–46. [Google Scholar]
- Martín, A.; González, A. Guía de Gestion Integrada de Plagas: Platanera; Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente: Madrid, Spain, 2016.
- Kogan, M. Integrated Pest Management: Historical perspectives and contemporary developments. Annu. Rev. Entomol. 2003, 43, 243–270. [Google Scholar] [CrossRef]
- Miller, J.C. Ecological relationships among parasites and the practice of biological control. Environ. Entomol. 1983, 12, 620–624. [Google Scholar] [CrossRef]
- van Driesche, R.G.; Carruthers, R.I.; Center, T.; Hoddle, M.S.; Hough-Goldstein, J.; Morin, L.; Smith, L.; Wagner, D.L.; Blossey, B.; Brancatini, V.; et al. Classical biological control for the protection of natural ecosystems. Biol. Control 2010, 54, S2–S33. [Google Scholar] [CrossRef] [Green Version]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl 2017, 63, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Cabello, T. Control biológico de Noctuidos y otros Lepidópteros. In Control Biológico de Plagas; Jacas, J., Urbaneja, A., Eds.; Phytoma España: Valencia, Spain, 2009; pp. 279–306. [Google Scholar]
- Vilardebo, A.; Guérout, R. Plusia chalcites Esp. (Lépidoptére Noctuidae) nuisible au bananier dans les régions cotéres de l’ouest africain. Fruits 1964, 19, 195–208. [Google Scholar]
- Cabello, T. Natural enemies of noctuid pests (Lep., Noctuidae) on alfalfa, corn, cotton and soybean crops in southern Spain. J. Appl. Entomol. 1989, 108, 80–88. [Google Scholar] [CrossRef]
- van der Linden, A. Control of caterpillars in integrated pest management. Integrated Control in Glasshouses. IOBC/WPRS Bull. 1996, 19, 91–94. [Google Scholar]
- Garzia, G.T.; Siscaro, G.; la Malfa, G.; Lipari, V.; Noto, G.; Leonardi, C. Notes on natural enemies of armyworms infesting tomato protected crops in South-Eastern Sicily. Acta Hortic. 2003, 614, 837–841. [Google Scholar] [CrossRef]
- Murillo, H.; Vanlaerhoven, S.L.; Marcos García, M.A.; Hunt, D.W. Food web associations and effect of trophic resources and environmental factors on parasitoids expanding their host range into non-native hosts. Entomol. Exp. Appl. 2018, 166, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Pizzol, J.; Voegele, J.; Marconi, A. Efficacité de Trichogramma evanescens contre Chrysodeixis chalcites (Lep., Noctuidae) en culture de tomate sous serre. In Proceedings of the ANPP: 4° Conference International sur les Ravageurs en Agriculture, Montpellier, France, 6–8 January 1997; pp. 751–757. [Google Scholar]
- De Clercq, P.; Merleveden, F.; Mestadagh, I.; Vanderdurpel, K.; Mahaghegh, J.; Degheele, D. Predation on the tomato looper Chrysodeixis chalcites (Lep.: Noctuidae) by Podisus maculiventris and Podisus nigrispinus (Het.; Pentatomidae). J. Appl. Entomol. 1998, 122, 93–98. [Google Scholar] [CrossRef]
- Messelink, G. Biological control of caterpillars with Cotesia marginiventris (Hym.: Braconidae) in sweet pepper and tomato. IOBC/WPRS Bull. 2002, 25, 181–184. [Google Scholar]
- Garcia, P.; Oliveira, L.; Tavares, J. Natural parasitism of Chrysodeixis chalcites and Autographa gamma (Lep.: Noctuidae) eggs on tomato fields. Bol. Mus. Munic. Funchal 1998, 5, 177–181. [Google Scholar]
- Garcia, P.; Oliveira, L.; Vieira, V.; Tavares, J. Parasióides entomófagos da ilha da Madeira: Distribuiçao e hospeeiros. Bol. Da SPEN 1999, 6, 433–440. [Google Scholar]
- Polaszek, A.; Rugman-Jones, P.F.; Stouthamer, R.; Hernandez-Suárez, E.; Cabello, T.; del Pino, M. Molecular and morphological diagnoses of five species of Trichogramma: Biological control agents of Chrysodeixis chalcites (Lepidoptera: Noctuidae) and Tuta absoluta (Lepidoptera: Gelechiidae) in the Canary Islands. BioControl 2012, 57, 21–35. [Google Scholar] [CrossRef]
- del Pino, M.; Hernández-Suárez, E.; Cabello, T.; Rugman-Jones, P.; Stouthamer, R.; Polaszek, A. Trichogramma canariensis (Insecta: Hymenoptera: Trichogrammatidae) a parasitoid of Eggs of the twinspot moth Chrysodeixis chalcites (Lepidoptera: Noctuidae) in the Canary Islands. Arthropod Syst. Phylogeny 2013, 71, 169–179. [Google Scholar]
- del Pino, M.; Rugman-Jones, P.; Hernández-Suárez, E.; Polaszek, A.; Stouthamer, R. Rapid molecular identification of five species of Trichogramma occurring in the Canary Islands with notes on their distribution in banana groves. BioControl 2013, 58, 515–524. [Google Scholar] [CrossRef]
- del Pino, M.; Gallego, J.R.; Suárez, E.H.; Cabello, T. Effect of temperature on life history and parasitization behavior of Trichogramma achaeae Nagaraja and Nagarkatti (Hym.: Trichogrammatidae). Insects 2020, 11, 482. [Google Scholar] [CrossRef]
- van Harten, A.; Miranda, A.M.C. Importation, rearing, release and recapture of parasites of pests in Cape Verde during the period March 1981–March 1983. Rev. Investig. Agrária 1985, 1, 13–25. [Google Scholar]
- Lima, M.L.L.; van Harten, A. Biological control of crop pests in Cape Verde. Current situation and future programmes. Rev. Investig. Agrária 1985, 1, 3–12. [Google Scholar]
- Hernández, M.P. Seguimiento de Chrysodeixis chalcites y Spodoptera littoralis en platanera en la isla de El Hierro; búsqueda de entomófagos y entomopatógenos. In Trabajo Fin de Carrera. Escuela Técnica Superior de Ingeniería Agraria; Universidad de La Laguna: Tenerife, Spain, 2007; p. 79. [Google Scholar]
- Bernal, A.; Williams, T.; Hernández-Suárez, E.; Carnero, A.; Caballero, P.; Simón, O. A native variant of Chrysodeixis chalcites nucleopolyhedrovirus: The basis for a promising bioinsecticide for control of C. chalcites on Canary Islands’ banana crops. Biol. Control 2013, 67, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Simón, O.; Bernal, A.; Williams, T.; Carnero, A.; Hernández-Suárez, E.; Muñoz, D.; Caballero, P. Efficacy of an alphabaculovirus-based biological insecticide for control of Chrysodeixis chalcites (Lepidoptera: Noctuidae) on tomato and banana crops. Pest Manag. Sci. 2015, 71, 1623–1630. [Google Scholar] [CrossRef]
- Fuentes, E.G.; Hernández-Suárez, E.; Simón, O.; Williams, T.; Caballero, P. Chrysodeixis chalcites nucleopolyhedrovirus (ChchNPV): Natural occurrence and efficacy as a biological insecticide on young banana plants in greenhouse and open-field conditions on the Canary Islands. PLoS ONE 2017, 12, e0181384. [Google Scholar] [CrossRef] [Green Version]
- Cabello, T.; Gallego, J.R.; Fernández, F.J.; Gámez, M.; Vila, E.; del Pino, M.; Hernández-Suárez, E. Biological control strategies for the south american tomato moth Tuta absoluta (Lepidoptera: Gelechiidae) in greenhouse tomatoes. J. Econ. Entomol. 2012, 105, 2085–2096. [Google Scholar] [CrossRef]
- Cakmak, T.; Piedra-Buena, A.; Hernández-Suárez, E.; Álvarez, C. Chrysodeixis chalcites (Esper) (Lepidoptera: Noctuidae) oviposition preferences on different growing stages of banana (Musa acuminate Colla, Musaceae) plants. Phytoparasitica 2019, 47, 485–498. [Google Scholar] [CrossRef]
- Townsend, G.R.; Heuberger, J.W. Methods for estimating losses caused by diseases in fungicide experiments. Plant Dis. Report. 1943, 27, 340–343. [Google Scholar]
- Jamovi Statistical Software v. 2.2. Available online: https://www.jamovi.org (accessed on 14 March 2022).
- Bosque, J.L.; Figueras, M.; Izquierdo, J. Parasitismo sobre Plusiinae (Lepidoptera: Noctuidae) en tomate. Boletín Sanid. Veg. Plagas 1996, 22, 683–692. [Google Scholar]
- Alcázar, M.D.; Belda, J.E.; Barranco, P.; Cabello, T. Parasitoides de Especies Plaga en Hortícolas de Invernaderos en Almería; Junta de Andalucía, Consejería de Agricultura y Pesca: Sevilla, Spain, 2002; p. 181. [Google Scholar]
- Gabarra, R.; Arnó, J.; Castañé, C.; Izquierdo, J.; Alomar, O.; Riudavets, J.; Albajes, R. Fauna útil trobada en els cultius d’horta de Catalunya. Doss. Agrar. 2000, 6, 83–103. [Google Scholar]
- Hervet, V.A.; Murillo, H.; Fernández-Triana, J.L.; Shaw, M.R.; Laird, R.A.; Floate, K.D. First report of Cotesia vanessae (Hymenoptera: Braconidae) in North America. Can. Entomol. 2014, 146, 560–566. [Google Scholar] [CrossRef]
- Michel-Salzat, A.; Whitfield, J.B. Preliminary evolutionary relationships within the parasitoid wasp genus Cotesia (Hymenoptera: Braconidae: Microgastrinae): Combined analysis of four genes. Syst. Entomol. 2004, 29, 371–382. [Google Scholar] [CrossRef]
- Arechavaleta, M.; Rodríguez, S.; Zurita, N.; García, A. Lista de Especies Silvestres de Canarias. Hongos, Plantas y Animales Terrestres; Gobierno de Canarias : Santa Cruz de Tenerife, Spain, 2010; p. 579. [Google Scholar]
- Laing, J.E.; Levin, D.B. A review of the biology and a bibliography of Apanteles glomeratus (L.) (Hymenoptera: Braconidae). Biocontrol News Inf. 1982, 3, 7–23. [Google Scholar]
- Lozan, A.I.; Monaghan, M.T.; Spitzer, K.; Jaroš, J.; Žurovcová, M.; Brož, V. DNA-based confirmation that the parasitic wasp Cotesia glomerata (Braconidae, Hymenoptera) is a new threat to endemic butterflies of the Canary Islands. Conserv. Genet. 2008, 9, 1431–1437. [Google Scholar] [CrossRef]
- van Achterberg, C.; Shaw, M.R. Revision of the western Palaearctic species of Aleiodes Wesmael (Hymenoptera, Braconidae, Rogadinae). Part 1: Introduction, key to species groups, outlying distinctive species, and revisionary notes on some further species. ZooKeys 2016, 639, 1–164. [Google Scholar] [CrossRef]
- Guerrero, E.R.; Koponen, M. Contribución a la fauna braconológica de las Islas Canarias (Hymenoptera, Braconidae). Lleraea 2000, 28, 99–117. [Google Scholar]
- Figueiredo, E.; Mexia, A. Parasitoid complex associated with lepidoptera on horticultural protected crops in the Oeste region of Portugal. Bull. OILB/SROP 2000, 23, 205–208. [Google Scholar]
- Baumgarten, H.T.; Fiedler, K. Parasitoids of lycaenid butterfly caterpillars: Different patterns in resource use and their impact on the hosts’ symbiosis with ants. Zool. Anz. 1997, 236, 167–180. [Google Scholar]
- Torres-Vila, L.M.; Rodríguez-Molina, M.C.; Palo, E.; del Estal, P.; Lacasa, A. El complejo parasitario larvario de Helicoverpa armígera Hübner sobre tomate en las Vegas del Guadiana (Extremadura). Bol. San. Veg. Plagas 2000, 26, 323–333. [Google Scholar]
- Medina, P.; Morales, J.J.; Budia, F.; Adan, A.; del Estal, P.; Viñuela, E. Compatibility of endoparasitoid Hyposoter didymator (Hymenoptera: Ichneumonidae) protected stages with five selected insecticides. J. Econ. Entomol. 2007, 100, 1789–1796. [Google Scholar] [CrossRef]
- Jones, R.A. Ctenochares bicolorus (L.), an African ichneumonid (Hymenoptera) found in Britain. Br. J. Ent. Nat. Hist. 2001, 14, 96–99. [Google Scholar]
- van Lenteren, J.C.; Bolckmans, K.; Kohl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Urbaneja, A.; van der Blom, J.; Lara, L.; Timmer, R.; Blockmans, K. Utilización de Cotesia marginiventris (Cresson) (Hym.: Braconidae) para el control biológico de orugas (Lep.: Noctuidae) en el manejo integrado de plagas en pimiento bajo invernadero. Boletín Sanid. Veg. Plagas 2002, 28, 239–250. [Google Scholar]
- Riddick, E.W. Production of the parasitoid Cotesia marginiventris (Hymenoptera: Braconidae) in unicellular trays using the host Spodoptera exigua (Lepidoptera: Noctuidae). J. Entomol. Sci. 2004, 39, 362–372. [Google Scholar] [CrossRef]
- Nagaraja, H.; Ramesh, B.; Ravindra, A.C. Trichogrammatids used commercially by Biotech International Ltd., Bangalore, India. Egg Parasit. News 2002, 14, 32–33. [Google Scholar]
- Cabello, T.; Vargas, P. Resistencia de los estados de desarrollo de Trichogramma cordubensis y T. pintoi (Hym.: Trichogrammatidae) a las altas temperaturas. Boletín Sanid. Veg. Plagas 1989, 15, 263–266. [Google Scholar]
- Oliveira, R.C.; Pratissoli, D.; Rodrigues–Vianna, U.; Fialho, E. Influencia do estresse sobre as características biologicas de femeas do parasitóide de ovos Trichogramma pretiosum (Hym.: Trichogrammatidae), quando submetidos a temperaturas extremas. Acad. Insecta 2003, 2, 1–6. [Google Scholar]
- Smith, S.M. Biological control with Trichogramma: Advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41, 375–406. [Google Scholar] [CrossRef]
- Sithanantham, S.; Ballal, C.R.; Jalali, S.K.; Bakthavatsalam, N. Biological Control of Insect Pests Using Egg Parasitoids; Springer: New Delhi, India, 2013. [Google Scholar]
- Mills, N. Egg Parasitoids in Biological Control and Integrated Pest Management. In Egg Parasitoids in Agroecosystems with Emphasis on Trichogramma; Consoli, F., Parra, J., Zucchi, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 389–411. [Google Scholar] [CrossRef]
- Pizzol, J.; Frandon, J.; Ferran, A.; Marconi, A.; Migliore, O.; Bezert, J. Study of the dispersion of Trichogramma evanescens Westwood in protected tomato crops. In Proceedings of the Colloque International Tomate sous Abri, Protection Intégrée—Agriculture Biologique, Avignon, France, 17–19 September 2003; pp. 168–172. [Google Scholar]
- Suverkropp, B.P.; Bigler, F.; van Lenteren, J.C. Dispersal behaviour of Trichogramma brassicae in maize fields. Bull. Insectology 2009, 62, 113–120. [Google Scholar]
- Chapman, A.V.; Kuhar, T.P.; Schultz, P.B.; Brewster, C.C. Dispersal of Trichogramma ostriniae (Hymenoptera: Trichogrammatidae) in potato fields. Environ. Entomol. 2009, 38, 677–685. [Google Scholar] [CrossRef] [Green Version]
- López-Cepero, J.; Piedra-Buena, A.; Puerta, M. Biodiversidad en la Platanera; Cooperativa Platanera de Canarias (COPLACA): Santa Cruz de Tenerife, Spain, 2021. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Island | No. of Eggs | Parasitism (%) | ||
|---|---|---|---|---|
| Collected | Non-Parasitized | Parasitized | ||
| El Hierro | 53 | 18 | 35 | 66.04 |
| Gran Canaria | 248 | 112 | 136 | 54.84 |
| La Palma | 1097 | 109 | 988 | 90.06 |
| Tenerife | 1370 | 702 | 668 | 48.76 |
| Total | 2776 | 944 | 1785 | 64.30 |
| Island | No. Fields | No. Eggs per Field | Parasitism per Field (%) | ||
|---|---|---|---|---|---|
| Sampled | Parasitized | Average | Maximum | ||
| El Hierro | 7 | 7.57 ± 11.90 c | 5.00 ± 10.57 c | 48.57 ± 11.27 ab | 100.00 |
| Gran Canaria | 2 | 58.50 ± 22.27 a | 41.50 ± 19.78 a | 70.82 ± 21.08 ab | 71.64 |
| La Palma | 22 | 49.14 ± 6.71 b | 42.05 ± 5.96 a | 81.01 ± 6.36 a | 100.00 |
| Tenerife | 28 | 48.93 ± 5.95 b | 23.86 ± 5.28 b | 38.58 ± 5.63 b | 97.06 |
| Total | 59 | 44.42 ± 4.38 | 29.00 ± 3.89 | 56.68 ± 4.59 | 100.00 |
| Banana Field System | Noctuid Species | No. of Larvae | Parasitism (d) | ||
|---|---|---|---|---|---|
| Collected (a) | Died (b) | Parasitized (c) | (d = 100c/a − b) | ||
| Open field | Ch. Chalcites | 1203 | 376 | 121 | 14.63 |
| C. circunflexa | 92 | 10 | 10 | 12.20 | |
| S. exigua | 5 | 0 | 0 | 0.0 | |
| S. littoralis | 62 | 15 | 0 | 0.0 | |
| T. orichalcea | 13 | 0 | 0 | 0.0 | |
| Greenhouse | Ch. Chalcites | 4276 | 1381 | 147 | 5.08 |
| C. circunflexa | 3 | 0 | 0 | 0.0 | |
| S. exigua | 42 | 6 | 0 | 0.0 | |
| S. littoralis | 226 | 48 | 7 | 3.93 | |
| Total | 5922 | 1836 | 285 | 6.98 | |
| Parasitoid | Host Plant | Host | No. Fields 2 | Parasitism per Field (%) | ||
|---|---|---|---|---|---|---|
| Species | Stage at Sampling 1 | Average | Maximum | |||
| Aplomyia confinis | Geranium | Ch. chalcites | L2–L5 | 1 | 5.26 | 5.26 |
| Banana | Ch. chalcites | L2–L5 | 16 | 4.22 | 10.35 | |
| Cotesia glomerata | Banana | Ch. chalcites | L3–L4 | 2 | 3.01 | 3.70 |
| Cotesia sp. | Squash | Ch. chalcites | L2–L4 | 1 | 16.67 | 16.67 |
| Geranium | Ch. chalcites | L2–L4 | 1 | 12.50 | 12.50 | |
| Bean | Ch. chalcites | L2–L4 | 2 | 5.33 | 7.14 | |
| Potato | Ch. chalcites | L2–L4 | 1 | 4.17 | 4.17 | |
| Pepper | Ch. chalcites | L2–L4 | 1 | 12.50 | 12.50 | |
| Banana | Ch. chalcites | L2–L4 | 11 | 8.18 | 19.23 | |
| Tomato | Ch. chalcites | L2–L4 | 6 | 13.02 | 33.33 | |
| N. glauca | C. circunflexa | L2–L4 | 1 | 4.55 | 4.55 | |
| Ctenochares bicolorus | Banana | Ch. chalcites | L3–P | 3 | 4.60 | 6.89 |
| Hyposoter rufiventris | Geranium | Ch. chalcites | L2–L4 | 1 | 5.26 | 5.26 |
| Potato | Ch. chalcites | L2–L4 | 1 | 1.96 | 1.96 | |
| Banana | Ch. chalcites | L2–L4 | 5 | 2.18 | 3.49 | |
| Banana | S. littoralis | L2–L4 | 2 | 17.19 | 25.00 | |
| Tomato | Ch. chalcites | L2–L4 | 3 | 7.42 | 8.33 | |
| Aleiodes sp. | N. glauca | Ch. chalcites | L2–L3 | 1 | 4.76 | 4.76 |
| Tomato | Ch. chalcites | L2–L3 | 1 | 4.88 | 4.88 | |
| Species | Year and Island a | Field System b | ||||
|---|---|---|---|---|---|---|
| 2007 | 2008 | 2009 | 2010 | Openfield | Greenhouse | |
| Larval/Pupa parasitoids | ||||||
| Braconidae | ||||||
| Cotesia sp. | TF 3, LP 9,10,12 | TF 1,2,3,5,6,7, LP 10 | Ba, To, Sq, Be, Ge, Ng | Ba, Ca, Pe, Po | ||
| Cotesia glomerata (Linnaeus, 1758) | TF 3, HR 14 | Ng | Ba | |||
| Aleiodes sp. | HR 14 | TF 5 | To, Ng | |||
| Ichneumonidae | ||||||
| Ctenochares bicolorus (Linnaeus, 1767) | TF 3 | TF 3 | TF 1, LP 9 | Ba | Ba | |
| Hyposoter rufiventris (Pérez, 1895) | TF 7 | TF 3, HR 14 | TF 3,5, LP 12 | Ba, To, Ge | Ba, Po | |
| Tachinidae | ||||||
| Aplomyia confinis (Fallén, 1820) | TF 2,5 | TF 3 | TF 1, LP 8,12 | Ba, Ge | Ba | |
| Egg parasitoids | ||||||
| Trichogrammatidae | ||||||
| Trichogramma achaeae Nagaraja and Nagarkatti, 1970 | TF 3, LP 9,10, HR 14 | TF 1,4,5,7, LP 8,9,10,11, HR 14, GC 13 | LP 9 | Ba, To, Ca, Be, Ng, Sn | Ba, Pe, Ca, Sq | |
| Trichogramma bourarachae Pintureau and Babault, 1988 | TF 4,7 | Ng | ||||
| Trichogramma canariensis del Pino and Polaszek, 2013 | GC 13 | GC 13 | Ba, To | Ba | ||
| Trichogramma euproctidis (Guirault, 1911) | TF 3 | TF 1,3,4,5,7 | Ba, To, Ng, Sn | Ba, To, Ng | ||
| Trichogramma evanescens Westwood, 1833 | TF 1,4,6, LP 9 | To, Ng | Ca, Sq | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
del Pino, M.; Cabello, T.; Hernández-Suárez, E. Biological Control Options for the Golden Twin-Spot Moth, Chrysodeixis chalcites (Esper) (Lepidoptera: Noctuidae) in Banana Crops of the Canary Islands. Insects 2022, 13, 516. https://doi.org/10.3390/insects13060516
del Pino M, Cabello T, Hernández-Suárez E. Biological Control Options for the Golden Twin-Spot Moth, Chrysodeixis chalcites (Esper) (Lepidoptera: Noctuidae) in Banana Crops of the Canary Islands. Insects. 2022; 13(6):516. https://doi.org/10.3390/insects13060516
Chicago/Turabian Styledel Pino, Modesto, Tomás Cabello, and Estrella Hernández-Suárez. 2022. "Biological Control Options for the Golden Twin-Spot Moth, Chrysodeixis chalcites (Esper) (Lepidoptera: Noctuidae) in Banana Crops of the Canary Islands" Insects 13, no. 6: 516. https://doi.org/10.3390/insects13060516
APA Styledel Pino, M., Cabello, T., & Hernández-Suárez, E. (2022). Biological Control Options for the Golden Twin-Spot Moth, Chrysodeixis chalcites (Esper) (Lepidoptera: Noctuidae) in Banana Crops of the Canary Islands. Insects, 13(6), 516. https://doi.org/10.3390/insects13060516